
White-Nose Syndrome Confirmed in Italy: A Preliminary Assessment of Its Occurrence in Bat Species

 , , ,
, , ,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
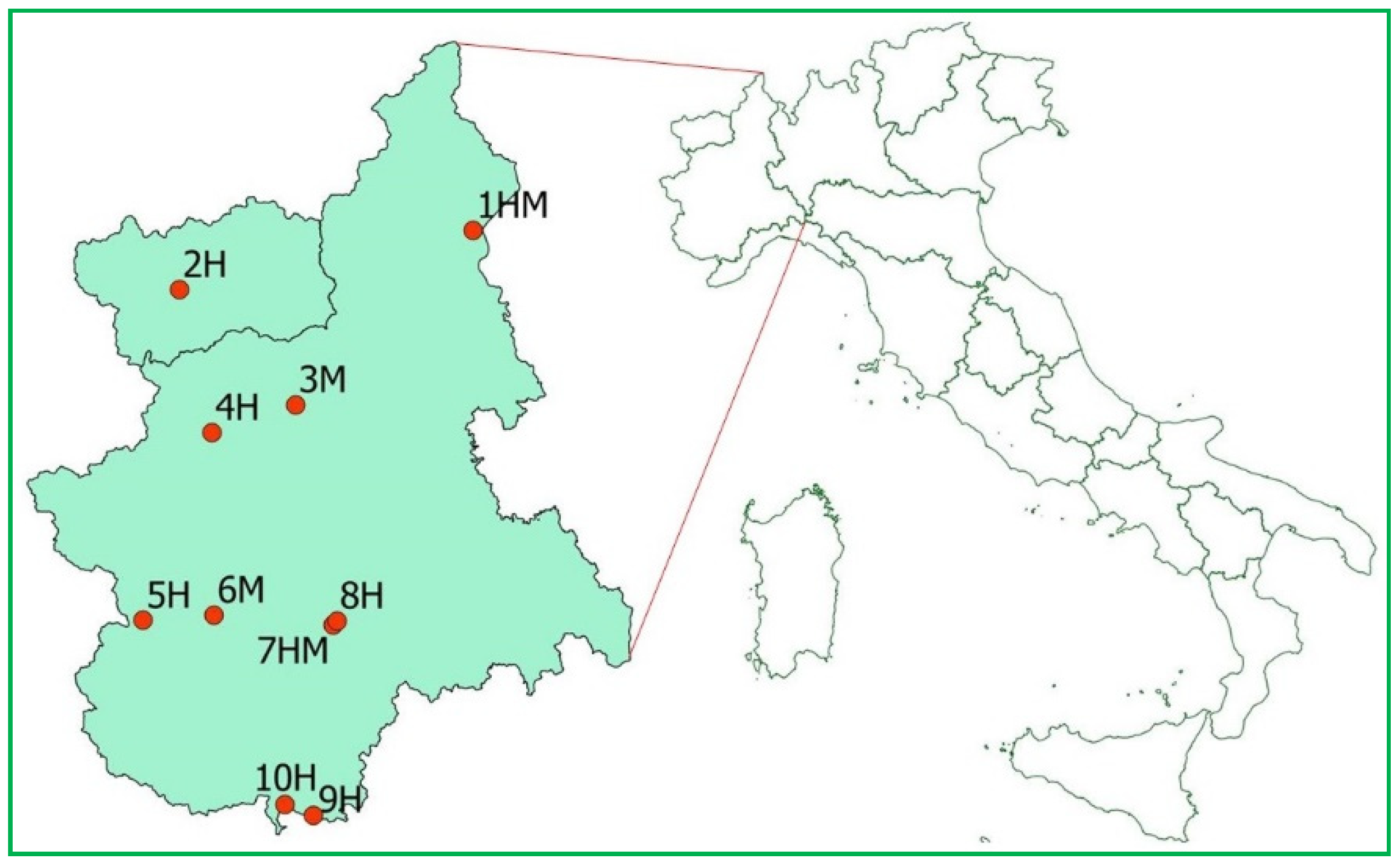
2.1. Sample Collection
2.2. Fungal Isolation and Identification
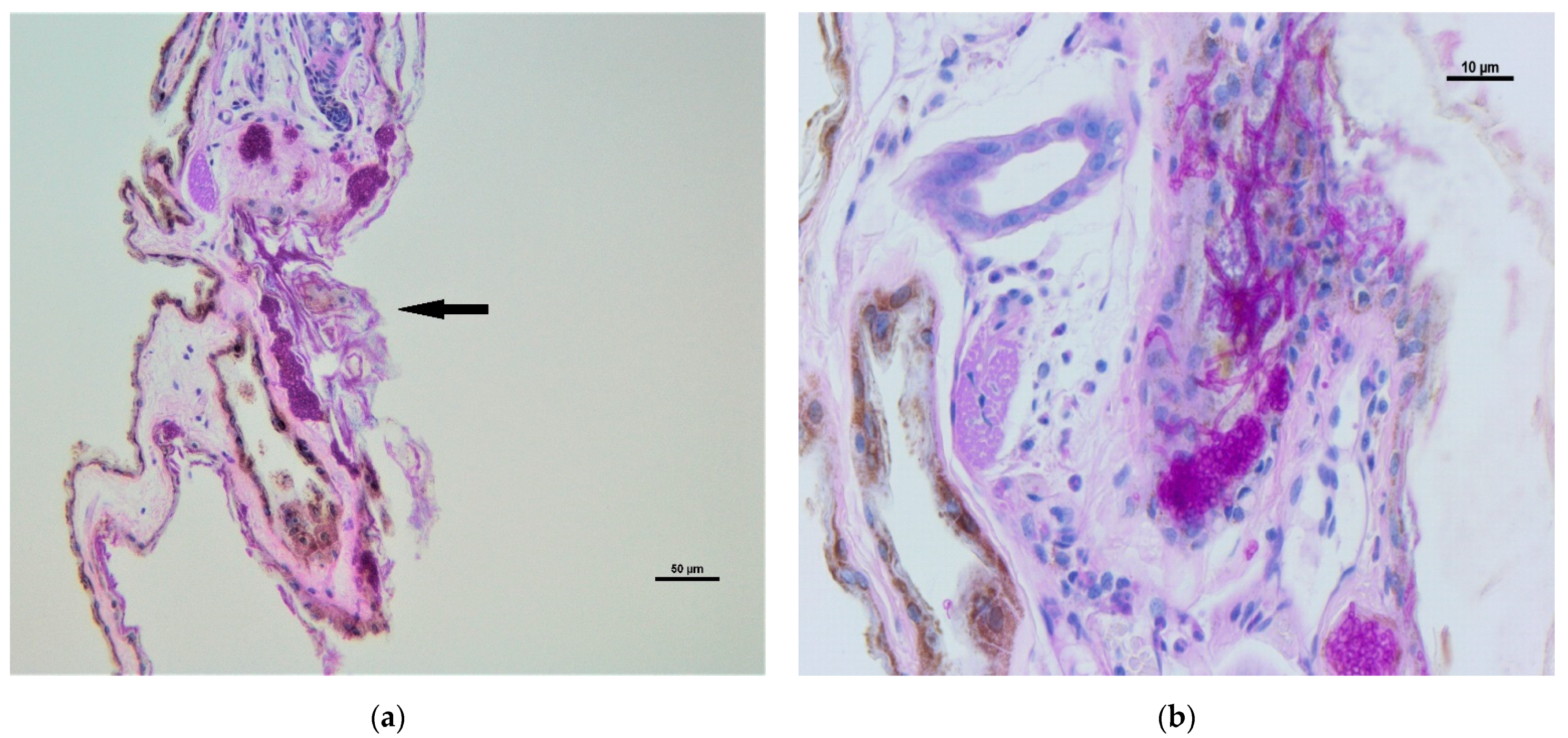
2.3. Histopathological Analyses
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hoyt, J.R.; Kilpatrick, A.M.; Langwig, K.E. Ecology and impacts of white-nose syndrome on bats. Nat. Rev. Genet. 2021, 19, 196–210. [Google Scholar] [CrossRef] [PubMed]
- Farina, L.L.; Lankton, J.S. Chiroptera. In Pathology of Wildlife and Zoo Animals; Terio, K., McAloose, D., St. Leger, J., Eds.; Academic Press: Cambridge, UK, 2018; pp. 607–633. [Google Scholar]
- Blehert, D.S.; Hicks, A.C.; Behr, M.; Meteyer, C.U.; Berlowski-Zier, B.M.; Buckles, E.L.; Coleman, J.T.H.; Darling, S.R.; Gargas, A.; Niver, R.; et al. Bat White-Nose Syndrome: An Emerging Fungal Pathogen? Science 2009, 323, 227. [Google Scholar] [CrossRef]
- Gargas, A.; Trest, M.; Christensen, M.; Volk, T.; Blehert, D. Geomyces destructans sp. nov. associated with bat white-nose syndrome. Mycotaxon 2009, 108, 147–154. [Google Scholar] [CrossRef]
- Meteyer, C.U.; Verant, M.L. White-Nose Syndrome: Cutaneous Invasive Ascomycosis in Hibernating Bats. In Fowler’s Zoo and Wild Animal Medicine Current Therapy; Miller, R.E., Lamberski, N., Calle, P.P., Eds.; Elsevier: St. Louis, MO, USA, 2019; Volume 9, pp. 507–513. [Google Scholar]
- Pikula, J.; Amelon, S.K.; Bandouchova, H.; Bartonička, T.; Berkova, H.; Brichta, J.; Hoper, S.; Kokurewicz, T.; Kolarik, M.; Köllner, B.; et al. White-nose syndrome pathology grading in Nearctic and Palearctic bats. PLoS ONE 2017, 12, e0180435. [Google Scholar] [CrossRef] [PubMed]
- Zukal, J.; Bandouchova, H.; Brichta, J.; Cmokova, A.; Jaron, K.S.; Kolarik, M.; Karacova., V.; Kubátová, A.; Nováková, A.; Orlov, O.; et al. White-nose syndrome without borders: Pseudogymnoascus destructans infection tolerated in Europe and Palearctic Asia but not in North America. Sci. Rep. 2016, 6, 19829. [Google Scholar] [CrossRef] [PubMed]
- Pikula, J.; Bandouchova, H.; Novotný, L.; Meteyer, C.U.; Zukal, J.; Irwin, N.R.; Zima, J.; Martínková, N. Histopathology con-firms white-nose syndrome in bats in Europe. J. Wildl. Dis. 2012, 48, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Wibbelt, G.; Puechmaille, S.J.; Ohlendorf, B.; Mühldorfer, K.; Bosch, T.; Görföl, T.; Passior, K.; Kurth, A.; Lacremans, D.; Forget, F. Skin Lesions in European Hibernating Bats Associated with Geomyces destructans, the Etiologic Agent of White-Nose Syndrome. PLoS ONE 2013, 8, e74105. [Google Scholar] [CrossRef]
- Cryan, P.M.; Meteyer, C.U.; Boyles, J.G.; Blehert, D.S. Wing pathology of white-nose syndrome in bats suggests life-threatening disruption of physiology. BMC Biol. 2010, 8, 135. [Google Scholar] [CrossRef]
- Reeder, D.M.; Frank, C.L.; Turner, G.G.; Meteyer, C.U.; Kurta, A.; Britzke, E.R.; Vodzak, M.E.; Darlin, S.R.; Stihler, C.W.; Hicks, A.C.; et al. Frequent arousal from hibernation linked to se-verity of infection and mortality in bats with white-nose syndrome. PLoS ONE 2012, 7, e38920. [Google Scholar] [CrossRef]
- Warnecke, L.; Turner, J.M.; Bollinger, T.K.; Misra, V.; Cryan, P.M.; Blehert, D.S.; Wibbelt, G.; Willis, C.K.R. Pathophysiology of white-nose syndrome in bats: A mechanistic model linking wing damage to mortality. Biol. Lett. 2013, 9, 20130177. [Google Scholar] [CrossRef]
- Verant, M.L.; Meteyer, C.U.; Speakman, J.R.; Cryan, P.M.; Lorch, J.M.; Blehert, D.S. White-nose syndrome initiates a cascade of physiologic disturbances in the hibernating bat host. BMC Physiol. 2014, 14, 1–11. [Google Scholar] [CrossRef]
- Frick, W.F.; Puechmaille, S.J.; Willis, C.K. White-nose syndrome in bats. In Bats in the Anthropocene: Conservation of Bats in a Changing World; Voigt, C., Kingston, T., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2016; pp. 245–262. [Google Scholar]
- McGuire, L.P.; Mayberry, H.W.; Willis, C.K.R. White-nose syndrome increases torpid metabolic rate and evaporative water loss in hibernating bats. Am. J. Physiol. Integr. Comp. Physiol. 2017, 313, R680–R686. [Google Scholar] [CrossRef]
- Meteyer, C.U.; Buckles, E.L.; Blehert, D.S.; Hicks, A.C.; Green, D.E.; Shearn-Bochsler, V.; Thomas, N.J.; Gargas, A.; Behr, M.J. His-topathologic criteria to confirm white-nose syndrome in bats. J. Vet. Diagn. Investig. 2009, 21, 411–414. [Google Scholar] [CrossRef]
- Leopardi, S.; Blake, D.; Puechmaille, S.J. White-Nose Syndrome fungus introduced from Europe to North America. Curr. Biol. 2015, 25, R217–R219. [Google Scholar] [CrossRef] [PubMed]
- Drees, K.P.; Lorch, J.M.; Puechmaille, S.J.; Parise, K.L.; Wibbelt, G.; Hoyt, J.R.; Sun, K.; Jargalsaikhan, A.; Dalannast, M.; Palmer, J.M.; et al. Phylogenetics of a Fungal Invasion: Origins and Widespread Dispersal of White-Nose Syndrome. mBio 2017, 8, e01941-17. [Google Scholar] [CrossRef] [PubMed]
- Wibbelt, G.; Kurth, A.; Hellmann, D.; Weishaar, M.; Barlow, A.; Veith, M.; Prüger, J.; Görföl, T.; Grosche, L.; Bontadina, F.; et al. White-Nose Syndrome Fungus (Geomyces destructans) in Bats, Europe. Emerg. Infect. Dis. 2010, 16, 1237–1243. [Google Scholar] [CrossRef] [PubMed]
- Puechmaille, S.J.; Wibbelt, G.; Korn, V.; Fuller, H.; Forget, F.; Mühldorfer, K.; Kurth, A.; Bogdanowicz, W.; Borel, C.; Bosch, T.; et al. Pan-European Distribution of White-Nose Syndrome Fungus (Geomyces destructans) Not Associated with Mass Mortality. PLoS ONE 2011, 6, e19167. [Google Scholar] [CrossRef] [PubMed]
- Martínková, N.; Bačkor, P.; Bartonička, T.; Blažková, P.; Červený, J.; Falteisek, L.; Gaisler, J.; Hanzal, V.; Horáček, D.; Hubálek, Z.; et al. Increasing incidence of Geomyces destructans fungus in bats from the Czech Republic and Slovakia. PLoS ONE 2010, 5, e13853. [Google Scholar] [CrossRef] [PubMed]
- Zahradníková, A., Jr.; Kovacova, V.; Martínková, N.; Orlova, M.V.; Orlov, O.L.; Piacek, V.; Zukal, J.; Pikula, J. Historic and geographic surveillance of Pseudogymnoascus destructans possible from collections of bat parasites. Transbound. Emerg. Dis. 2018, 65, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Campana, M.G.; Kurata, N.P.; Foster, J.T.; Helgen, L.E.; Reeder, D.M.; Fleischer, R.C.; Helgen, K.M. White-Nose Syndrome Fungus in a 1918 Bat Specimen from France. Emerg. Infect. Dis. 2017, 23, 1611–1612. [Google Scholar] [CrossRef]
- Ballmann, A.E.; Torkelson, M.R.; Bohuski, E.A.; Russell, R.E.; Blehert, D.S. Dispersal Hazards of Pseudogymnoascus destructans by Bats and Human Activity at Hibernacula in Summer. J. Wildl. Dis. 2017, 53, 725–735. [Google Scholar] [CrossRef]
- Lorch, J.M.; Palmer, J.M.; Lindner, D.L.; Ballmann, A.E.; George, K.G.; Griffin, K.; Knowles, S.; Huckabee, J.R.; Haman, K.H.; Anderson, C.D.; et al. First Detection of Bat White-Nose Syndrome in Western North America. mSphere 2016, 1, e00148-16. [Google Scholar] [CrossRef]
- Khankhet, J.; Vanderwolf, K.J.; McAlpine, D.F.; McBurney, S.; Overy, D.P.; Slavic, D.; Xu, J. Clonal expansion of the Pseudo-gymnoascus destructans genotype in North America is accompanied by significant variation in phenotypic expression. PLoS ONE 2014, 9, e104684. [Google Scholar] [CrossRef]
- Ren, P.; Haman, K.H.; Last, L.A.; Rajkumar, S.S.; Keel, M.K.; Chaturvedi, V. Clonal spread of Geomyces destructans among bats, midwestern and southern United States. Emerg. Infect. Dis. 2012, 18, 883. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, S.S.; Li, X.; Rudd, R.J.; Okoniewski, J.C.; Xu, J.; Chaturvedi, S.; Chaturvedi, V. Clonal genotype of Geomyces de-structans among bats with white nose syndrome, New York, USA. Emerg. Infect. Dis. 2011, 17, 1273. [Google Scholar] [CrossRef] [PubMed]
- Palmer, J.M.; Kubatova, A.; Novakova, A.; Minnis, A.M.; Kolarik, M.; Lindner, D.L. Molecular characterization of a hetero-thallic mating system in Pseudogymnoascus destructans, the fungus causing white-nose syndrome of bats. G3 Genes Genomes Genet. 2014, 4, 1755–1763. [Google Scholar]
- Garzoli, L.; Riccucci, M.; Patriarca, E.; Debernardi, P.; Boggero, A.; Pecoraro, L.; Picco, A.M. First isolation of Pseudogymnoascus destructans, the fungal causative agent of white-nose disease, in bats from Italy. Mycopathologia 2019, 184, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Patriarca, E.; Debernardi, P.; Grazioli, F.; Garzoli, L.; Magagnoli, S.; Riccucci, M.; Boggero, A. La Grotta di Rio Martino: Importanza per i Chirotteri, fruizione antropica e gestione. In IV Convegno Italiano sui Chirotteri, Padova, 17–19 October 2019; Roscioni, F., Pidinchedda, E., Preatoni, D.G., Eds.; Gruppo Italiano Ricerca Chirotteri—Associazione Teriologica Italiana: Padova, Italy, 2019; p. 16. [Google Scholar]
- Speakman, J.R.; Racey, P.A. The influence of body condition on sexual development of male Brown long-eared bats (Plecotus auritus) in the wild. J. Zool. 1986, 210, 515–525. [Google Scholar] [CrossRef]
- Lanza, B. Mammalia V, Chiroptera, Fauna d’Italia; Edizioni Calderini de «Il Sole 24 ORE»: Milano, Italy, 2012; Volume 46. [Google Scholar]
- Dietz, C.; Kiefer, A. Bats of Britain and Europe; Bloomsbury Publishing: London, UK, 2016. [Google Scholar]
- Turner, G.G.; Meteyer, C.U.; Barton, H.; Gumbs, J.F.; Reeder, D.M.; Overton, B.; Bandouchova, H.; Bartonička, T.; Martínková, N.; Pikula, J.; et al. Nonlethal screening of bat-wing skin with the use of ultraviolet fluorescence to detect lesions indicative of white-nose syndrome. J. Wildl. Dis. 2014, 50, 566–573. [Google Scholar] [CrossRef]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Fungal Barcoding Consortium. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. 1990, 18, 315–322. [Google Scholar]
- Sigler, L.; Carmichael, J.W. Taxonomy of Malbranchea and some other Hyphomycetes with arthroconidia. Mycotaxon 1976, 4, 349–488. [Google Scholar]
- CABI. Pseudogymnoascus destructans (white-nose syndrome fungus). In Invasive Species Compendium; Blehert, D., Lankau, E., Eds.; CAB International: Wallingford, UK, 2020. [Google Scholar]
- Martínková, N.; Pikula, J.; Zukal, J.; Kovacova, V.; Bandouchova, H.; Bartonička, T.; Botvinkin, A.D.; Brichta, J.; Bundarova, H.; Kokurewicz, T.; et al. Hibernation temperature-dependent Pseudogymnoascus destructans infection intensity in Palearctic bats. Virulence 2018, 9, 1734–1750. [Google Scholar] [CrossRef]
- Dobony, C.A.; Hicks, A.C.; Langwig, K.E.; Von Linden, R.I.; Okoniewski, J.C.; Rainbolt, R.E. Little Brown Myotis Persist Despite Exposure to White-Nose Syndrome. J. Fish Wildl. Manag. 2011, 2, 190–195. [Google Scholar] [CrossRef]
- Langwig, K.E.; Frick, W.F.; Reynolds, R.; Parise, K.L.; Drees, K.P.; Hoyt, J.R.; Cheng, T.L.; Kunz, T.H.; Foster, J.T.; Kilpatrick, A.M. Host and pathogen ecology drive the seasonal dynamics of a fungal disease, white-nose syndrome. Proc. R. Soc. B Boil. Sci. 2015, 282, 20142335. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, G.M.; Wilcox, E.V.; Bernard, R.F.; Stiver, W.H. Detection of Pseudogymnoascus destructans on free-flying male bats captured during summer in the southeastern USA. J. Wildl. Dis. 2016, 52, 922–926. [Google Scholar] [CrossRef] [PubMed]
- Huebschman, J.J.; Hoerner, S.A.; White, J.P.; Kaarakka, H.M.; Parise, K.L.; Foster, J.T. Detection of Pseudogymnoascus destructans on Wisconsin Bats During Summer. J. Wildl. Dis. 2019, 55, 673–677. [Google Scholar] [CrossRef] [PubMed]
- Campbell, L.J.; Walsh, D.P.; Blehert, D.S.; Lorch, J.M. Long-term survival of Pseudogymnoascus destructans at elevated tem-peratures. J. Wildl. Dis. 2020, 56, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Flieger, M.; Bandouchova, H.; Cerny, J.; Chudíčková, M.; Kolarik, M.; Kovacova, V.; Martínková, N.; Novák, P.; Šebesta, O.; Stodůlková, E.; et al. Vitamin B2 as a virulence factor in Pseudogymnoascus destructans skin infection. Sci. Rep. 2016, 6, 33200. [Google Scholar] [CrossRef] [PubMed]
- Meteyer, C.U.; Barber, D.; Mandl, J.N. Pathology in euthermic bats with white nose syndrome suggests a natural manifestation of immune reconstitution inflammatory syndrome. Virulence 2012, 3, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Zukal, J.; Bandouchova, H.; Bartonička, T.; Berkova, H.; Brack, V.; Brichta, J.; Dolinay, M.; Jaron, K.S.; Kováčová, V.; Kovarik, M.; et al. White-Nose Syndrome Fungus: A Generalist Pathogen of Hibernating Bats. PLoS ONE 2014, 9, e97224. [Google Scholar] [CrossRef] [PubMed]
- Gianni, C.; Caretta, G.; Romano, C. Skin infection due to Geomyces pannorum var. pannorum. Fallbericht. Hautinfektion durch Geomyces pannorum var. pannorum. Mycoses 2003, 46, 430–432. [Google Scholar] [CrossRef]
- Sandoval-Denis, M.; Sutton, D.A.; Fothergill, A.W.; Cano-Lira, J.; Gené, J.; Decock, C.A.; De Hoog, G.S.; Guarro, J. Scopulariopsis, a Poorly Known Opportunistic Fungus: Spectrum of Species in Clinical Samples and In Vitro Responses to Antifungal Drugs. J. Clin. Microbiol. 2013, 51, 3937–3943. [Google Scholar] [CrossRef]
- Cunha, A.O.B.; Bezerra, J.D.P.; Oliveira, T.G.L.; Barbier, E.; Bernard, E.; Machado, A.R.; Souza-Motta, C.M. Living in the dark: Bat caves as hotspots of fungal diversity. PLoS ONE 2020, 15, e0243494. [Google Scholar] [CrossRef] [PubMed]
- Voyron, S.; Lazzari, A.; Riccucci, M.; Calvini, M.; Varese, G. First mycological investigation on Italian bats. Hystrix 2011, 22, 189–197. [Google Scholar]
- Zhelyazkova, V.; Hubancheva, A.; Radoslavov, G.; Toshkova, N.; Puechmaille, S.J. Did you wash your caving suit? Cavers’ role in the potential spread of Pseudogymnoascus destructans, the causative agent of White-Nose Disease. Int. J. Speleol. 2020, 49, 7. [Google Scholar] [CrossRef]
- Turbill, C.; Welbergen, J.A. Anticipating white-nose syndrome in the Southern Hemisphere: Widespread conditions favourable to Pseudogymnoascus destructans pose a serious risk to Australia’s bat fauna. Austral Ecol. 2020, 45, 89–96. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Species | Site and Role | N. Examined | Mean BM ± SD | Mean BCI ± SD | N. Swabs | N. Biopsies | ||
|---|---|---|---|---|---|---|---|---|
| (g) | (g/mm) | − | + | − | + | |||
| Rhinolophus ferrumequinum | 2H | 6 | 19.6 ± 1.96 | 33.8 ± 3.00 | 6 | 0 | 5 | 0 |
| 8H | 4 | 18.7 ± 2.08 | 32.7 ± 3.42 | 4 | 0 | 4 | 0 | |
| 9H | 2 | 15.2 ± 3.18 | 25.8 ± 5.56 | 2 | 0 | 2 | 0 | |
| Rhinolophus hipposideros | 9H | 4 | 4.8 ± 0.34 | 12.4 ± 0.70 | 4 | 0 | 4 | 0 |
| 10H | 5 | Nr | Nr | 5 | 0 | 0 | 0 | |
| Myotis blythii | 4H | 1 | Nr | Nr | 0 | 1 | 0 | 0 |
| 5H | 4 | 22.3 ± 0.77 | 38.6 ± 0.78 | 1 | 2 | 3 | 1 (1) | |
| Myotis capaccinii | 1HM | 13 | 9.2 ± 0.61 | 22.0 ± 1.27 | 8 * | 0 * | 9 | 1 |
| Myotis daubentonii | 5H | 10 | 6.7 ± 0.55 | 17.6 ± 1.31 | 6 | 1 | 3 | 2 (2) |
| Myotis emarginatus | 4H | 3 | 7.6 ± 1.04 | 19.0 ± 2.15 | 0 | 3 | 3 | 0 |
| 5H | 49 | 7.3 ± 0.97 | 18.1 ± 1.88 | 3 | 8 | 6 | 4 (1) | |
| Myotis myotis | 2H | 1 | 24.5 | 40.1 | 0 | 1 | 0 | 1 |
| 4H | 2 | 26.6 ± 1.06 | 42.3 ± 0.35 | 0 | 2 | 2 | 0 | |
| 5H | 1 | 23 | 38.1 | 1 | 0 | 1 | 0 | |
| 7HM | 2 | 23.9 ± 2.97 | 40.2 ± 4.04 | 2 | 0 | 1 | 1 | |
| 3M | 4 | 24.5 ± 0.88 | 39.5 ± 1.59 | 0 | 4 | 2 | 1 (1) | |
| 6M | 7 | 26.9 ± 1.95 | 42.4 ± 2.53 | 6 | 1 | 7 | 0 | |
| Barbastella barbastellus | 2H | 5 | 7.8 ± 0.47 | 19.9 ± 0.89 | 5 | 0 | 3 | 0 |
| 5H | 13 | 8.5 ± 0.74 | 21.8 ± 1.53 | 11 | 0 | 6 | 0 | |
| Hypsugo savii | 2H | 1 | Nr | Nr | 1 | 0 | 0 | 0 |
| Miniopterus schreibersii | 5H | 1 | 12.5 | 27.5 | 1 | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garzoli, L.; Bozzetta, E.; Varello, K.; Cappelleri, A.; Patriarca, E.; Debernardi, P.; Riccucci, M.; Boggero, A.; Girometta, C.; Picco, A.M. White-Nose Syndrome Confirmed in Italy: A Preliminary Assessment of Its Occurrence in Bat Species. J. Fungi 2021, 7, 192. https://doi.org/10.3390/jof7030192
Garzoli L, Bozzetta E, Varello K, Cappelleri A, Patriarca E, Debernardi P, Riccucci M, Boggero A, Girometta C, Picco AM. White-Nose Syndrome Confirmed in Italy: A Preliminary Assessment of Its Occurrence in Bat Species. Journal of Fungi. 2021; 7(3):192. https://doi.org/10.3390/jof7030192
Chicago/Turabian StyleGarzoli, Laura, Elena Bozzetta, Katia Varello, Andrea Cappelleri, Elena Patriarca, Paolo Debernardi, Marco Riccucci, Angela Boggero, Carolina Girometta, and Anna Maria Picco. 2021. "White-Nose Syndrome Confirmed in Italy: A Preliminary Assessment of Its Occurrence in Bat Species" Journal of Fungi 7, no. 3: 192. https://doi.org/10.3390/jof7030192
APA StyleGarzoli, L., Bozzetta, E., Varello, K., Cappelleri, A., Patriarca, E., Debernardi, P., Riccucci, M., Boggero, A., Girometta, C., & Picco, A. M. (2021). White-Nose Syndrome Confirmed in Italy: A Preliminary Assessment of Its Occurrence in Bat Species. Journal of Fungi, 7(3), 192. https://doi.org/10.3390/jof7030192