
The Non-Pathogenic Fusarium oxysporum Fo47 Induces Distinct Responses in Two Closely Related Solanaceae Plants against the Pathogen Verticillium dahliae



Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatment Method
2.2. Pathogen Material and Inoculum Preparation
2.3. Inoculation with Verticillium dahliae
2.4. Transcriptomic Profiling
2.5. qPCR Gene Expression
2.6. Statistical Analysis
3. Results
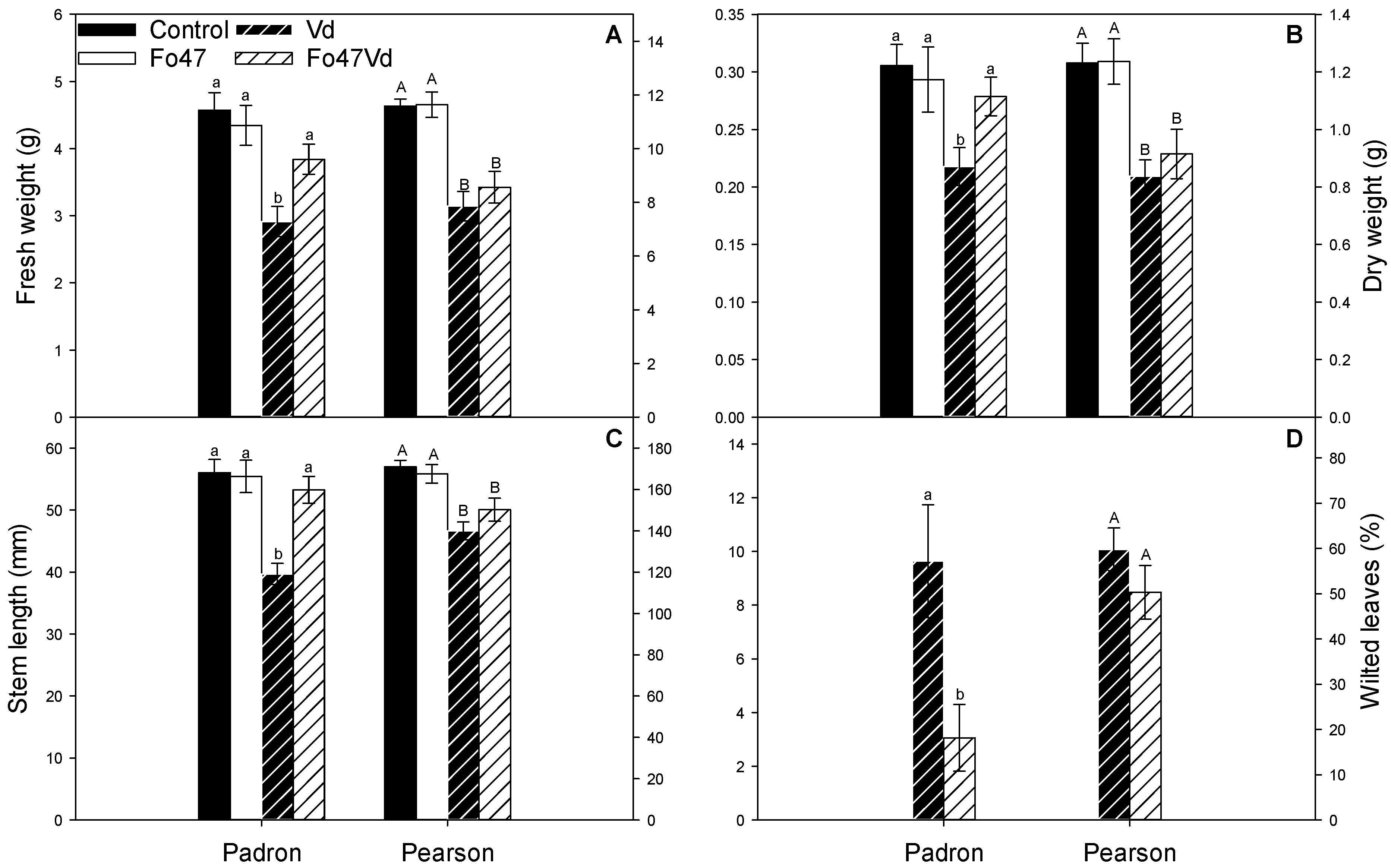
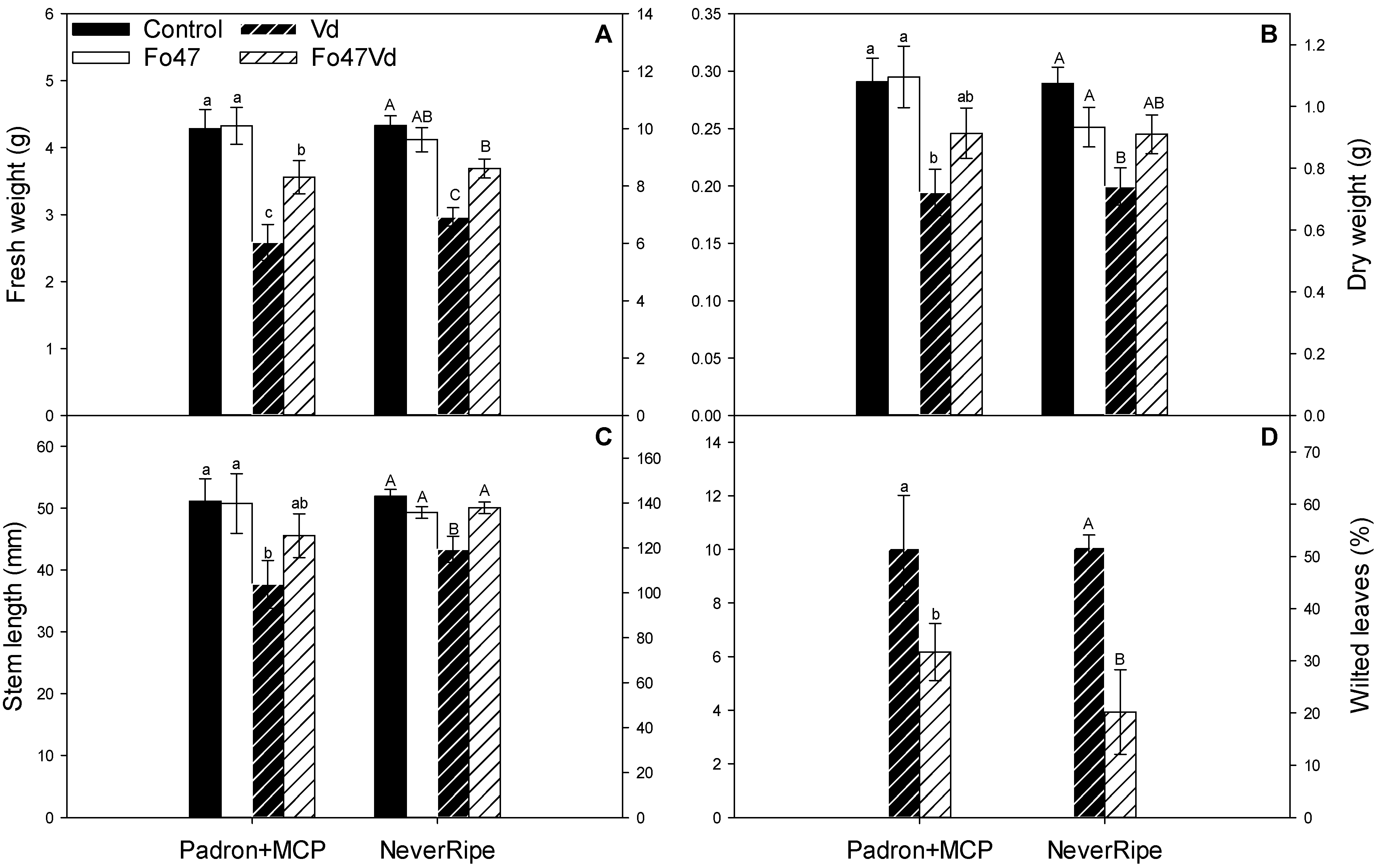
3.1. Fo47 Protects Pepper against V. dahliae but Not Tomato
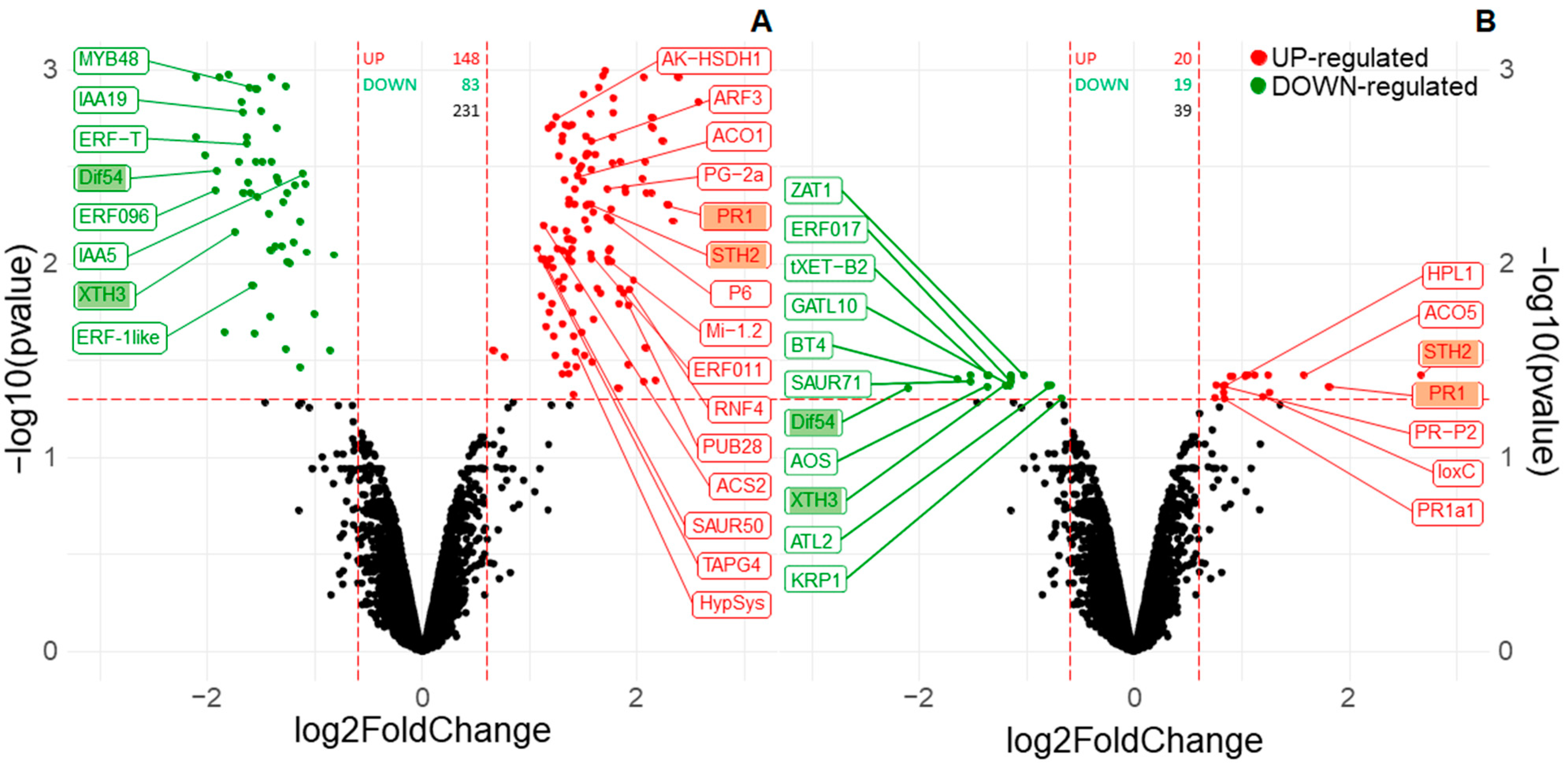
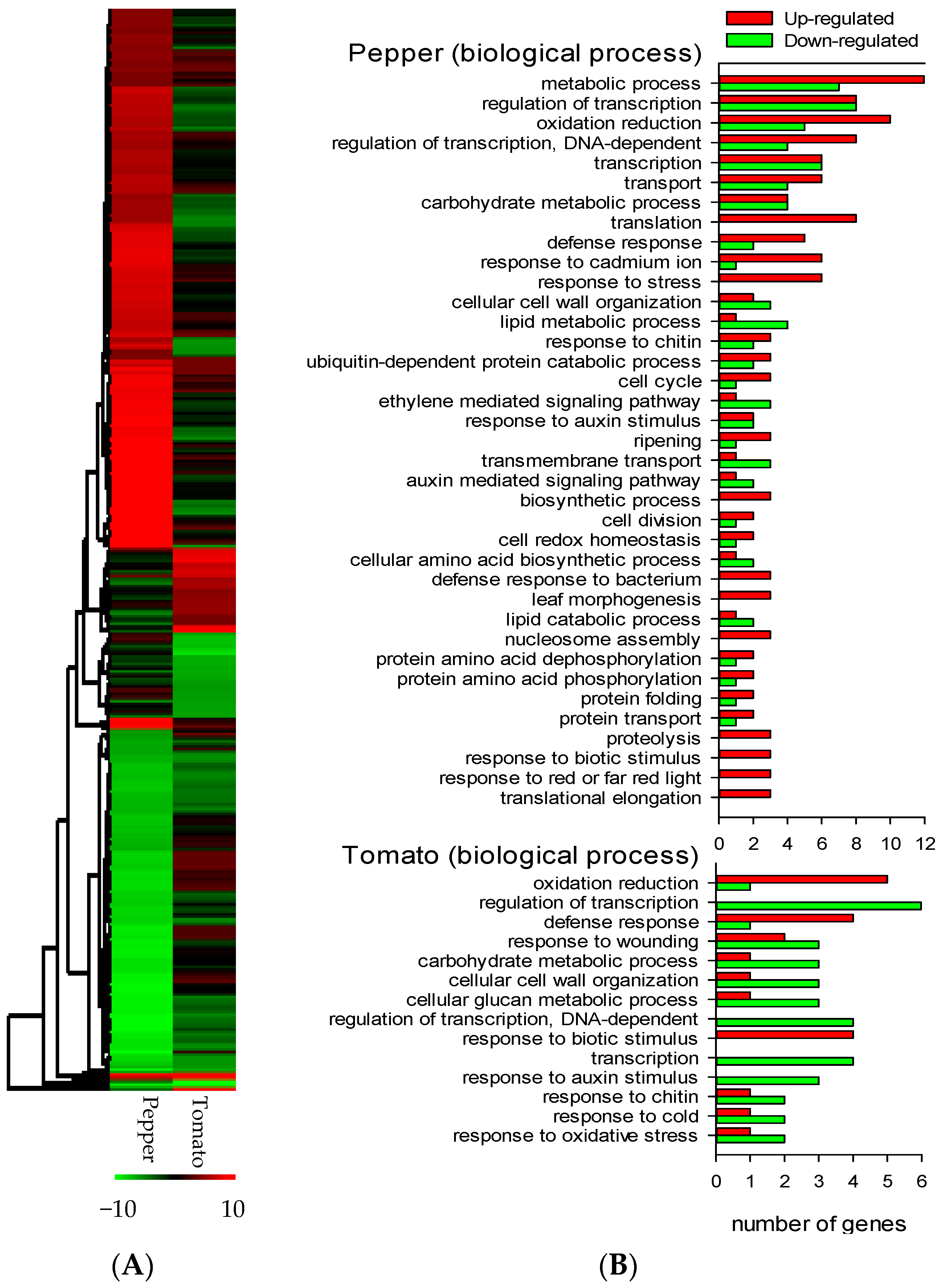
3.2. Fo47 Triggers Different Transcriptome Reprogramming in Pepper and Tomato but Both Include Ethyelene-Related Genes
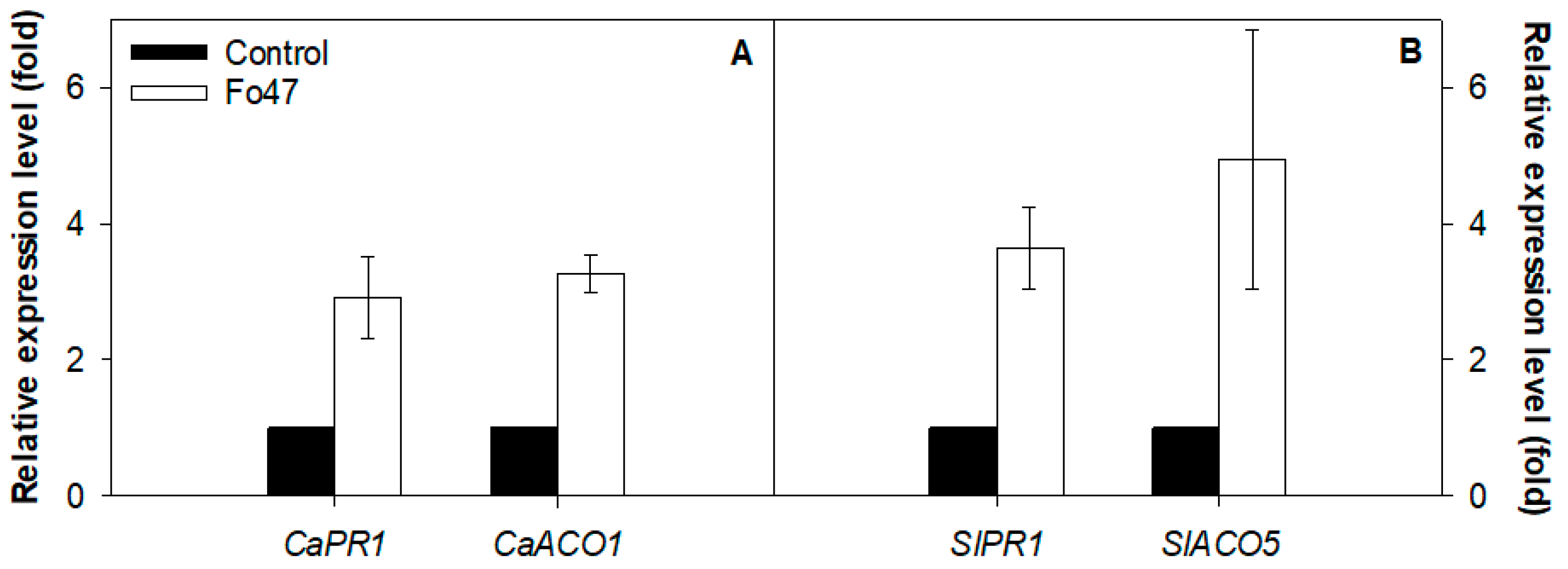
3.3. Ethylene Role in Fo47-Induced Resistance Depends on the Host
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fuchs, J.-G.; Moënne-Loccoz, Y.; Défago, G. Nonpathogenic Fusarium oxysporum Strain Fo47 Induces Resistance to Fusarium Wilt in Tomato. Plant Dis. 1997, 81, 492–496. [Google Scholar] [CrossRef]
- Duijff, B.J.; Pouhair, D.; Olivain, C.; Alabouvette, C.; Lemanceau, P. Implication of Systemic Induced Resistance in the Suppression of Fusarium Wilt of Tomato by Pseudomonas fluorescens WCS417r and by Nonpathogenic Fusarium oxysporum Fo47. Eur. J. Plant Pathol. 1998, 104, 903–910. [Google Scholar] [CrossRef]
- Larkin, R.P.; Fravel, D.R. Mechanisms of Action and Dose-Response Relationships Governing Biological Control of Fusarium Wilt of Tomato by Nonpathogenic Fusarium spp. Phytopathology 1999, 89, 1152–1161. [Google Scholar] [CrossRef] [PubMed]
- Elmer, W.H. Combining nonpathogenic strains of Fusarium oxysporum with sodium chloride to suppress fusarium crown rot of asparagus in replanted fields. Plant Pathol. 2004, 53, 751–758. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, J.; Jia, R.; Ma, Q.; Zong, Z.; Wang, Y. Suppression of plant wilt diseases by nonpathogenic Fusarium oxysporum Fo47 combined with actinomycete strains. Biocontrol. Sci. Technol. 2018, 28, 562–573. [Google Scholar] [CrossRef]
- Aimé, S.; Cordier, C.; Alabouvette, C.; Olivain, C. Comparative analysis of PR gene expression in tomato inoculated with virulent Fusarium oxysporum f. sp. lycopersici and the biocontrol strain F. oxysporum Fo47. Physiol. Mol. Plant Pathol. 2008, 73, 9–15. [Google Scholar] [CrossRef]
- Aimé, S.; Alabouvette, C.; Steinberg, C.; Olivain, C. The Endophytic Strain Fusarium oxysporum Fo47: A Good Candidate for Priming the Defense Responses in Tomato Roots. Mol. Plant Microbe Interact. 2013, 26, 918–926. [Google Scholar] [CrossRef]
- Kavroulakis, N.; Ntougias, S.; Zervakis, G.I.; Ehaliotis, C.; Haralampidis, K.; Papadopoulou, K.K. Role of ethylene in the protection of tomato plants against soil-borne fungal pathogens conferred by an endophytic Fusarium solani strain. J. Exp. Bot. 2007, 58, 3853–3864. [Google Scholar] [CrossRef]
- Constantin, M.E.; De Lamo, F.J.; Vlieger, B.V.; Rep, M.; Takken, F.L.W. Endophyte-Mediated Resistance in Tomato to Fusarium oxysporum Is Independent of ET, JA, and SA. Front. Plant Sci. 2019, 10, 979. [Google Scholar] [CrossRef]
- Veloso, J.; Alabouvette, C.; Olivain, C.; Flors, V.; Pastor, V.; García, T.; Díaz, J. Modes of action of the protective strain Fo47 in controlling verticillium wilt of pepper. Plant Pathol. 2015, 65, 997–1007. [Google Scholar] [CrossRef]
- Zhang, W.; Corwin, J.A.; Copeland, D.; Feusier, J.; Eshbaugh, R.; Chen, F.; Atwell, S.; Kliebenstein, D.J. Plastic Transcriptomes Stabilize Immunity to Pathogen Diversity: The Jasmonic Acid and Salicylic Acid Networks within the Arabidopsis/Botrytis Pathosystem. Plant Cell 2017, 29, 2727–2752. [Google Scholar] [CrossRef]
- Achuo, E.A.; Audenaert, K.; Meziane, H.; Hofte, M. The salicylic acid-dependent defence pathway is effective against different pathogens in tomato and tobacco. Plant Pathol. 2004, 53, 65–72. [Google Scholar] [CrossRef]
- Glazebrook, J. Contrasting Mechanisms of Defense Against Biotrophic and Necrotrophic Pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef] [PubMed]
- Dhar, N.; Chen, J.-Y.; Subbarao, K.V.; Klosterman, S.J. Hormone Signaling and Its Interplay With Development and Defense Responses in Verticillium-Plant Interactions. Front. Plant Sci. 2020, 11, 1. [Google Scholar] [CrossRef]
- Fradin, E.F.; Zhang, Z.; Ayala, J.C.J.; Castroverde, C.D.; Nazar, R.N.; Robb, J.; Liu, C.-M.; Thomma, B.P. Genetic Dissection of Verticillium Wilt Resistance Mediated by Tomato Ve1. Plant Physiol. 2009, 150, 320–332. [Google Scholar] [CrossRef]
- Barchenger, D.W.; Rodriguez, K.; Jiang, L.; Hanson, S.F.; Bosland, P.W. Allele-specific CAPS marker in a Ve1 homolog of Capsicum annuum for improved selection of Verticillium dahliae resistance. Mol. Breed. 2017, 37, 134. [Google Scholar] [CrossRef]
- Robison, M.M.; Griffith, M.; Pauls, K.P.; Glick, B.R. Dual Role for ethylene in susceptibility of tomato to Verticillium Wilt. J. Phytopathol. 2001, 149, 385–388. [Google Scholar] [CrossRef]
- Kim, Y.J.; Hwang, B.K. Pepper gene encoding a basic pathogenesis-related 1 protein is pathogen and ethylene inducible. Physiol. Plant. 2000, 108, 51–60. [Google Scholar] [CrossRef]
- Eyal, Y.; Meller, Y.; Lev-Yadun, S.; Fluhr, R. A basic-type PR-1 promoter directs ethylene responsiveness, vascular and abscission zone-specific expression. Plant J. 1993, 4, 225–234. [Google Scholar] [CrossRef]
- Tornero, P.; Gadea, J.; Conejero, V.; Vera, P. Two PR-1 Genes from Tomato Are Differentially Regulated and Reveal a Novel Mode of Expression for a Pathogenesis-Related Gene During the Hypersensitive Response and Development. Mol. Plant Microbe Interact. 1997, 10, 624–634. [Google Scholar] [CrossRef]
- Schenk, P.M.; Kazan, K.; Wilson, I.; Anderson, J.P.; Richmond, T.; Somerville, S.C.; Manners, J.M. Coordinated plant defense responses in Arabidopsis revealed by microarray analysis. Proc. Natl. Acad. Sci. USA 2000, 97, 11655–11660. [Google Scholar] [CrossRef]
- Diaz, J.; Silvar, C.; Varela, M.M.; Bernal, A.; Merino, F. Fusarium confers protection against several mycelial pathogens of pepper plants. Plant Pathol. 2005, 54, 773–780. [Google Scholar] [CrossRef]
- Díaz, J.; Have, A.T.; Van Kan, J.A. The Role of Ethylene and Wound Signaling in Resistance of Tomato to Botrytis cinerea. Plant Physiol. 2002, 129, 1341–1351. [Google Scholar] [CrossRef] [PubMed]
- Novo, M.; Pomar, F.; Gayoso, C.; Merino, F. Cellulase Activity in Isolates of Verticillium dahliae Differing in Aggressiveness. Plant Dis. 2006, 90, 155–160. [Google Scholar] [CrossRef]
- de Cáceres, F.M.; Varela, J.D.; García, A.S. Virulence of different isolates causing verticillium wilt pepper in the Padrón region. Span. J. Agric. Res. 2003, 1, 53–58. [Google Scholar]
- MAQC-Consortium; Shi, L.; Reid, L.H.; Jones, W.D.; Shippy, R.; Warrington, J.A.; Baker, S.C.; Collins, P.J.; Longueville, F.d.; Kawasaki, E.S.; et al. The MicroArray Quality Control (MAQC) project shows inter- and intraplatform reproducibility of gene expression measurements. Nat. Biotechnol. 2006, 24, 1151–1161. [Google Scholar]
- Klaus, B.; Reisenauer, S. An end to end workflow for differential gene expression using Affymetrix microarrays. F1000Research 2016, 5, 1384. [Google Scholar] [CrossRef]
- Silvar, C.; Merino, F.; Díaz, J. Differential activation of defense-related genes in susceptible and resistant pepper cultivars infected with Phytophthora capsici. J. Plant Physiol. 2008, 165, 1120–1124. [Google Scholar] [CrossRef]
- Silvar, C.; Merino, F.; Díaz, J. Resistance in pepper plants induced by Fusarium oxysporum f. sp lycopersici involves different defence-related genes. Plant Biol. 2009, 11, 68–74. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Dytham, C. Choosing and Using Statistics: A Biologist’s Guide; Wiley-Blackwell: Hoboken, NJ, USA, 1999. [Google Scholar]
- Llorente, F.; Muskett, P.; Sánchez-Vallet, A.; López, G.; Ramos, B.; Sánchez-Rodríguez, C.; Jordá, L.; Parker, J.; Molina, A. Repression of the Auxin Response Pathway Increases Arabidopsis Susceptibility to Necrotrophic Fungi. Mol. Plant 2008, 1, 496–509. [Google Scholar] [CrossRef] [PubMed]
- Hackett, R.M.; Ho, C.-W.; Lin, Z.; Foote, H.C.; Fray, R.G.; Grierson, D. Antisense Inhibition of the Nr Gene Restores Normal Ripening to the Tomato Never-ripe Mutant, Consistent with the Ethylene Receptor- Inhibition Model. Plant Physiol. 2000, 124, 1079–1086. [Google Scholar] [CrossRef]
- O’Donnell, P.J.; Jones, J.B.; Antoine, F.R.; Ciardi, J.; Klee, H.J. Ethylene-dependent salicylic acid regulates an expanded cell death response to a plant pathogen. Plant J. 2001, 25, 315–323. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, P.J.; Schmelz, E.A.; Moussatche, P.; Lund, S.T.; Jones, J.B.; Klee, H.J. Susceptible to intolerance—A range of hormonal actions in a susceptible Arabidopsis pathogen response. Plant J. 2003, 33, 245–257. [Google Scholar] [CrossRef]
- Pegg, G. The response of ethylene-treated tomato plants to infection by Verticillium albo-atrum. Physiol. Plant Pathol. 1976, 9, 215–226. [Google Scholar] [CrossRef]
- Lund, S.T.; Stall, R.E.; Klee, H.J. Ethylene regulates the susceptible response to pathogen infection in tomato. Plant Cell 1998, 10, 371–382. [Google Scholar] [CrossRef]
- Cronshaw, D.; Pegg, G. Ethylene as a toxin synergist in Verticillium wilt of tomato. Physiol. Plant Pathol. 1976, 9, 33–44. [Google Scholar] [CrossRef]
- VanderMolen, G.E.; Labavitch, J.M.; Strand, L.L.; DeVay, J.E. Pathogen-induced vascular gels: Ethylene as a host intermediate. Physiol. Plant. 1983, 59, 573–580. [Google Scholar] [CrossRef]
- Pantelides, I.S.; Tjamos, S.E.; Striglis, I.A.; Chatzipavlidis, I.; Paplomatas, E.J. Mode of action of a non-pathogenic Fusarium oxysporum strain against Verticillium dahliae using Real Time QPCR analysis and biomarker transformation. Biol. Control 2009, 50, 30–36. [Google Scholar] [CrossRef]
- Robison, M.M.; Shah, S.; Tamot, B.; Pauls, K.P.; Moffatt, B.A.; Glick, B.R. Reduced symptoms of Verticillium wilt in transgenic tomato expressing a bacterial ACC deaminase. Mol. Plant Pathol. 2001, 2, 135–145. [Google Scholar] [CrossRef]
- Thaler, J.S.; Owen, B.; Higgins, V.J. The Role of the Jasmonate Response in Plant Susceptibility to Diverse Pathogens with a Range of Lifestyles. Plant Physiol. 2004, 135, 530–538. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Bel, P.; Sanmartín, N.; Pastor, V.; Mateu, D.; Cerezo, M.; Vidal-Albalat, A.; Pastor-Fernández, J.; Pozo, M.J.; Flors, V. Mycorrhizal tomato plants fine tunes the growth-defence balance upon N depleted root environments. Plant Cell Environ. 2018, 41, 406–420. [Google Scholar] [CrossRef]
- Meents, A.K.; Furch, A.C.U.; Almeida-Trapp, M.; Özyürek, S.; Scholz, S.S.; Kirbis, A.; Lenser, T.; Theißen, G.; Grabe, V.; Hansson, B.; et al. Beneficial and Pathogenic Arabidopsis Root-Interacting Fungi Differently Affect Auxin Levels and Responsive Genes During Early Infection. Front. Microbiol. 2019, 10, 380. [Google Scholar] [CrossRef]
- Veloso, J.F.C.A.; Díaz, J. Fusarium oxysporum Fo47 confers protection to pepper plants against Verticillium dahliae and Phytophthora capsici, and induces the expression of defence genes. Plant Pathol. 2011, 61, 281–288. [Google Scholar] [CrossRef]
- Ament, K.; Van Schie, C.C.; Bouwmeester, H.J.; Haring, M.A.; Schuurink, R.C. Induction of a leaf specific geranylgeranyl pyrophosphate synthase and emission of (E,E)-4,8,12-trimethyltrideca-1,3,7,11-tetraene in tomato are dependent on both jasmonic acid and salicylic acid signaling pathways. Planta 2006, 224, 1197–1208. [Google Scholar] [CrossRef]
- Van Schie, C.C.N.; Haring, M.A.; Schuurink, R.C. Tomato linalool synthase is induced in trichomes by jasmonic acid. Plant Mol. Biol. 2007, 64, 251–263. [Google Scholar] [CrossRef]
- Cameron, R.K.; Paiva, N.L.; Lamb, C.J.; Dixon, R.A. Accumulation of salicylic acid and PR-1 gene transcripts in relation to the systemic acquired resistance (SAR) response induced by Pseudomonas syringae pv. tomato in Arabidopsis. Physiol. Mol. Plant Pathol. 1999, 55, 121–130. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. (Eds.) Plant Physiology, 5th ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2010; p. 782. [Google Scholar]
- Grunewald, W.; Vanholme, B.; Pauwels, L.; Plovie, E.; Inzé, D.; Gheysen, G.; Goossens, A. Expression of the Arabidopsis jasmonate signalling repressor JAZ1/TIFY10A is stimulated by auxin. EMBO Rep. 2009, 10, 923–928. [Google Scholar] [CrossRef]
- Nagpal, P.; Ellis, C.M.; Weber, H.; Ploense, S.E.; Barkawi, L.S.; Guilfoyle, T.J.; Hagen, G.; Alonso, J.M.; Cohen, J.D.; Farmer, E.E.; et al. Auxin response factors ARF6 and ARF8 promote jasmonic acid production and flower maturation. Development 2005, 132, 4107–4118. [Google Scholar] [CrossRef]
- Dombrecht, B.; Xue, G.P.; Sprague, S.J.; Kirkegaard, J.A.; Ross, J.J.; Reid, J.B.; Fitt, G.P.; Sewelam, N.; Schenk, P.M.; Manners, J.M.; et al. MYC2 Differentially Modulates Diverse Jasmonate-Dependent Functions in Arabidopsis. Plant Cell 2007, 19, 2225–2245. [Google Scholar] [CrossRef]




 : up-regulated,
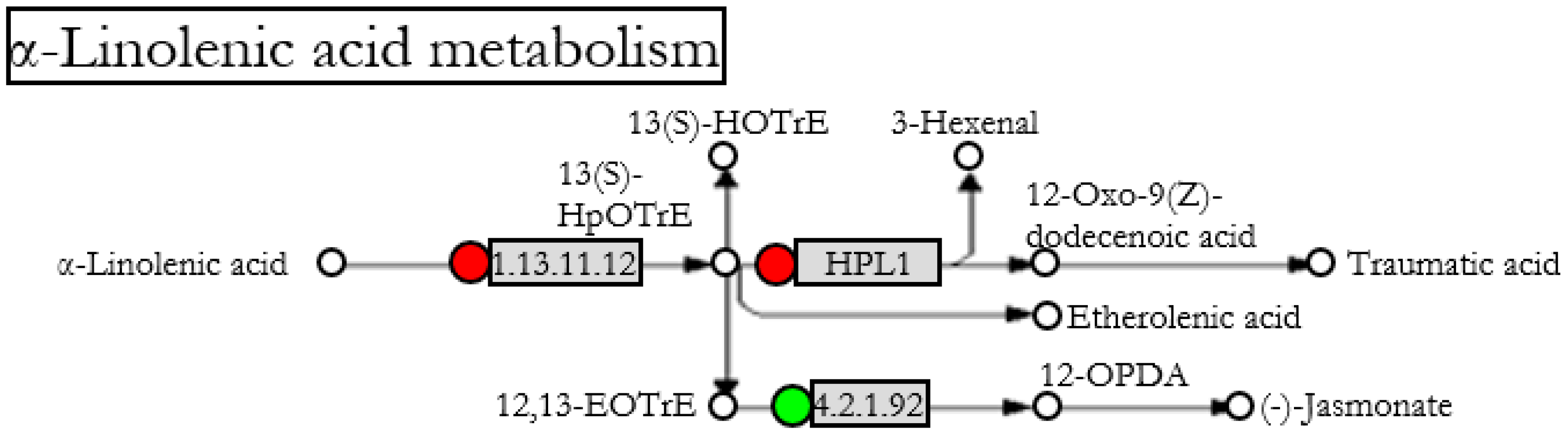
: up-regulated,  : down-regulated) after Fo47 treatment. The pathway has been simplified from the original in KEGG databases (accession: SLY00592).
: up-regulated, : down-regulated) after Fo47 treatment. The pathway has been simplified from the original in KEGG databases (accession: SLY00592).
: down-regulated) after Fo47 treatment. The pathway has been simplified from the original in KEGG databases (accession: SLY00592).
: up-regulated, : down-regulated) after Fo47 treatment. The pathway has been simplified from the original in KEGG databases (accession: SLY00592).
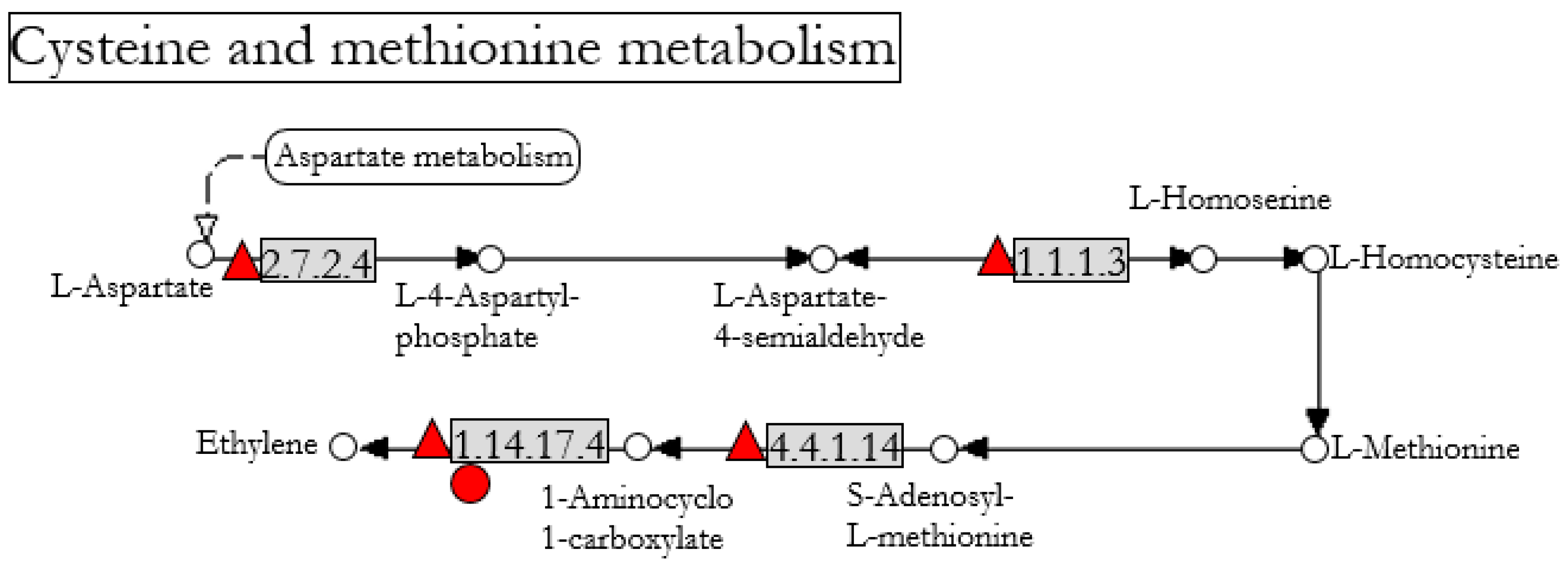
 : up-regulated) and tomato (
: up-regulated) and tomato ( : up-regulated) after Fo47 treatment. The pathway has been simplified from the original in KEGG databases (accession: CANN00270).
: up-regulated) and tomato (: up-regulated) after Fo47 treatment. The pathway has been simplified from the original in KEGG databases (accession: CANN00270).
: up-regulated) after Fo47 treatment. The pathway has been simplified from the original in KEGG databases (accession: CANN00270).
: up-regulated) and tomato (: up-regulated) after Fo47 treatment. The pathway has been simplified from the original in KEGG databases (accession: CANN00270).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene | Fold | Description |
|---|---|---|---|
| 107864567 778321 | STH2 | 2.57 (Pepper) 6.24 (Tomato) | Pathogenesis-related protein STH-2-like |
| 107840155 100191111 | PR1 | 3.41 (Pepper) 4.42 (Tomato) | Basic form of pathogenesis-related protein 1-like |
| 107877508 101245668 | XET24 | 0.23 (Pepper) 0.30 (Tomato) | Xyloglucan endotransglucosylase/hydrolase protein 24-like |
| 107873721 544295 | Dif54 | 0.24 (Pepper) 0.10 (Tomato) | Extensin-like protein |
| Gene ID | Gene | Organism | Fold | Description |
|---|---|---|---|---|
| 544083 | PR-1a1 | Tomato | 2.27 | Solanum lycopersicum PR-1a1 |
| 107864567778321 | STH2 | Pepper Tomato | 2.57 6.24 | Pathogenesis-related protein STH-2-like |
| 544069 | PR-P2 | Tomato | 2.14 | Solanum lycopersicum PR-P2 |
| 107840155 100191111 | PR1 | Pepper Tomato | 3.41 4.42 | Basic form of pathogenesis-related protein 1 |
| 107842907 | P6 | Pepper | 2.25 | Pathogenesis-related protein P6 |
| Gene ID | Gene | Organism | Fold | Description |
|---|---|---|---|---|
| 107839239 | ACS2 | Pepper | 3.39 | 1-aminocyclopropane-1-carboxylate synthase |
| 107860267 | ERF011 | Pepper | 2.97 | Ethylene-responsive transcription factor |
| 107853805 | ACO1 | Pepper | 2.73 | 1-aminocyclopropane-1-carboxylate oxidase |
| 107872603 | ERF-1like | Pepper | 0.33 | Ethylene-responsive transcription factor |
| 107865816 | ERF-T | Pepper | 0.32 | Ethylene-responsive transcription factor |
| 107866610 | ERF096 | Pepper | 0.26 | Ethylene-responsive transcription factor |
| 543800 | ACO5 | Tomato | 3.40 | 1-aminocyclopropane-1-carboxylate oxidase |
| 101253257 | ERF017 | Tomato | 0.34 | Ethylene-responsive transcription factor |
| Gene ID | Gene | Organism | Fold | Description |
|---|---|---|---|---|
| 778363 | ARF3 | Pepper | 3.09 | Auxin response factor 3 |
| 101256828 | SAUR50 | Pepper | 2.11 | SAUR-like auxin-responsive protein |
| 101055544 | IAA5 | Pepper | 0.46 | Auxin-responsive protein |
| 101055549 | IAA19 | Pepper | 0.31 | Auxin-responsive protein |
| 101251823 | KRP1 | Tomato | 0.46 | Calcium-binding protein KRP1 |
| 101265243 | SAUR71 | Tomato | 0.50 | SAUR-like auxin-responsive protein |
| 101259898 | BT4 | Tomato | 0.48 | BTB and TAZ domain protein 4 |
| Gene ID | EC | Gene | Organism | Fold | Description |
|---|---|---|---|---|---|
| 544008 | 1.13.11.12 | loxC | Tomato | 2.31 | Lipoxygenase |
| 543642 | HPL1 | HPL1 | Tomato | 2.09 | Fatty acid hydroperoxide lyase |
| 606711 | 4.2.1.92 | AOS | Tomato | 0.28 | Allene oxide synthase |
| Gene ID | EC | Gene | Organism | Fold | Description |
|---|---|---|---|---|---|
| 107879648 | 2.7.2.4 1.1.1.3 | AK-HSDH1 | Pepper | 3.52 | Bifunctional aspartokinase/homoserine dehydrogenase 1 |
| 107839239 | 4.4.1.14 | ACS2 | Pepper | 3.39 | 1-aminocyclopropane-1-carboxylate synthase |
| 107853805 | 1.14.17.4 | ACO1 | Pepper | 2.73 | 1-aminocyclopropane-1-carboxylate oxidase |
| 543800 | 1.14.17.4 | ACO5 | Tomato | 2.97 | 1-aminocyclopropane-1-carboxylate oxidase |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veloso, J.; Díaz, J. The Non-Pathogenic Fusarium oxysporum Fo47 Induces Distinct Responses in Two Closely Related Solanaceae Plants against the Pathogen Verticillium dahliae. J. Fungi 2021, 7, 344. https://doi.org/10.3390/jof7050344
Veloso J, Díaz J. The Non-Pathogenic Fusarium oxysporum Fo47 Induces Distinct Responses in Two Closely Related Solanaceae Plants against the Pathogen Verticillium dahliae. Journal of Fungi. 2021; 7(5):344. https://doi.org/10.3390/jof7050344
Chicago/Turabian StyleVeloso, Javier, and José Díaz. 2021. "The Non-Pathogenic Fusarium oxysporum Fo47 Induces Distinct Responses in Two Closely Related Solanaceae Plants against the Pathogen Verticillium dahliae" Journal of Fungi 7, no. 5: 344. https://doi.org/10.3390/jof7050344
APA StyleVeloso, J., & Díaz, J. (2021). The Non-Pathogenic Fusarium oxysporum Fo47 Induces Distinct Responses in Two Closely Related Solanaceae Plants against the Pathogen Verticillium dahliae. Journal of Fungi, 7(5), 344. https://doi.org/10.3390/jof7050344