
The Emericellipsins A–E from an Alkalophilic Fungus Emericellopsis alkalina Show Potent Activity against Multidrug-Resistant Pathogenic Fungi

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Characterization of Alkalophilic Strains of Emericellopsis alkalina
2.2. Cultivation of the Fungi and Extraction of Emericellipsins A–E
2.3. Purification and Identification of Emericellipsins A–E
2.3.1. HPLC Analysis
2.3.2. MALDI TOF/TOF MS
2.3.3. LC/ESI-MS Analysis
2.3.4. UV Spectra
2.4. Biological Assays
2.4.1. Antifungal Activity
2.4.2. Cell Culture and Cytotoxicity Assays
2.4.3. Hemolytic Activity
2.4.4. Heterochromatin Condensation in Human Buccal Epithelium Cells
3. Results
3.1. Antimicrobial Activity of Crude Extracts of E. alkaline Strains
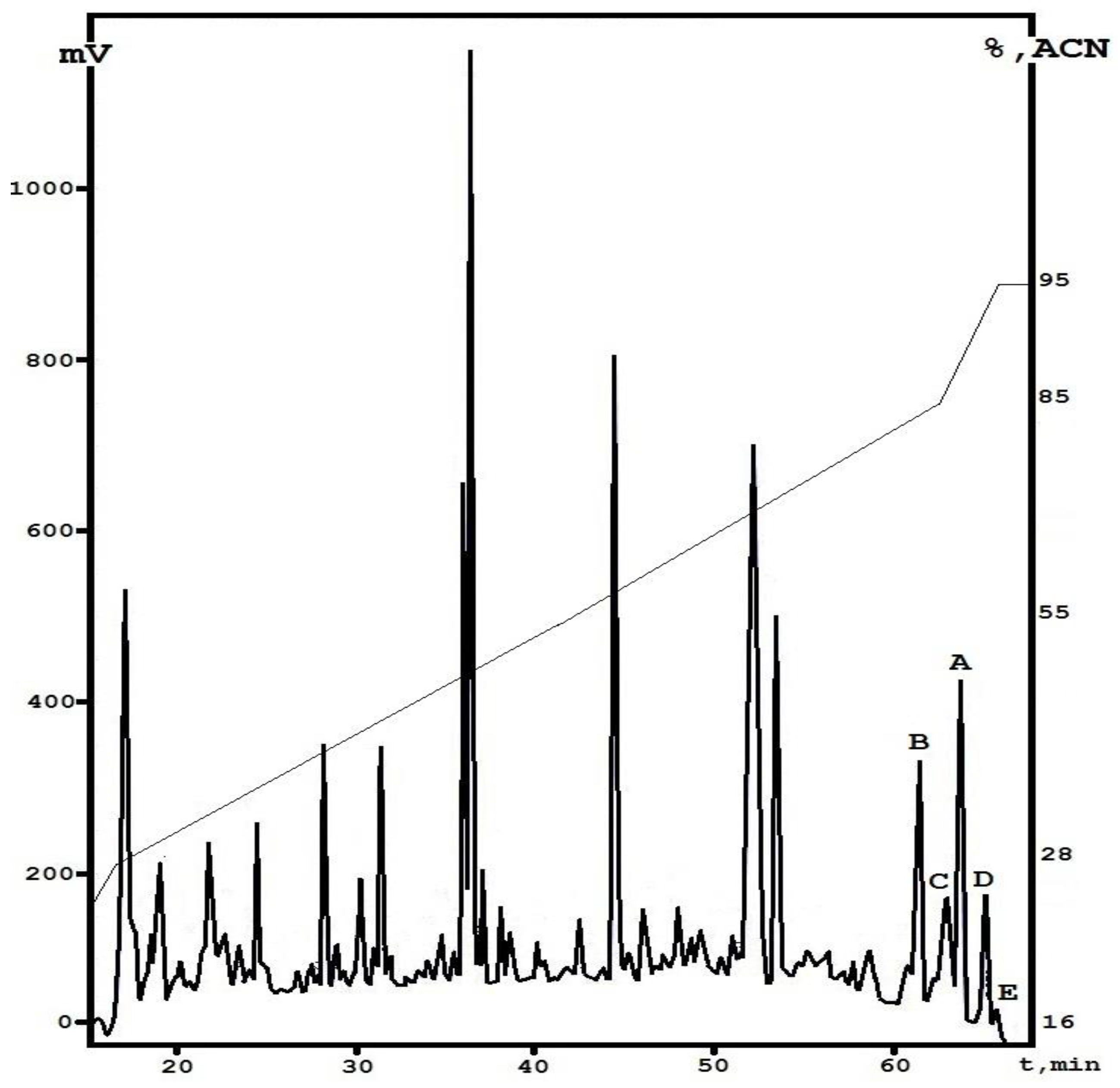
3.2. Isolation of the Individual Fractions from E. alkalina Concentrate
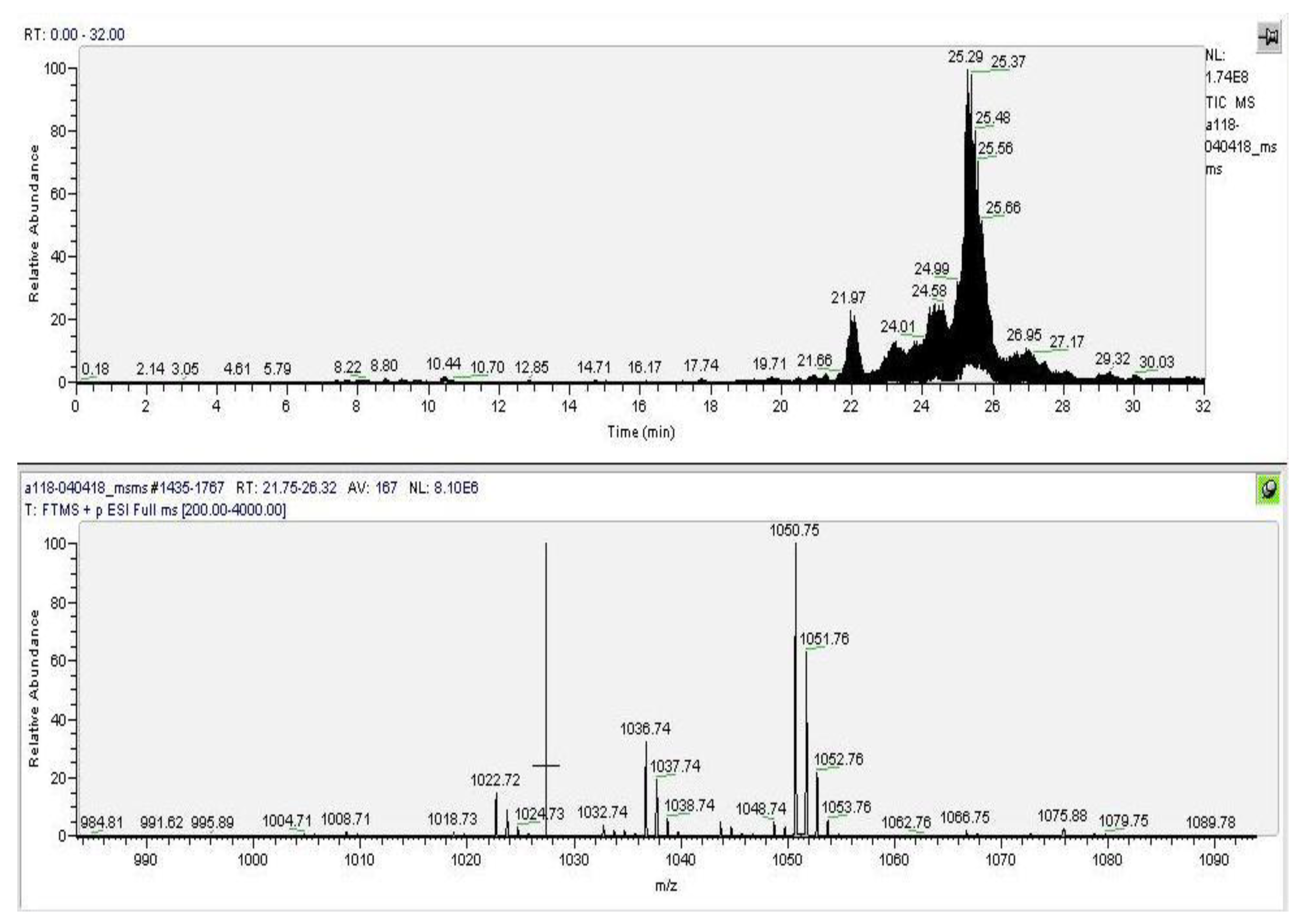
3.3. Initial Structural Analysis of the Individual Fractions
3.4. Identification of the Active Fractions and Structure Diversity of Peptaibols from E. alkalina
3.5. Antifungal Activity of Emericellipsins A–E by Disc-diffusion Assay
3.6. Activity of Emericellipsin A against an Extended Panel of Yeast Clinical Isolates
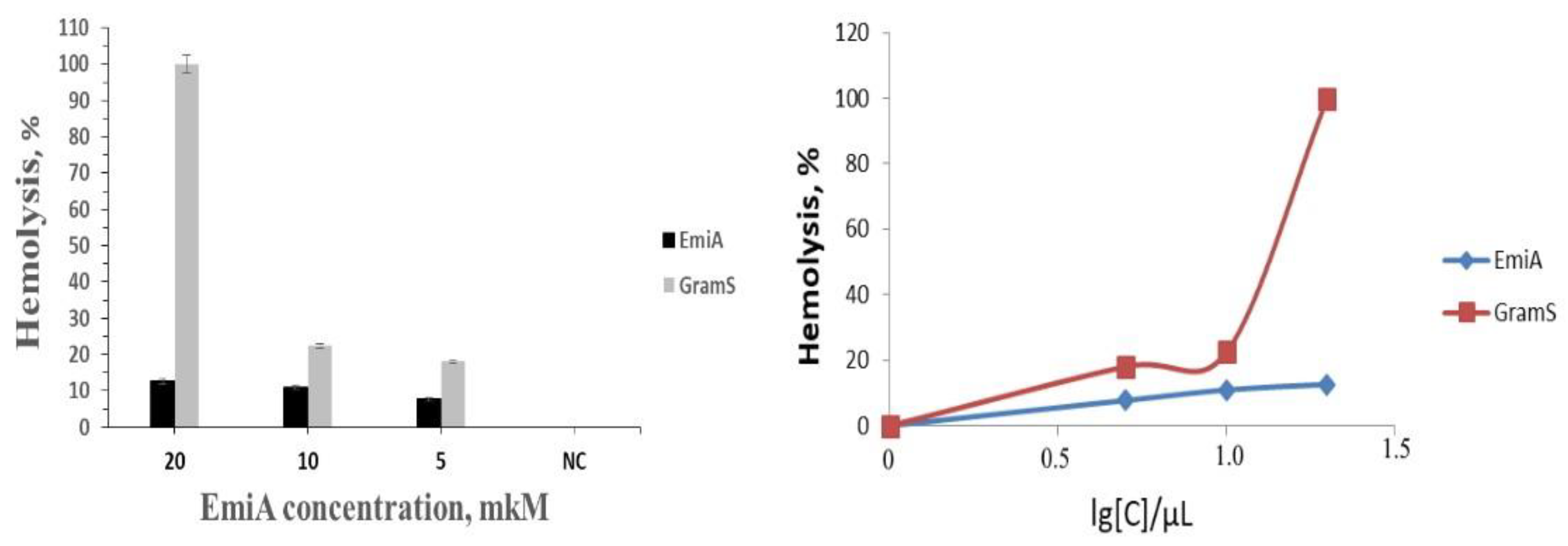
3.7. Cytotoxicity and Hemolytic Activity of Emericellipsin A
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Antinori, L.M.; Sollima, S.; Galli, M.; Corbellino, M. Candidemia and invasive candidiasis in adults: A narrative review. Eur. J. Intern. Med. 2016, 34, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Chastain, D.B.; Henao-Martınez, A.F.; Franco-Paredes, C. Opportunistic invasive mycoses in AIDS: Cryptococcosis, histoplasmosis, coccidiodomycosis, and talaromycosis. Curr. Infect. Dis. Rep. 2017, 19, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, V.; Bouchara, J.P.; Hagen, F.; Alastruey-Izquierdo, A.; Badali, H.; Bocca, A.L.; Cano-Lira, J.F.; Cao, C.; Chaturvedi, S.; Chotirmall, S.H.; et al. Eighty years of mycopathologia: A retrospective analysis of progress made in understanding human and animal fungal pathogens. Mycopathologia 2018, 183, 859–877. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, W.X.; Huang, K.W.; Cao, Z.X.; Hao, J.Y. Hospital acquired pneumonia occurring after acute stage of the serious SARS and its treating strategies. Chin. J. Nosocomiol. 2003, 11, 1081–1087. [Google Scholar]
- Zmeili, O.S.; Soubani, A.O. Pulmonary aspergillosis: A clinical update. Q. J. Med. 2007, 100, 317–334. [Google Scholar] [CrossRef]
- Butle, M.S.; Robertson, A.A.; Cooper, M.A. Natural product and natural product derived drugs in clinical trials. Nat. Prod. Rep. 2014, 31, 1612–1661. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Dou, X.; Song, J.; Liu, Y.; Zhu, X.; Xu, L.; Li, W.; Shan, A. Antimicrobial peptides: Promising alternatives in the post feeding antibiotic ERA. Med. Res. Rev. 2019, 39, 831–859. [Google Scholar] [CrossRef]
- Meyer, V.; Jung, S. Antifungal Peptides of the AFP Family Revisited: Are These Cannibal Toxins? Microorganisms 2018, 6, 50. [Google Scholar] [CrossRef]
- Meyer, V. A small protein that fights fungi: AFP as a new promising antifungal agent of biotechnological value. Appl. Microbiol. Biotechnol. 2008, 78, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Ishiyama, D.; Satou, T.; Senda, H.; Fujimaki, T.; Honda, R.; Kanazawa, S. Heptaibin, a novel antifungal peptaibol antibiotic from Emericellopsis sp. BAUA8289. J. Antibiot. 2000, 53, 728–732. [Google Scholar] [CrossRef]
- Zotti, M.D.; Biondi, B.; Peggion, C.P.; Hahm, Y.; Formaggio, K.S.; Toniolo, F.C. Synthesis, preferred conformation, protease stability, and membrane activity of heptaibin, a medium-length peptaibiotic. J. Pept. Sci. 2011, 17, 585–594. [Google Scholar] [CrossRef]
- Strobel, G.A.; Torczynski, R.; Bollon, A. Acremonium sp.: A leucinostatin A producing endophyte of European yew (Taxus baccata). Plant Sci. 1997, 128, 97–108. [Google Scholar] [CrossRef]
- Oh, S.U.; Yun, B.S.; Lee, S.J.; Kim, J.H.; Yoo, I.D. Atroviridins A-C and neoatroviridins A-D, novel peptaibol antibiotics produced by Trichoderma atroviride F80317. I. Taxonomy, fermentation, isolation and biological activities. J. Antibiot. 2002, 55, 557–564. [Google Scholar] [CrossRef]
- Shi, M.; Chen, L.; Wang, X.W.; Zhang, T.; Zhao, P.B.; Song, X.Y.; Sun, C.Y.; Chen, X.L.; Zhou, B.C.; Zhang, Y.Z. Antimicrobial peptaibols from Trichoderma pseudokoningii induce programmed cell death in plant fungal pathogens. Microbiology 2012, 158, 166–175. [Google Scholar] [CrossRef]
- Grum-Grzhimaylo, A.A.; Georgieva, M.L.; Debets, A.J.M.; Bilanenko, E.N. Are alkalitolerant fungi of the Emericellopsis lineage (Bionectriaceae) of marine origin? IMA Fungus 2013, 4, 211–226. [Google Scholar] [CrossRef] [PubMed]
- Zuccaro, A.; Summerbell, R.C.; Gams, W.; Schoers, H.J.; Mitchell, J.I. A new Acremonium species associated with Fucus spp., and its affinity with a phylogenetically distinct marine Emericellopsis clade. Stud. Mycol. 2004, 50, 283–297. [Google Scholar]
- Domsch, K.H.; Gams, W.; Anderson, T.H. Compendium of Soil Fungi, 2nd ed.; IHW: Eching, Germany, 2007; p. 384. ISBN 9783930167692. [Google Scholar]
- Gonçalves, M.F.M.; Vicente, T.F.L.; Esteves, A.C.; Alves, A. Novel halotolerant species of Emericellopsis and Parasarocladium associated with macroalgae in an estuarine environment. Mycologia 2020, 112, 154–171. [Google Scholar] [CrossRef]
- Baranova, A.A.; Georgieva, M.L.; Bilanenko, E.N.; Andreev, Y.A.; Rogozhin, E.A.; Sadykova, V.S. Antimicrobial potential of alkalophilic micromycetes Emericellopsis alkalina. Appl. Biochem. Microbiol. 2017, 53, 703–710. [Google Scholar] [CrossRef]
- Baranova, A.A.; Rogozhin, E.A.; Georgieva, M.L.; Bilanenko, E.N.; Kul’ko, A.B.; Yakushev, A.V.; Alferova, V.A.; Sadykova, V.S. Antimicrobial Peptides Produced by Alkaliphilic Fungi Emericellopsis alkalina: Biosynthesis and Biological Activity Against Pathogenic Multidrug-Resistant Fungi. Appl. Biochem. Microbiol. 2019, 55, 145–151. [Google Scholar] [CrossRef]
- Rogozhin, E.A.; Sadykova, V.S.; Baranova, A.A.; Vasilchenko, A.S.; Lushpa, V.A.; Mineev, K.S.; Georgieva, M.L.; Kul’ko, A.B.; Krasheninnikov, M.E.; Lyundup, A.V.; et al. A novel lipopeptaibol emericellipsin A with antimicrobial and antitumor activity produced by the extremophilic fungus Emericellopsis Alkalina. Molecules 2018, 23, 2785. [Google Scholar] [CrossRef]
- CLSI document M38-A2. Clinical and Laboratory Standards Institute (CLSI). Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi; Approved Standard, 2nd ed.; CLSI: Wayne, PA, USA, 2008; 35p, ISBN 1-56238-668-9. [Google Scholar]
- CLSI document M27-A3. Clinical and Laboratory Standards Institute (CLSI). Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts: Approved Standard, 3rd ed.; CLSI: Wayne, PA, USA, 2008; 25p, ISBN 1-56238-666-2. [Google Scholar]
- Tevyashova, A.N.; Korolev, A.M.; Trenin, A.S.; Dezhenkova, L.G.; Shtil, A.A.; Polshakov, V.I.; Savelyev, O.U.; Olsufyeva, E.N. New conjugates of polyene macrolide amphotericin B with benzoxaboroles: Synthesis and properties. J. Antibiot. 2016, 69, 549–560. [Google Scholar] [CrossRef] [PubMed]
- Shckorbatov, Y.G.; Grigoryeva, N.N.; Shakhbazov, V.G.; Grabina, V.A.; Bogoslavsky, A.M. Microwave irradiation influences on the state of human cell nuclei. Bioelectromagnetics 1998, 19, 414–419. [Google Scholar] [CrossRef]
- Setianingrum, F.; Rautemaa-Richardson, R.; Denning, D.W. Pulmonary cryptococcosis: A review of pathobiology and clinical aspects. Med. Mycol. 2019, 57, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Liang, G.; Liu, W. Fungal Co-infections Associated with Global COVID-19 Pandemic: A Clinical and Diagnostic Perspective from China. Mycopathologia 2020, 185, 599–606. [Google Scholar] [CrossRef] [PubMed]
- El Fane, M.; Badaoui, L.; Ouladlahsen, A.; Sodqi, M.; Marih, L.; Chakib, A.; Marhoum, K.E.F. Cryptococcosis during HIV infection. J. Mycol. Med. 2015, 25, 257–262. [Google Scholar] [CrossRef]
- Ayers, S.; Ehrmann, B.M.; Adcock, A.F.; Kroll, D.J.; Carcache de Blanco, E.J.; Shen, Q.; Swanson, S.M.; Falkinham, J.O.; Wani, M.C.; Mitchell, S.M.; et al. Peptaibols from two unidentified fungi of the order Hypocreales with cytotoxic, antibiotic, and anthelmintic activities. J. Pept. Sci. 2012, 18, 500–510. [Google Scholar] [CrossRef]
- Gangneux, J.P.; Bougnoux, M.E.; Dannaoui, E.; Cornet, M.; Zahar, J.R. Invasive fungal diseases during COVID-19: We should be prepared. J. Mycol. Med. 2020, unpublished. [Google Scholar] [CrossRef]
- Peeri, N.C.; Shrestha, N.; Rahman, M.S.; Zaki, R.; Tan, Z.; Bibi, S.; Mahdi Baghbanzadeh, M.; Aghamohammadi, N.; Zhang, W.; Haque, U. The SARS, MERS and novel coronavirus (COVID-19) epidemics, the newest and biggest global health threats: What lessons have we learned? Int. J. Epidemiol. 2020, 49, 717–726. [Google Scholar] [CrossRef]
- Yamazaki, H.; Rotinsulu, H.; Takahashi, O.; Kirikoshi, R.; Namikoshi, M. Induced production of a new dipeptide with a disulfide bridge by long-term fermentation of marine-derived Trichoderma cf. brevicompactum. Tetrahedron Lett. 2016, 57, 5764–5767. [Google Scholar] [CrossRef]
- Marik, T.; Urbán, P.; Tyagi, C.; Szekeres, A.; Leitgeb, B.; Vágvölgyi, M.; Manczinger, L.; Druzhinina, I.S.; Kredics, L. Diversity profile and dynamics of peptaibols produced by green mould Trichoderma species in interactions with their hosts Agaricus bisporus and Pleurotus ostreatus. Chem. Biodivers. 2017, 14, 1–21. [Google Scholar] [CrossRef]
- Katoch, M.; Singh, D.; Kapoor, K.K.; Vishwakarma, R.A. Trichoderma lixii (IIIM-B4), an endophyte of Bacopa monnieri L. producing peptaibols. BMC Microbiol. 2019, 19, 1–10. [Google Scholar] [CrossRef]
- Höfer, S.; Berg, A.; Brückner, H.; Mayerhöfer, T.G. Dispersion analysis of the orthorhombic peptide single crystal Z-(Aib)(3)-L-Ala-OtBu. Spectrochim Acta A Mol. Biomol. Spectrosc. 2019, 223, 1–8. [Google Scholar] [CrossRef]
- Brückner, H.; Fox, S.; Degenkolb, T. Sequences of Acretocins, Peptaibiotics Containing the Rare 1-Aminocyclopropanecarboxylic Acid, from Acremonium crotocinigenum CBS 217.70. Chem. Biodivers. 2019, 16, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wu, P.; Yao, L.; Xue, J.; Xu, L.; Li, H.; Deng, W.; Wei, X. Acremotins A-D, peptaibiotics produced by the soil-derived fungus Acremonium persicinum SC0105. J. Antibiot. 2018, 71, 927–938. [Google Scholar] [CrossRef]
- Quandt, C.A.; Bushley, K.E.; Spatafora, J.W. The genome of the truffle-parasite Tolypocladium ophioglossoides and the evolution of antifungal peptaibiotics. BMC Genom. 2015, 16, 1–14. [Google Scholar] [CrossRef]
- Marik, T.; Tyagi, C.; Racic, G.; Rakk, D.; Szekeres, A.; Vágvölgyi, C.; Kredics, L. New 19-Residue Peptaibols from Trichoderma Clade Viride. Microorganisms 2018, 6, 85. [Google Scholar] [CrossRef]
- Dalzini, A.; Bergamini, C.; Biondi, B.; Zotti, M.; Panighel, F.R.; Peggion, C.; Bortolus, M.; Maniero, A. The rational search for selective anticancer derivatives of the peptide Trichogin GA IV: A multitechnique biophysical approach. Sci. Rep. 2016, 6, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Saravanakumar, K.; Rajendren, N.; Kathiresan, K.; Myeong-Hyeon, W. Medicinal Drug-Related Bioactive Agents from Marine Fungi. In Encyclopedia of Marine Biotechnology, 1st ed.; Kim, S.-K., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2020; Volume 98, pp. 2173–2190. [Google Scholar]
- Niu, X.; Thaochan, N.; Hu, O. Diversity of Linear Non-Ribosomal Peptide in Biocontrol Fungi. J. Fungi 2020, 6, 61. [Google Scholar] [CrossRef]
- Ovchinnikova, T.V.; Levitskaya, N.G.; Voskresenskaya, O.G.; Yakimenko, Z.A.; Tagaev, A.A.; Ovchinnikova, A.Y.; Murashev, A.N.; Kamenskii, A.A. Neuroleptic properties of the ion-channel-forming peptaibol zervamicin: Locomotor activity and behavioral effects. Chem. Biodivers. 2007, 4, 1374–1387. [Google Scholar] [CrossRef]
- Rateb, M.E.; Ebel, R. Secondary metabolites of fungi from marine habitats. Nat. Prod. Rep. 2011, 28, 290–344. [Google Scholar] [CrossRef] [PubMed]
- De Ullivarri, M.F.; Arbulu, S.; Garcia-Gutierrez, E.; Cotter, P.D. Antifungal peptides as Therapeutic Agents. Cell. Infect. Microbiol. 2020, 10, 1–22. [Google Scholar] [CrossRef]
- Bunkóczi, G.; Schiell, M.; Vértesy, L.; Sheldrick, G.M. Crystal structures of cephaibols. J. Pept. Sci. 2003, 9, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Shenkarev, Z.O.; Paramonov, A.S.; Nadezhdin, K.D.; Bocharov, E.V.; Kudelina, I.A.; Skladnev, D.A.; Tagaev, A.A.; Yakimenko, Z.A.; Ovchinnikova, T.V.; Arseniev, A.S. Antiamoebin I in methanol solution: Rapid exchange between right-handed and left-handed 3(10)-helical conformations. Chem. Biodivers. 2007, 4, 1219–1242. [Google Scholar] [CrossRef]
- Shenkarev, Z.O.; Paramonov, A.S.; Lyukmanova, E.N.; Gizatullina, A.K.; Zhuravleva, A.V.; Tagaev, A.A.; Yakimenko, Z.A.; Telezhinskaya, I.N.; Kirpichnikov, M.P.; Ovchinnikova, T.V.; et al. Peptaibol antiamoebin I: Spatial structure, backbone dynamics, interaction with bicelles and lipid-protein nanodiscs, and pore formation in context of barrel-stave model. Chem. Biodivers. 2013, 10, 838–863. [Google Scholar] [CrossRef] [PubMed]
- Gessmann, R.; Axford, D.; Owen, R.L.; Brückner, H.; Petratos, K. Four complete turns of a curved 3₁₀-helix at atomic resolution: The crystal structure of the peptaibol trichovirin I-4A in a polar environment suggests a transition to α-helix for membrane function. Acta Cryst. D Biol.Cryst. 2012, 68, 109–116. [Google Scholar] [CrossRef]
- Bechinger, B.; Aisenbrey, C. The polymorphic nature of membrane-active peptides from biophysical and structural investigations. Curr. Protein Pept. Sci. 2012, 13, 602–610. [Google Scholar] [CrossRef]
- Peggion, C.; Biondi, B.; Battistella, C.; De Zotti, M.; Oancea, S.; Formaggio, F.; Toniolo, C. Spectroscopically labeled peptaibiotics. Synthesis and properties of selected trichogin GA IV analogs bearing a side-chain-monofluorinated aromatic amino acid for (19) F-NMR analysis. Chem. Biodivers. 2013, 10, 904–919. [Google Scholar] [CrossRef]
- Marik, T.; Tyagi, C.; Balázs, D.; Urbán, P.; Szepesi, Á.; Bakacsy, L.; Endre, G.; Rakk, D.; Szekeres, A.; Andersson, M.A.; et al. Structural Diversity and Bioactivities of Peptaibol Compounds From the Longibrachiatum Clade of the Filamentous Fungal Genus “Trichoderma”. Front. Microbiol. 2019, 10, 1434. [Google Scholar] [CrossRef] [PubMed]
- Jiao, W.H.; Khalil, Z.; Dewapriya, P.; Salim, A.A.; Lin, H.W.; Capon, R.J. Trichodermides A–E: New Peptaibols Isolated from the Australian Termite Nest-Derived Fungus Trichoderma virens CMB-TN16. J. Nat. Prod. 2018, 81, 976–984. [Google Scholar] [CrossRef]
- Singh, V.P.; Yedukondalu, N.; Sharma, V.; Kushwaha, M.; Sharma, R.; Chaubey, A.; Kumar, A.; Singh, D.; Vishwakarma, R.A. Lipovelutibols A-D: Cytotoxic Lipopeptaibols from the Himalayan Cold Habitat Fungus Trichoderma velutinum. J. Nat. Prod. 2018, 81, 219–226. [Google Scholar] [CrossRef]
- Zotti, M.D.; Sella, L.; Bolzonello, A.; Gabbatore, L.; Peggion, C.; Bortolotto, A.; Elmaghraby, I.; Tundo, S.; Favaron, F. Targeted Amino Acid Substitutions in a “Trichoderma” Peptaibol Confer Activity against Fungal Plant Pathogens and Protect Host Tissues from “Botrytis cinerea” Infection. Int. J. Mol. Sci. 2020, 21, 7521. [Google Scholar] [CrossRef] [PubMed]
- Gassiep, I.; Douglas, J.; Emeto, T.I.; Crawley, K.; Playford, E.G. Cryptococcal infections over a 15year period at a tertiary facility & impact of guideline management. Mycoses 2018, 61, 633–638. [Google Scholar] [CrossRef] [PubMed]
- Seyedjavadi, S.; Khani, S.; Zare-Zardini, H.; Halabian, R.; Goudarzi, M.; Khatami, S.; Fooladi, A.; Amani, J.; Razzaghi-Abyaneh, M. Isolation, functional characterization, and biological properties of MCh-AMP1, a novel antifungal peptide from Matricaria chamomilla L. Chem. Biol. Drug Des. 2019, 93, 949–959. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. of Strains, No. of VKM, VKPM, CBS Collections | Origin (Isolation Place, Soil’s Saltification Type) | Activity of Crude Peptaibols Extracts, Zone of Inhibition, mm | Total Content of EmiA | Presence of Homologues B–E | |
|---|---|---|---|---|---|
| A. niger ATCC 16404 | C. albicans ATCC 14053 | ||||
| E101 T =VKM F-4108; =CBS 127350 | Tanatar-II lake *, soda | 10 ± 0.2 | 0 | 0.61% | d. |
| A118 =VKPM F1428 | Zheltir’ lake *, soda-chloride | 26 ± 0.1 | 28 ± 0.6 | 1.2% | d. |
| A103 | Mirabilit lake *, soda-chloride-sulfate | 18 ± 0.1 | 15 ± 0.1 | 0.9% | d. |
| A114 =VKM FW-1473 | Solyonoe lake *, chloride | 25 ± 0.6 | 12 ± 0.6 | 0.05% | d. |
| A120 | Bezimyannoe lake *, soda | 12 ± 0.1 | 0 | 0.07% | n.d. |
| A121 VKM FW-1475 | Tanatar-II lake *, soda | 13 ± 0.4 | 9 ± 0.1 | 0.1% | n.d. |
| M20 =VKM FW-3040; =CBS 120044 | Zheltir’ lake *, soda-chloride | 26 ± 0.1 | 28 ± 0.6 | 1.07% | d. |
| 3KS17-13 | Malinovoye lake *, chloride | 7 ± 0.2 | 0 | 0.63% | n.d. |
| 5KS17-3 | Tanatar-II lake *, soda | 12 ± 0.2 | 0 | 0.42% | n.d. |
| 6KS17-1 | Tanatar-II lake *, soda | 6 ± 0.2 | 0 | 0.42% | n.d. |
| 7KS17-1 | Tanatar-II lake *, soda | 12 ± 0.2 | 0 | 0.42% | n.d. |
| 8KS17-1 | Tanatar-II lake *, soda | 11 ± 0.2 | 0 | n.d. | d. |
| 9KS17-3 | Tanatar-II lake *, soda | 14 ± 0.2 | 8 ± 0.2 | 0.54% | n.d. |
| 10KS17-1 | Tanatar-II lake *, soda | 15 ± 0.2 | 0 | n.d. | d. |
| 14KS17-1 | Tanatar I lake *, soda | 9 ± 0.2 | 12 ± 0.2 | 0.42% | d. |
| A126 =VKM FW-1472 | Transbaikal, Nuhe-Nur lake, soda | 12 ± 0.1 | 0 | 0.82% | n.d. |
| A128 | Transbaikal, Sulfatnoe lake, sulfate-soda | 14 ± 0.1 | 10 ± 0.1 | 0.12% | d. |
| Peptaibol | M+H, Da | Amino Acid Sequence |
|---|---|---|
| EmiA | 1050.69 | Methyldecanoyl-MePro-AHMOD-Ala-Aib-Ile-Iva-βAla-Alaol-Glyol |
| EmiB | 1036.77 | Methyldecanoyl-MePro-AHMOD-Ala-Aib-Ile-Aib-βAla-Alaol-Glyol |
| EmiC | 1066.76 | Methyldecanoyl-MePro-AHMOD-Ser-Aib-Ile-Iva-βAla-Alaol-Glyol |
| EmiD | 1052.76 | Methyldecanoyl-MePro-AHMOD-Ala-Iva-Ile-Iva-βAla-Alaol-Glyol |
| EmiE | 1079.75 | Methyldecanoyl-MePro-AHMOD-Ala-Aib-Ile-Iva-βAla-Aib-Glyol |
| Strain | Zone of Inhibition, mm | ||||||
|---|---|---|---|---|---|---|---|
| Compound | |||||||
| EmiA | EmiB | EmiC | EmiD | EmiE | Crude Peptaibols Complex | AmpB | |
| A. niger ATCC 16404 | 25 ± 0.21 | 10 ± 0.13 | 9 ± 0.31 | 0 | 0 | 20 ± 0.24 | 0 |
| A. niger 1133 m * | 18 ± 0.27 | 11 ± 0.170 | 0 | 0 | 0 | 12 ± 0.21 | 10 ± 0.14 |
| A. fumigatus VKM F-37 | 20 ± 0.3 | 0 | 0 | 0 | 0 | 12 ± 0.2 | 12 ± 0.23 |
| A. fumigatus 390 m * | 20 ± 0.40 | 0 | 0 | 0 | 0 | 10 ± 0.21 | 15 ± 0.51 |
| A. terreus 3 K | 12 ± 0.25 | 0 | 0 | 0 | 0 | 10 ± 0.31 | 0 |
| A. terreus 497 * | 14 ± 0.28 | 0 | 0 | 0 | 0 | 0 | 0 |
| Strain | Minimal Inhibitory Concentration (MIC) (µg/mL) | |||||
|---|---|---|---|---|---|---|
| Compound | ||||||
| EmiA | EmiB | EmiC | EmiD | EmiE | AmpB | |
| A. niger ATCC 16404 | 4 | 8 | 32 | ˃64 | ˃64 | 1 |
| A. niger 1133 m * | 4 | 8 | 32 | ˃64 | ˃64 | 1 |
| A. terreus 3 K | 0.5 | 4 | 8 | 16 | 32 | ˃64 |
| A. terreus 497 * | 1 | 4 | 8 | 16 | 64 | ˃64 |
| A. fumigatus VKM F-37 | 2 | 8 | 16 | 32 | ˃64 | 1 |
| A. fumigatus 390 m * | 2 | 8 | 16 | 32 | ˃64 | 1 |
| Strain | Minimal Inhibitory/Fungicidal Concentration (MIC/MFC) (µg/mL) | ||||||
|---|---|---|---|---|---|---|---|
| Compound | |||||||
| EmiA | AmpB | CAS | FZ | ||||
| MIC | MFC | MIC | MFC | MIC | MFC | ||
| Candida albicans 1402 | 2 | 4 | 0.5 | 0.5 | 0.06 | 0.5 | 64 |
| C. glabrata 1402 | 0.25 | 0.5 | 1 | 1 | 0.06 | 0.5 | 128 |
| C. krusei 1447 | 0.5 | 2 | 2 | 4 | 0.25 | 1 | R |
| C. tropicalis 156 | 1 | 2 | 0.5 | 1 | 0.06 | 0.5 | 64 |
| C. parapsilosis 571 | 1 | 1 | 1 | 1 | 1 | 1 | 128 |
| Cryptococcus neoformans 297 | 0.5 | 0.5 | 0.5 | 1 | >64 | n.d. | 16 |
| C. laurentii 325m | 0.5 | 0.5 | 0.25 | 0.5 | 16 | n.d. | 32 |
| Compound | IC50, µM | |||||
|---|---|---|---|---|---|---|
| HCT-116 | MCF-7 | HPF | K-562 | B16 | MDA-MB-231 | |
| EmiA | 2.30 ± 0.30 | 11.0 ± 1.20 | 11.5 ± 1.50 | 1.00 ± 0.14 | 16.0 ± 2.08 | 8.00 ± 1.04 |
| Doxorubicin | 0.20 ± 0.03 | 0.50 ± 0.06 | 0.20 ± 0.03 | 0.25 ± 0.03 | 0.60 ± 0.07 | 0.80 ± 0.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuvarina, A.E.; Gavryushina, I.A.; Kulko, A.B.; Ivanov, I.A.; Rogozhin, E.A.; Georgieva, M.L.; Sadykova, V.S. The Emericellipsins A–E from an Alkalophilic Fungus Emericellopsis alkalina Show Potent Activity against Multidrug-Resistant Pathogenic Fungi. J. Fungi 2021, 7, 153. https://doi.org/10.3390/jof7020153
Kuvarina AE, Gavryushina IA, Kulko AB, Ivanov IA, Rogozhin EA, Georgieva ML, Sadykova VS. The Emericellipsins A–E from an Alkalophilic Fungus Emericellopsis alkalina Show Potent Activity against Multidrug-Resistant Pathogenic Fungi. Journal of Fungi. 2021; 7(2):153. https://doi.org/10.3390/jof7020153
Chicago/Turabian StyleKuvarina, Anastasia E., Irina A. Gavryushina, Alexander B. Kulko, Igor A. Ivanov, Eugene A. Rogozhin, Marina L. Georgieva, and Vera S. Sadykova. 2021. "The Emericellipsins A–E from an Alkalophilic Fungus Emericellopsis alkalina Show Potent Activity against Multidrug-Resistant Pathogenic Fungi" Journal of Fungi 7, no. 2: 153. https://doi.org/10.3390/jof7020153
APA StyleKuvarina, A. E., Gavryushina, I. A., Kulko, A. B., Ivanov, I. A., Rogozhin, E. A., Georgieva, M. L., & Sadykova, V. S. (2021). The Emericellipsins A–E from an Alkalophilic Fungus Emericellopsis alkalina Show Potent Activity against Multidrug-Resistant Pathogenic Fungi. Journal of Fungi, 7(2), 153. https://doi.org/10.3390/jof7020153