
Polyphasic Identification of Distoseptispora with Six New Species from Fresh Water

Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection, Morphological Studies and Isolation
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Phylogenetic Analyses
3. Results
3.1. Phylogenetic Analyses
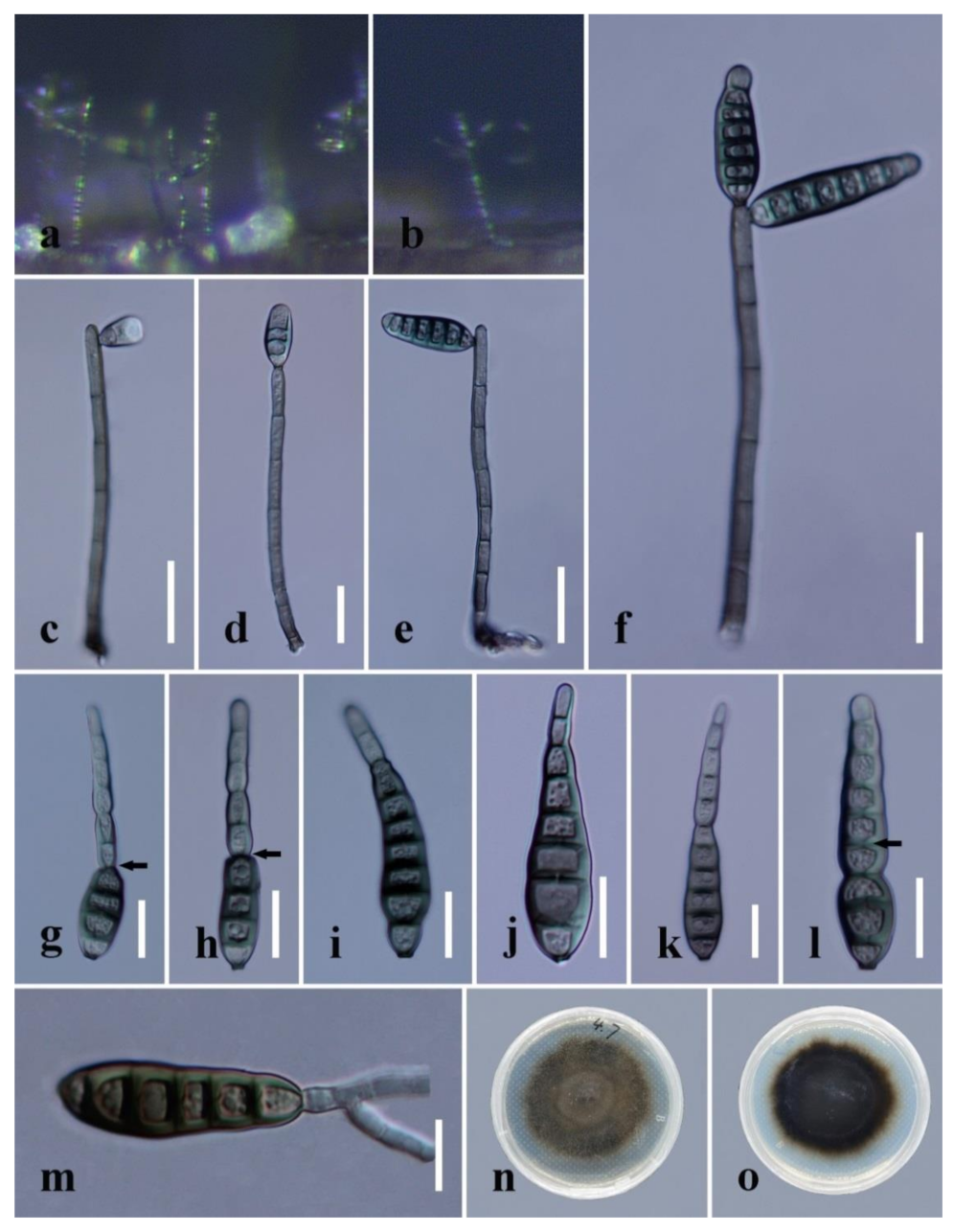
3.2. Taxonomy in Distoseptispora
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shearer, C.A.; Descals, E.; Kohlmeyer, B.; Kohlmeyer, J.; Marvanová, L.; Padgett, D.; Porter, D.; Raja, H.A.; Schmit, J.P.; Thorton, H.A.; et al. Fungal biodiversity in aquatic habitats. Biodivers. Conserv. 2007, 16, 49–67. [Google Scholar] [CrossRef]
- Hyde, K.; Goh, T. Fungi on submerged wood in Lake Barrine, north Queensland, Australia. Mycol. Res. 1998, 102, 739–749. [Google Scholar] [CrossRef]
- Hyde, K.D.; Hongsanan, S.; Jeewon, R.; Bhat, D.J.; McKenzie, E.H.C.; Jones, E.B.G.; Phookamsak, R.; Ariyawansa, H.A.; Boonmee, S.; Zhao, Q.; et al. Fungal diversity notes 367–490: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 80, 1–270. [Google Scholar] [CrossRef]
- Zhang, H.; Dong, W.; Hyde, K.D.; Maharachchikumbura, S.S.N.; Hongsanan, S.; Jayarama Bhat, D.; Al-Sadi, A.M.; Zhang, D. Towards a natural classification of Annulatascaceae-like taxa: Introducing Atractosporales ord. nov. and six new families. Fungal Divers. 2017, 85, 75–110. [Google Scholar] [CrossRef]
- Yang, J.; Maharachchikumbura, S.S.N.; Liu, J.K.; Hyde, K.D.; Jones, E.B.G.; Al-Sadi, A.M.; Liu, Z.Y. Pseudostanjehughesia aquitropica gen. et sp. nov. and Sporidesmium sensu lato species from freshwater habitats. Mycol. Prog. 2018, 17, 591–616. [Google Scholar] [CrossRef]
- Cai, L.; Zhang, K.Q.; McKenzie, E.H.C.; Hyde, K.D. Freshwater fungi from bamboo and wood submerged in the Liput River in the Philippines. Fungal Divers. 2003, 13, 1–12. [Google Scholar]
- Su, H.Y.; Hyde, K.D.; Maharachchikumbura, S.S.N.; Ariyawansa, H.A.; Luo, Z.L.; Promputtha, I.; Tian, Q.; Lin, C.G.; Shang, Q.J.; Zhao, Y.C.; et al. The families Distoseptisporaceae fam. nov., Kirschsteiniotheliaceae, Sporormiaceae and Torulaceae, with new species from freshwater in Yunnan Province, China. Fungal Divers. 2016, 80, 375–409. [Google Scholar] [CrossRef]
- Luo, Z.-L.; Hyde, K.D.; Liu, J.-K.J.; Maharachchikumbura, S.S.N.; Jeewon, R.; Bao, D.-F.; Bhat, D.J.; Lin, C.-G.; Li, W.-L.; Yang, J.; et al. Freshwater Sordariomycetes. Fungal Divers. 2019, 99, 451–660. [Google Scholar] [CrossRef]
- Dong, W.; Hyde, K.D.; Jeewon, R.; Doilom, M.; Yu, X.-D.; Wang, G.-N.; Liu, N.G.; Hu, D.M.; Nalumpang, S.; Zhang, H. Towards a natural classification of annulatascaceae-like taxa II: Introducing five new genera and eighteen new species from freshwater. Mycosphere 2021, 12, 1–88. [Google Scholar] [CrossRef]
- Link, H.F. Observationes in ordines plantarumnaturales. Diss. I. Mag. Ges. Naturf. Freunde Berl. 1809, 3, 3–42. [Google Scholar]
- Ellis, M.B. Clasterosporium and some allied Dematiaceae—Phragmosporae, I. Mycol. Pap. 1958, 70, 1–89. [Google Scholar]
- Ellis, M.B. (Ed.) Dematiaceous Hyphomycetes; Commonwealth Mycological Institute: London, UK, 1971. [Google Scholar]
- Subramanian, C.V. A reassessment of Sporidesmium (Hyphomycetes) and some related taxa. Proc. Indian Natl. Sci. Acad. USA 1992, 58, 179–190. [Google Scholar]
- Wu, W.; Zhuang, W. Sporidesmium, Endophragmiella and Related Genera from China; Fungal Diversity Press: Hong Kong, China, 2005. [Google Scholar]
- Hyde, K.D.; Dong, Y.; Phookamsak, R.; Jeewon, R.; Bhat, D.J.; Jones, E.G.; Liu, N.-G.; Abeywickrama, P.D.; Mapook, A.; Wei, D.; et al. Fungal diversity notes 1151–1276: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2020, 100, 5–277. [Google Scholar] [CrossRef]
- Xia, J.W.; Ma, Y.R.; Li, Z.; Zhang, X.G. Acrodictys-like wood decay fungi from southern China, with two new families Acrodictyaceae and Junewangiaceae. Sci. Rep.-UK 2017, 7, 7888. [Google Scholar] [CrossRef]
- Luo, Z.; Hyde, K.; Liu, J.; Bhat, D.; Bao, D.; Li, W.; Su, H. Lignicolous freshwater fungi from China II: Novel Distoseptispora (Distoseptisporaceae) species from northwestern Yunnan Province and a suggested unified method for studying lignicolous freshwater fungi. Mycosphere 2018, 9, 444–461. [Google Scholar] [CrossRef]
- Tibpromma, S.; Hyde, K.D.; McKenzie, E.H.C.; Bhat, D.J.; Phillips, A.J.L.; Wanasinghe, D.N.; Samarakoon, M.C.; Jayawardena, R.S.; Dissanayake, A.J.; Tennakoon, D.S.; et al. Fungal diversity notes 840–928: Micro-fungi associated with Pandanaceae. Fungal Divers. 2018, 93, 1–160. [Google Scholar] [CrossRef]
- Crous, P.; Wingfield, M.; Lombard, L.; Roets, F.; Swart, W.; Alvarado, P.; Carnegie, A.; Moreno, G.; Luangsaard, J.; Thangavel, R.; et al. Fungal Planet description sheets: 951–1041. Persoonia 2019, 43, 223–425. [Google Scholar] [CrossRef]
- Phookamsak, R.; Hyde, K.D.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Raspé, O.; Karunarathna, S.C.; Wanasinghe, D.N.; Hongsanan, S.; et al. Fungal diversity notes 929–1035: Taxonomic and phylogenetic contributions on genera and species of fungi. Fungal Divers. 2019, 95, 1–273. [Google Scholar] [CrossRef]
- Phukhamsakda, C.; McKenzie, E.H.C.; Phillips, A.J.L.; Jones, E.B.G.; Bhat, D.J.; Marc, S.; Bhunjun, C.S.; Wanasinghe, D.N.; Thongbai, B.; Camporesi, E.; et al. Microfungi associated with Clematis (Ranunculaceae) with an integrated approach to delimiting species boundaries. Fungal Divers. 2020, 102, 1–203. [Google Scholar] [CrossRef]
- Sun, Y.; Goonasekara, I.D.; Thambugala, K.M.; Jayawardena, R.S.; Wang, Y.; Hyde, K.D. Distoseptispora bambusae sp. nov.(Distoseptisporaceae) on bamboo from China and Thailand. Biodivers. Data J. 2020, 8, e53678. [Google Scholar] [CrossRef]
- Li, W.-L.; Liu, Z.-P.; Zhang, T.; Dissanayake, A.J.; Luo, Z.-L.; Su, H.-Y.; Liu, J.-K. Additions to Distoseptispora (Distoseptisporaceae) associated with submerged decaying wood in China. Phytotaxa 2021, 520, 75–86. [Google Scholar] [CrossRef]
- Yang, J.; Liu, L.-L.; Jones, E.B.G.; Li, W.-L.; Hyde, K.D.; Liu, Z.-Y. Morphological variety in Distoseptispora and introduction of six novel species. J. Fungi 2021, 7, 945. [Google Scholar] [CrossRef] [PubMed]
- Phukhamsakda, C.; Nilsson, R.H.; Bhunjun, C.S.; de Farias, A.R.G.; Sun, Y.-R.; Wijesinghe, S.N.; Raza, M.; Bao, D.-F.; Lu, L.; Tibpromma, S.; et al. The numbers of fungi: Contributions from traditional taxonomic studies and challenges of metabarcoding. Fungal Divers. 2022, 114, 327–386. [Google Scholar] [CrossRef]
- Chomnunti, P.; Hongsanan, S.; Aguirre-Hudson, B.; Tian, Q.; Peršoh, D.; Dhami, M.K.; Alias, A.S.; Xu, J.C.; Liu, X.Z.; Stadler, M.; et al. The sooty moulds. Fungal Divers. 2014, 66, 1–36. [Google Scholar] [CrossRef]
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.A.; Bhat, J.; Buyck, B.; Cai, L.; Dai, Y.C.; Abd-Elsalam, K.A.; Ertz, D.; Hidayat, I.; et al. The Faces of Fungi database: Fungal names linked with morphology, phylogeny and human impacts. Fungal Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols a Guide to Methods and Applications; Academic Press, Inc.: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 19, 1160–1166. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Miller, M.A.; Pfeiffer, W.T.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; San Diego Supercomputer Center: La Jolla, CA, USA, 2010; pp. 1–8. [Google Scholar]
- Miller, M.A.; Schwartz, T.; Pickett, B.E.; He, S.; Klem, E.B.; Scheuermann, R.H.; Passarotti, M.; Kaufman, S.; O’Leary, M.A. A RESTful API for Access to Phylogenetic Tools via the CIPRES Science Gateway. Evol. Bioinform. 2015, 11, 43–48. [Google Scholar] [CrossRef]
- Nylander, J.A.A. (Ed.) MrModeltest V2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Zhaxybayeva, O.; Gogarten, J.P. Bootstrap, Bayesian probability and maximum likelihood mapping: Exploring new tools for comparative genome analyses. BMC Genom. 2002, 3, 4. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree v1. 4.2, A Graphical Viewer of Phylogenetic Trees. Available from Left Angle Bracket. 2014. Available online: http://tree.bio.ed.ac.uk/software/figtree/rightanglebracket (accessed on 5 May 2022).
- Shoemaker, R.A. Lasiosphaeria caesariata with Sporidesmium hormiscioides and L. triseptata with S. adscendens. Sydowia 1985, 38, 278–283. [Google Scholar]
- Réblová, M. Studies in Chaetosphaeria sensu lato III. Umbrinosphaeria gen. nov. and Miyoshiella with Sporidesmium anamorphs. Mycotaxon 1999, 71, 13–43. [Google Scholar]
- Shenoy, B.D.; Jeewon, R.; Wu, W.P.; Bhat, D.J.; Hyde, K.D. Ribosomal and RPB2 DNA sequence analyses suggest that Sporidesmium and morphologically similar genera are polyphyletic. Mycol. Res. 2006, 110, 916–928. [Google Scholar] [CrossRef]
- Zhai, Z.-J.; Yan, J.-Q.; Li, W.-W.; Gao, Y.; Hu, H.-J.; Zhou, J.-P.; Song, H.-Y.; Hu, D.-M. Three novel species of Distoseptispora (Distoseptisporaceae) isolated from bamboo in Jiangxi Province, China. Mycokeys 2021, 88, 35–54. [Google Scholar] [CrossRef]
- Jeewon, R.; Hyde, K.D. Establishing species boundaries and new taxa among fungi: Recommendations to resolve taxonomic ambiguities. Mycosphere 2016, 7, 1669–1677. [Google Scholar] [CrossRef]
- Hyde, K.D.; Tennakoon, D.S.; Jeewon, R.; Bhat, D.J.; Maharachchikumbura, S.S.N.; Rossi, W.; Leonardi, M.; Lee, H.B.; Mun, H.Y.; Houbraken, J.; et al. Fungal diversity notes 1036–1150: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2019, 96, 1–242. [Google Scholar] [CrossRef]
- Hyde, K.D.; Suwannarach, N.; Jayawardena, R.S.; Manawasinghe, I.S.; Liao, C.F.; Doilom, M.; Cai, L.; Zhao, P.; Buyck, B.; Phukhamsakda, C.; et al. Mycosphere notes 325–344—Novel species and records of fungal taxa from around the world. Mycosphere 2021, 12, 1101–1156. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D.; Chen, Y.J.; Ba, D.F.; Boonmee, S.; Calabon, M.S.; et al. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Crane, J.L.; Dumont, K.P. Hyphomycetes from the West Indies and Venezuela. Can. J. Bot. 1975, 53, 843–851. [Google Scholar] [CrossRef]
- Baker, W.A.; Partridge, C.; Morganjones, G. Notes on hypomycetes. LXXXVII. Rhexoacrodictys, a new segregate genus to accomodate four species previously classified n acrodictys. Mycotaxon 2002, 82, 95–113. [Google Scholar]
- Delgado, G. South Florida microfungi: Veramycella bispora, a new palmicolous, anamorphic genus and species, with some new records for the continental USA. Mycotaxon 2009, 107, 357–373. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | PCR Primers | PCR Amplification Reaction System | PCR Conditions |
|---|---|---|---|
| LSU, ITS | LR5/LROR [28], ITS5/ITS4 [29] | Total: 25 μL 12.5 μL Master Mix 9.5 μL ddH2O 1 μL of each primer (10 μM) 1 μL DNA template | 1. 94 °C, 3 min 2. 94 °C, 45 s 3. 56 °C, 50 s 4. 72 °C, 1 min 5. Repeat steps 2–4 for 35 cycles 6. 72 °C, 10 min 7. Keep at 4 °C |
| TEF1-α | 983F/2218R [30] | Total: 50 μL 45 μL T3 Mix 2 μL of each primer (10 μM) 1 μL DNA template | 1. 98 °C, 2 min 2. 98 °C, 10 s 3. 56 °C, 10 s 4. 72 °C, 20 s 5. Repeat steps 2–4 for 35 cycles 6. 72 °C, 5 min 7. Keep at 4 °C |
| RPB2 | 5F/7cR [31] | Total: 50 μL 45 μL T3 Mix 2 μL of each primer (10 μM) 1 μL DNA template | 1. 98 °C, 2 min 2. 98 °C, 10 s 3. 51 °C, 10 s 4. 72 °C, 20 s 5. Repeat steps 2–4 for 35 cycles 6. 72 °C, 5 min 7. Keep at 4 °C |
| Species | Voucher/Culture | GenBank Accession Numbers | |||
|---|---|---|---|---|---|
| LSU | ITS | TEF1-α | RPB2 | ||
| Acrodictys bambusicola | CGMCC 3.18641 | KX033564 | KU999973 | – | – |
| Acrodictys elaeidicola | CGMCC 3.18642 | KX033569 | KU999978 | – | – |
| Aquapteridospora aquatica | MFLUCC 17–2371T | MW287767 | MW286493 | – | – |
| Aquapteridospora fusiformis | MFLU 18–1601T | MK849798 | MK828652 | MN194056 | – |
| Aquapteridospora lignicola | MFLUCC 15–0377T | KU221018 | MZ868774 | MZ892980 | MZ892986 |
| Bullimyces aurisporus | AF316–1bT | JF775590 | – | – | – |
| Bullimyces communis | AF281–5 | JF775587 | – | – | – |
| Cancellidium applanatum | CBS 337.76 | MH872755 | NR159777 | _ | _ |
| Cancellidium cinereum | MFLUCC 18–0424T | MT370363 | MT370353 | MT370488 | MT370486 |
| Distoseptispora adscendens | HKUCC 10820 | DQ408561 | – | – | DQ435092 |
| Distoseptispora amniculi | MFLUCC17–2129T | MZ868761 | MZ868770 | – | MZ892982 |
| Distoseptispora appendiculata | MFLUCC 18–0259T | MN163023 | MN163009 | MN174866 | – |
| Distoseptispora aqualignicola | KUNCC 21–10729T | ON40845 | OK341186 | OP413480 | OP413474 |
| Distoseptispora aquamyces | KUNCC 21–10732T | OK341199 | OK341187 | OP413482 | OP413476 |
| Distoseptispora aquatica | MFLUCC 15–0374T | KU376268 | MF077552 | – | – |
| Distoseptispora aquatica | MFLUCC 16–0904 | MK849794 | MK828649 | MN194053 | – |
| Distoseptispora aquatica | MFLUCC 18–0646 | MK849793 | MK828648 | MN194052 | – |
| Distoseptispora aquatica | S-965 | MK849792 | MK828647 | MN194051 | MN124537 |
| Distoseptispora atroviridis | GZCC 20–0511T | MZ868763 | MZ868772 | MZ892978 | MZ892984 |
| Distoseptispora atroviridis | GZCC 19–0531 | MZ227223 | MW133915 | – | – |
| Distoseptispora bambusae | MFLUCC 20–0091T | MT232718 | MT232713 | MT232880 | MT232881 |
| Distoseptispora bambusae | MFLUCC 14–0583 | MT232717 | MT232712 | – | MT232882 |
| Distoseptispora bambusae | KUNCC 21–10732 | OK341200 | OK341188 | OP413492 | OP413487 |
| Distoseptispora bangkokensis | MFLUCC 18–0262T | MZ518206 | MZ518205 | – | – |
| Distoseptispora cangshanensis | MFLUCC 16–0970 T | MG979761 | MG979754 | MG988419 | – |
| Distoseptispora caricis | CPC 36498T | MN567632 | MN562124 | – | MN556805 |
| Distoseptispora caricis | CPC 36442T | – | MN562125 | – | MN556806 |
| Distoseptispora chinense | GZCC 21–0665 T | MZ474867 | MZ474871 | MZ501609 | – |
| Distoseptispora clematidis | MFLUCC 17–2145T | MT214617 | MT310661 | – | MT394721 |
| Distoseptispora clematidis | KUMCC 21–10727 | OK341197 | OK341184 | OP413488 | OP413483 |
| Distoseptispora crassispora | KUMCC 21–10726T | OK341196 | OK310698 | OP413479 | OP413473 |
| Distoseptispora curvularia | KUMCC 21–10725T | OK341195 | OK310697 | OP413478 | OP413472 |
| Distoseptispora dehongensis | KUMCC 18–0090 T | MK079662 | MK085061 | MK087659 | – |
| Distoseptispora dehongensis | MFLUCC 19–0335 | OK341201 | OK341189 | OP413491 | OP413486 |
| Distoseptispora dehongensis | MFLUCC 17–2326 | OK341193 | OK341183 | OP413493 | – |
| Distoseptispora effusa | GZCC 19–0532T | MZ227224 | MW133916 | MZ206156 | – |
| Distoseptispora euseptata | MFLUCC 20–0154T | MW081544 | MW081539 | – | MW151860 |
| Distoseptispora euseptata | DLUCC S2024 | MW081545 | MW081540 | MW084994 | MW084996 |
| Distoseptispora fasciculata | KUMCC 19–0081T | MW287775 | MW286501 | MW396656 | – |
| Distoseptispora fluminicola | MFLUCC 15–0417T | KU376270 | NR154041 | – | – |
| Distoseptispora fusiformis | GZCC 20–0512T | MZ868764 | MZ868773 | MZ892979 | MZ892985 |
| Distoseptispora guizhouensis | GZCC 21–0666T | MZ474869 | MZ474868 | MZ501610 | MZ501611 |
| Distoseptispora guttulata | MFLUCC 16–0183T | MF077554 | MF077543 | MF135651 | – |
| Distoseptispora hyalina | MFLUCC 17–2128 T | MZ868760 | MZ868769 | MZ892976 | MZ892981 |
| Distoseptispora hydei | MFLUCC 20–0115T | MT742830 | MT734661 | – | MT767128 |
| Distoseptispora lancangjiangensis | DLUCC 1864T | MW879522 | MW723055 | – | – |
| Distoseptispora leonensis | HKUCC 10822 | DQ408566 | – | – | DQ435089 |
| Distoseptispora lignicola | MFLUCC 18–0198T | MK849797 | MK828651 | – | – |
| Distoseptispora longispora | HFJAU 0705T | MH555357 | MH555359 | – | – |
| Distoseptispora martinii | CGMCC 3.18651 | KX033566 | KU999975 | – | – |
| Distoseptispora meilingensis | JAUCC 4727T | OK562396 | OK562390 | OK562408 | – |
| Distoseptispora meilingensis | JAUCC 4728T | OK562397 | OK562391 | OK562409 | – |
| Distoseptispora multiseptata | MFLUCC 16–1044 | MF077555 | MF077544 | MF135652 | MF135644 |
| Distoseptispora multiseptata | MFLUCC 15–0609T | KX710140 | KX710145 | MF135659 | – |
| Distoseptispora neorostrata | MFLUCC 18–0376T | MN163017 | MN163008 | – | – |
| Distoseptispora nonrostrata | KUNCC 21–10730T | OK341198 | OK310699 | OP413481 | OP413475 |
| Distoseptispora obclavata | MFLUCC 18–0329T | MN163010 | MN163012 | – | – |
| Distoseptispora obpyriformis | DLUCC 0867 | MG979765 | MG979757 | MG988423 | MG988416 |
| Distoseptispora obpyriformis | MFLUCC 17–1694T | MG979764 | – | MG988422 | MG988415 |
| Distoseptispora pachyconidia | KUMCC 21–10724T | OK341194 | OK310696 | OP413477 | OP413471 |
| Distoseptispora palmarum | MFLUCC 18–1446T | MK079663 | MK085062 | MK087660 | MK087670 |
| Distoseptispora phangngaensis | MFLUCC 16–0857T | MF077556 | MF077545 | MF135653 | – |
| Distoseptispora rayongensis | MFLUCC 18–0417 | MH457138 | MH457173 | MH463254 | MH463256 |
| Distoseptispora rayongensis | MFLUCC 18–0415T | MH457137 | MH457172 | MH463253 | MH463255 |
| Distoseptispora rayongensis | MFLU 19–0543 | MN163010 | MN513037 | OP413490 | OP413485 |
| Distoseptispora rostrata | DLUCC 0885 | MG979767 | MG979759 | MG988425 | – |
| Distoseptispora rostrata | MFLUCC 16–0969T | MG979766 | MG979758 | MG988424 | MG988417 |
| Distoseptispora saprophytica | MFLUCC 18–1238T | MW287780 | MW286506 | MW396651 | MW504069 |
| Distoseptispora songkhlaensis | MFLUCC 18–1234T | MW287755 | MW286482 | MW396642 | – |
| Distoseptispora suoluoensis | MFLUCC 17–1305 | MF077558 | MF077547 | – | MZ945510 |
| Distoseptispora suoluoensis | MFLUCC 17–0224T | MF077557 | MF077546 | MF135654 | – |
| Distoseptispora tectonae | MFLUCC 12–0291T | KX751713 | KX751711 | KX751710 | KX751708 |
| Distoseptispora tectonae | MFLUCC 15–0981 | MW287763 | MW286489 | MW396641 | – |
| Distoseptispora tectonae | MFLUCC 16–0946 | MG979768 | MG979760 | MG988426 | MG988418 |
| Distoseptispora tectonae | KUNCC 21–10728 | OK348852 | OK341185 | OP413489 | OP413484 |
| Distoseptispora tectonigena | MFLUCC 12–0292T | KX751714 | KX751712 | – | KX751709 |
| Distoseptispora thailandica | MFLUCC 16–0270T | MH260292 | MH275060 | MH412767 | – |
| Distoseptispora thysanolaenae | KUMCC 18–0182T | MK064091 | MK045851 | MK086031 | – |
| Distoseptispora thysanolaenae | HKAS 112710 | MW879524 | MW723057 | MW729783 | – |
| Distoseptispora verrucosa | GZCC 20–0434T | MZ868762 | MZ868771 | MZ892977 | MZ892983 |
| Distoseptispora xishuangbannaensis | KUMCC 17–0290T | MH260293 | MH275061 | MH412768 | MH412754 |
| Distoseptispora yongxiuensis | JAUCC 4725T | OK562394 | OK562388 | OK562406 | – |
| Distoseptispora yongxiuensis | JAUCC 4726T | OK562395 | OK562389 | OK562407 | – |
| Distoseptispora yunjushanensis | JAUCC 4724T | OK562398 | OK562392 | OK562410 | – |
| Distoseptispora yunjushanensis | JAUCC 4723T | OK562399 | OK562393 | OK562411 | – |
| Distoseptispora yunnanensis | MFLUCC 20–0153T | MW081546 | MW081541 | MW081541 | MW151861 |
| Fluminicola saprophytica | MFLUCC 15–0976T | MF374367 | MF374358 | MF370956 | MF370954 |
| Myrmecridium banksiae | CPC 19852T | JX069855 | JX069871 | – | – |
| Myrmecridium schulzeri | CBS 100.54 | EU041826 | EU041769 | – | – |
| Papulosa amerospora | AFTOL-ID 748 | DQ470950 | – | DQ471069 | DQ470901 |
| Pleurophragmium bambusinum | MFLUCC 12–0850 | KU863149 | KU940161 | KU940213 | – |
| Pseudostanjehughesia aquitropica | MFLUCC 16–0569T | MF077559 | MF077548 | MF135655 | – |
| Pseudostanjehughesia lignicola | MFLUCC 15–0352T | MK849787 | MK828643 | MN194047 | MN124534 |
| Wongia griffinii | DAR 80512T | KU850471 | KU850473 | – | – |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Zhu, R.; Qing, Y.; Yang, H.; Li, C.; Wang, G.; Zhang, D.; Ning, P. Polyphasic Identification of Distoseptispora with Six New Species from Fresh Water. J. Fungi 2022, 8, 1063. https://doi.org/10.3390/jof8101063
Zhang H, Zhu R, Qing Y, Yang H, Li C, Wang G, Zhang D, Ning P. Polyphasic Identification of Distoseptispora with Six New Species from Fresh Water. Journal of Fungi. 2022; 8(10):1063. https://doi.org/10.3390/jof8101063
Chicago/Turabian StyleZhang, Huang, Rong Zhu, Yun Qing, Hao Yang, Chunxue Li, Gennuo Wang, Di Zhang, and Ping Ning. 2022. "Polyphasic Identification of Distoseptispora with Six New Species from Fresh Water" Journal of Fungi 8, no. 10: 1063. https://doi.org/10.3390/jof8101063
APA StyleZhang, H., Zhu, R., Qing, Y., Yang, H., Li, C., Wang, G., Zhang, D., & Ning, P. (2022). Polyphasic Identification of Distoseptispora with Six New Species from Fresh Water. Journal of Fungi, 8(10), 1063. https://doi.org/10.3390/jof8101063