
Evaluation of Native Entomopathogenic Fungi for the Control of Fall Armyworm (Spodoptera frugiperda) in Thailand: A Sustainable Way for Eco-Friendly Agriculture

 ,
,  , ,
, , 

Abstract
1. Introduction
2. Materials and Methods
2.1. Collection and Isolation of Entomopathogenic Fungi
2.2. Identification of Entomopathogenic Fungi
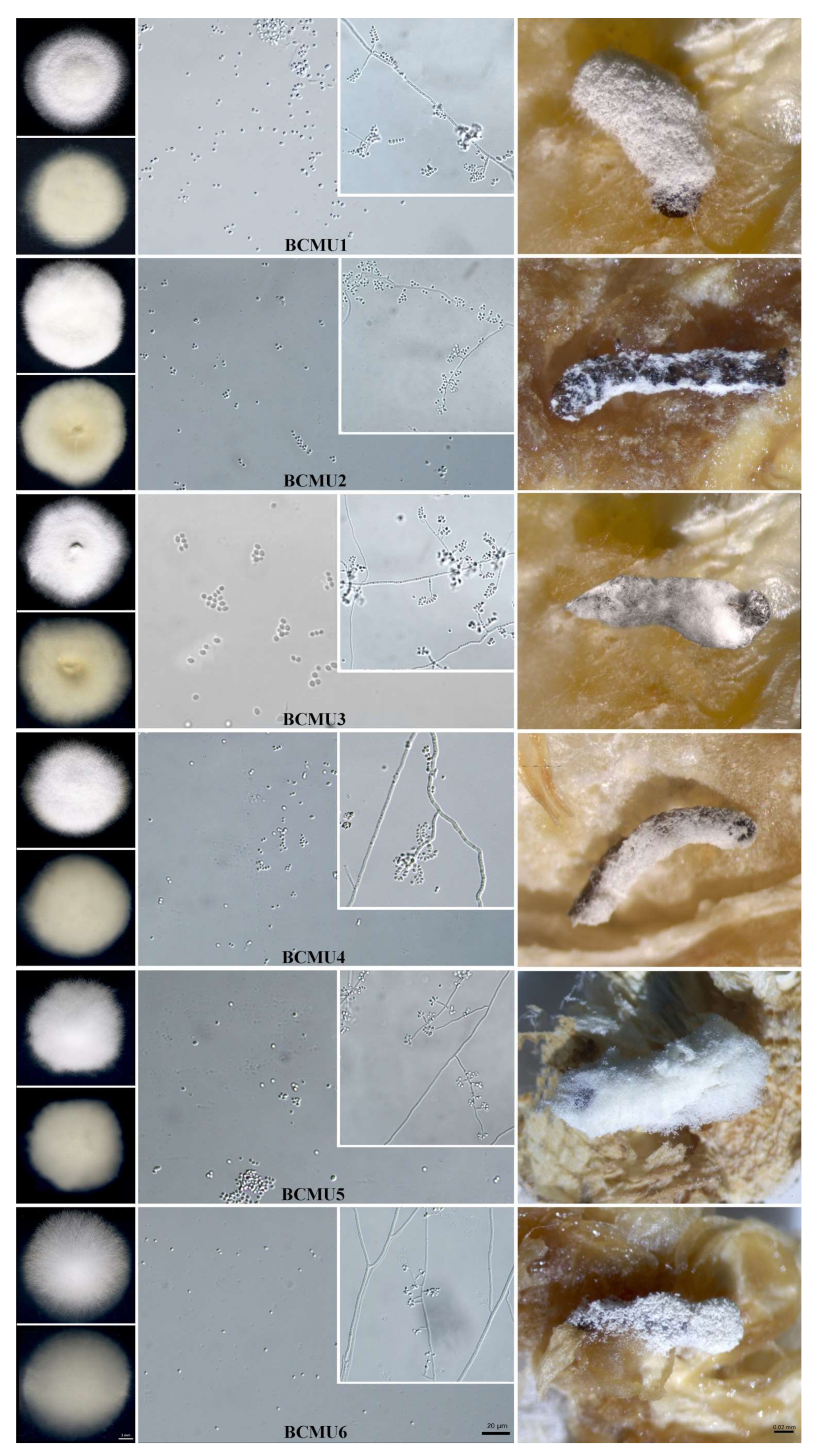
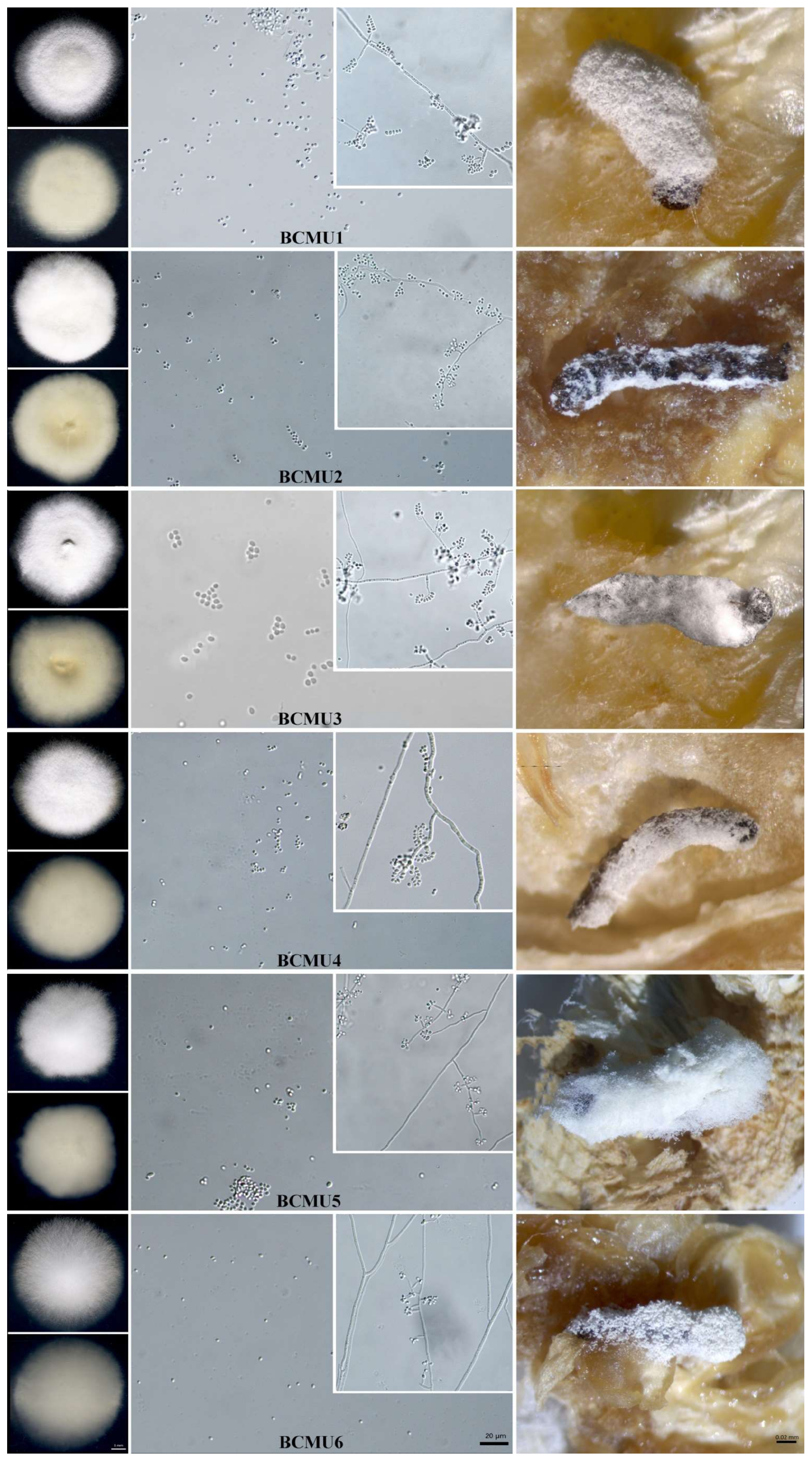
2.2.1. Morphological Study
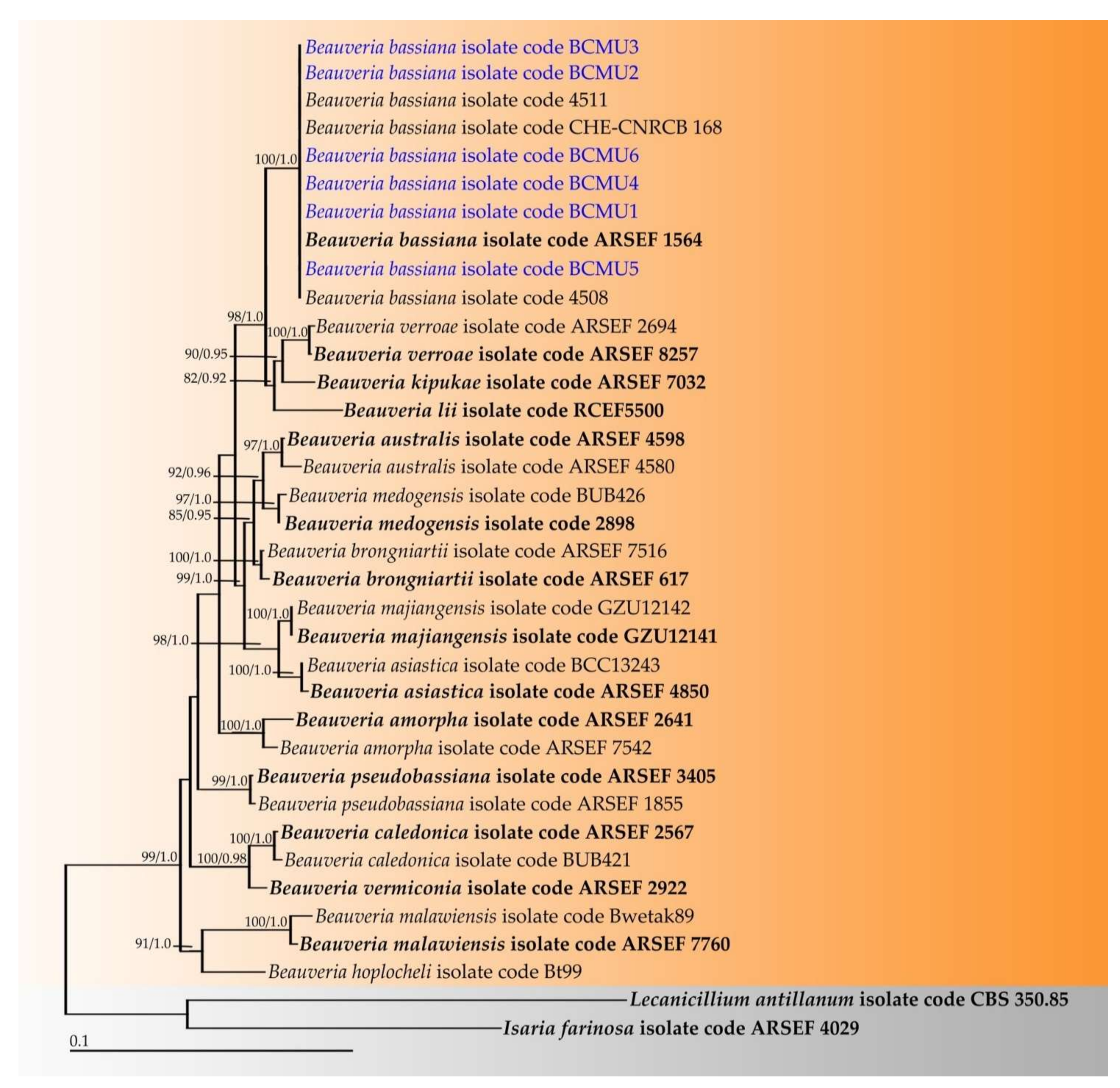
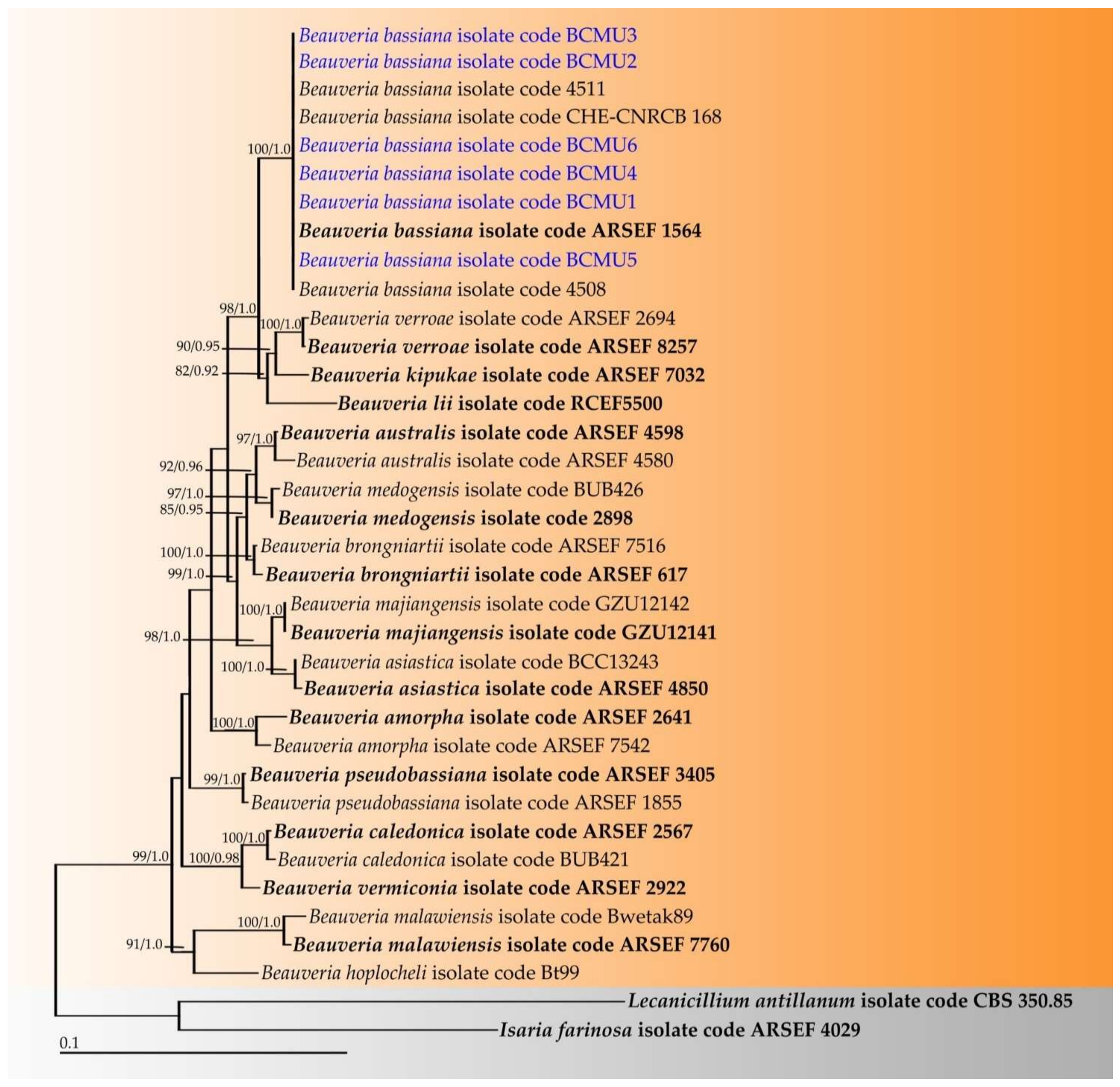
2.2.2. Molecular Study
2.3. Efficacy Test
2.3.1. Insect Source and Rearing
2.3.2. Fungal Inoculum Preparation
2.3.3. Insect Bioassay
2.4. Molecular Characterization of GAS1 Gene
3. Results
3.1. Collection and Morphological Characterization of Entomopathogenic Fungi
3.2. Phylogenetic Results of Obtained Entomopathogenic Fungi
3.3. Efficacy Test
3.4. Molecular Characterization of GAS1 Gene
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rosegrant, M.R.; Ringler, C.; Sulser, T.B.; Ewing, M.; Palazzo, A.; Zhu, T.; Nelson, G.C.; Koo, J.; Robertson, R.; Msangi, S.; et al. Agriculture and Food Security under Global Change: Prospects for 2025/2050; International Food Policy Research Institute (IFPR): Washington, DC, USA, 2009; pp. 145–178. [Google Scholar]
- Womack, E.D.; Williams, W.P.; Smith, J.S.; Warburton, M.L.; Bhattramakki, D.; Hesler, L. Mapping Quantitative Trait Loci for Resistance to Fall Armyworm (Lepidoptera: Noctuidae) Leaf-Feeding Damage in Maize Inbred Mp705. J. Econ. Entomol. 2020, 113, 956–963. [Google Scholar] [CrossRef] [PubMed]
- Ekasingh, B.; Gypmantasiri, P.; Thong-Ngam, K.; Grudloyma, P. Maize in Thailand: Production Systems, Constraints, and Research Priorities; CIMMYT: Bangkok, Thailand, 2014. [Google Scholar]
- Hernandez-Mendoza, J.L.; López-Barbosa, E.C.; Garza-González, E.; Mayek-Perez, N. Spatial distribution of Spodoptera frugiperda (Lepidoptera: Noctuidae) in maize landraces grown in Colima, Mexico. J. Trop. Insect Sci. 2008, 28, 126–129. [Google Scholar] [CrossRef]
- Sartiami, D.; Harahap, I.S.; Kusumah, Y.M.; Anwar, R. First record of fall armyworm (Spodoptera frugiperda) in Indonesia and its occurrence in three provinces. IOP Conf. Ser. Earth Environ. Sci. 2020, 468, 012021. [Google Scholar] [CrossRef]
- Abel, C.A.; Scott, M.P. Evaluation of 21 Thailand Maize Germplasms for Resistance to Leaf Feeding Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Kansas Entomol. Soc. 2020, 93, 97–102. [Google Scholar] [CrossRef]
- Early, R.; González-Moreno, P.; Murphy, S.T.; Day, R. Forecasting the global extent of invasion of the cereal pest Spodoptera frugiperda, the fall armyworm. NeoBiota 2018, 40, 25–50. [Google Scholar] [CrossRef]
- Aktar, W.; Sengupta, D.; Chowdhury, A. Impact of pesticides use in agriculture: Their benefits and hazards. Interdiscip. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef]
- Norjmaa, U.; Nasamdulam, D.; Enkhjargal, B.; Banzragch, D. Morphological and molecular identification of Beauveria bassiana from agricultural soils. Mong. J. Agric. Sci. 2019, 27, 20–24. [Google Scholar] [CrossRef]
- Sandhu, S.S.; Sharma, A.K.; Beniwal, V.; Goel, G.; Batra, P.; Kumar, A.; Jaglan, S.; Sharma, A.K.; Malhotra, S. Myco-Biocontrol of Insect Pests: Factors Involved, Mechanism, and Regulation. J. Pathog. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Rajula, J.; Rahman, A.; Krutmuang, P. Entomopathogenic fungi in Southeast Asia and Africa and their possible adoption in biological control. Biol. Control 2020, 151, 104399. [Google Scholar] [CrossRef]
- Draganova, S.; Donkova, R.; Georgieva, D. Impact of Strains of Entomopathogenic Fungi on Some Main Groups of Soil Microorganisms. J. Plant Prot. Res. 2008, 48, 169–179. [Google Scholar] [CrossRef]
- Kulu, I.P.; Abadi, A.L.; Afandhi, A. Morphological and molecular identification of Beauveria bassiana as entomopathogen agent from central kalimantan Peatland, Indonesia. Int. J. ChemTech Res. 2015, 8, 2079–2084. [Google Scholar]
- Ruiz-Nájera, R.E.; Ruiz-Estudillo, R.A.; Sánchez-Yáñez, J.M.; Molina-Ochoa, J.; Skoda, S.R.; Coutiño-Ruiz, R.; Pinto-Ruiz, R.; Guevara-Hernández, F.; Foster, J.E. Occurrence of Entomopathogenic Fungi and Parasitic Nematodes on Spodoptera frugiperda (Lepidoptera: Noctuidae) Larvae Collected in Central Chiapas, México. Fla. Entomol. 2013, 96, 498–503. [Google Scholar] [CrossRef]
- Acosta, R.I.T.; Humber, R.A.; Sánchez-Peña, S.R. Zoophthora radicans (Entomophthorales), a fungal pathogen of Bagrada hilaris and Bactericera cockerelli (Hemiptera: Pentatomidae and Triozidae): Prevalence, pathogenicity, and interplay of environmental influence, morphology, and sequence data on fungal identification. J. Invertebr. Pathol. 2016, 139, 82–91. [Google Scholar]
- Thaochan, N.; Sausa-Ard, W. Occurrence and effectiveness of indigenous Metarhizium anisopliae against adults Zeugodacus cucurbitae (Coquillett)(Diptera: Tephritidae) in Southern Thailand. Songklanakarin J. Sci. Technol. 2017, 39, 325–334. [Google Scholar] [CrossRef]
- Wanjiru, J.; Sunday, K. Ovicidal effects of entomopathogenic fungal isolates on the invasive Fall armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Appl. Entomol. 2019, 143, 626–634. [Google Scholar] [CrossRef]
- Keppanan, R.; Sivaperumal, S.; Hussain, M.; Bamisile, B.S.; Aguila, L.C.R.; Qasim, M.; Mekchaya, S.; Wangade, L.; Krutmuang, P. Molecular characterization of pathogenesis involving the GAS 1 gene from Entomopathogenic fungus Lecanicillium lecanii and its virulence against the insect host Diaphorina citri. Pestic. Biochem. Physiol. 2019, 157, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Xia, Y.; Keyhani, N.O. Contribution of the gas1 gene of the entomopathogenic fungus Beauveria bassiana, encoding a putative glycosylphosphatidylinositol-anchored β-1,3-glucanosyltransferase, to conidial thermotolerance and virulence. Appl. Environ. Microbiol. 2011, 77, 2676–2684. [Google Scholar] [CrossRef]
- Cao, Y.; Zhu, X.; Jiao, R.; Xia, Y. The Magas1 gene is involved in pathogenesis by affecting penetration in Metarhizium acridum. J. Microbiol. Biotechnol. 2012, 22, 889–893. [Google Scholar] [CrossRef]
- Meyling, N.V. Methods for Isolation of Entomopathogenic Fungi from the Soil Environment. 1–18 January 2007. Available online: http://orgprints.org/11200 (accessed on 4 November 2021).
- Wang, Y.; Tang, D.X.; Duan, D.E.; Wang, Y.B.; Yu, H. Morphology, molecular characterization, and virulence of Beauveria pseudobassiana isolated from different hosts. J. Invertebr. Pathol. 2020, 172, 107333. [Google Scholar] [CrossRef]
- Affandi, A.; Chailani, S.R.; Mimbar, S.M.; Wiroatmodjo, B. Isolation and Phenotypic Characterization of Morphology in Fungus Beauveria bassiana (Balsamo) Vuillemin Colony Naturally From Leaf Surface, Soil, and Insect as Host in Tomato Plantation. AGRIVITA J. Agric. Sci. 2012, 34, 303–310. [Google Scholar] [CrossRef][Green Version]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Rehner, S.A. Primers for Elongation Factor 1-Alpha (EF1-Alpha). 2001. Available online: http://ocid.NACSE.ORG/research/deephyphae/EF1primer.pdf. (accessed on 11 December 2021).
- Castlebury, L.A.; Rossman, A.Y.; Sung, G.H.; Hyten, A.S.; Spatafora, J.W. Multigene phylogeny reveals new lineage for Stachybotrys chartarum, the indoor air fungus. Mycol. Res. 2004, 108, 864–872. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Muscle: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 1–19. [Google Scholar] [CrossRef]
- Hall, T. BioEdit. Available online: http://www.mbio.ncsu.edu/bioedit/bioedit.html (accessed on 24 August 2021).
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Gateway Computing Environments Workshop, GCE 2010. Available online: http://www.proceedings.com/10226.html (accessed on 4 November 2021).
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and high-performance computing Europe PMC Funders Group. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree Tree Figure Drawing Tool Version 131. Institute of Evolutionary 623 Biology, University of Edinburgh. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 4 November 2021).
- Rehner, S.A.; Minnis, A.M.; Sung, G.H.; Luangsa-ard, J.J.; Devotto, L.; Humber, R.A. Phylogeny and systematics of the anamorphic, entomopathogenic genus Beauveria. Mycologia 2011, 103, 1055–1073. [Google Scholar] [CrossRef]
- Serna-Domínguez, M.G.; Andrade-Michel, G.Y.; Arredondo-Bernal, H.C.; Gallou, A. Two efficient methods for isolation of high-quality genomic DNA from entomopathogenic fungi. J. Microbiol. Methods 2018, 148, 55–63. [Google Scholar] [CrossRef]
- Imoulan, A.; Wu, H.J.; Lu, W.L.; Li, Y.; Li, B.B.; Yang, R.H.; Wang, W.J.; Wang, X.L.; Kirk, P.M.; Yao, Y.J. Beauveriamedogensis sp. nov., a new fungus of the entomopathogenic genus from China. J. Invertebr. Pathol. 2016, 139, 74–81. [Google Scholar] [CrossRef]
- Zhang, S.L.; He, L.M.; Chen, X.; Huang, B. Beauveria lii sp. nov. isolated from Henosepilachna vigintioctopunctata. Mycotaxon 2013, 121, 199–206. [Google Scholar] [CrossRef]
- Rehner, J.F.; de Muro, S.A.; Bischoff, M.A. Description and phylogenetic placement of Beauveria malawiensis sp. nov. (Clavicipitaceae, Hypocreales). Mycotaxon 2006, 98, 137–145. [Google Scholar]
- Glare, T.; Campbell, M.; Biggs, P.; Winter, D.; Durrant, A.; McKinnon, A.; Cox, M. Mitochondrial evolution in the entomopathogenic fungal genus Beauveria. Arch. Insect Biochem. Physiol. 2020, 105, e21754. [Google Scholar] [CrossRef]
- Khonsanit, A.; Luangsa-ard, J.J.; Thanakitpipattana, D.; Noisripoom, W.; Chaitika, T.; Kobmoo, N. Cryptic diversity of the genus Beauveria with a new species from Thailand. Mycol. Prog. 2020, 19, 291–315. [Google Scholar] [CrossRef]
- Chen, W.H.; Liu, M.; Huang, Z.X.; Yang, G.M.; Han, Y.F.; Liang, J.D.; Liang, Z.Q. Beauveria majiangensis, a new entomopathogenic fungus from Guizhou, China. Phytotaxa 2018, 333, 243–250. [Google Scholar] [CrossRef]
- Robène-Soustrade, I.; Jouen, E.; Pastou, D.; Payet-Hoarau, M.; Goble, T.; Linderme, D.; Lefeuvre, P.; Calmès, C.; Reynaud, B.; Nibouche, S.; et al. Description and phylogenetic placement of Beauveria hoplocheli sp. nov. used in the biological control of the sugarcane white grub, Hoplochelus marginalis, on Reunion Island. Mycologia 2015, 107, 1221–1232. [Google Scholar] [CrossRef] [PubMed]
- Rehner, S.A.; Buckley, E.A. Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- Spatafora, J.W.; Sung, G.H.; Sung, J.M.; Hywel-Jones, N.L.; White, J.F. Phylogenetic evidence for an animal pathogen origin of ergot and the grass endophytes. Mol. Ecol. 2007, 16, 1701–1711. [Google Scholar] [CrossRef] [PubMed]
- Imoulan, A.; Elmeziane, A. Pathogenicity of Beauveria bassiana isolated from Moroccan Argan forests soil against larvae of Ceratitis capitata (Diptera: Tephritidae) in laboratory conditions. World J. Microbiol. Biotechnol. 2014, 30, 959–965. [Google Scholar] [CrossRef]
- Sade, C.; Da, B.; Roberto, J.; Parra, P. New method for rearing Spodoptera frugiperda in laboratory shows that larval cannibalism is not obligatory. Rev. Bras. Entomol. 2013, 57, 347–349. [Google Scholar]
- Bolzan, A.; Padovez, F.E.; Nascimento, A.R.; Kaiser, I.S.; Lira, E.C.; Amaral, F.S.; Kanno, R.H.; Malaquias, J.B.; Omoto, C. Selection and characterization of the inheritance of resistance of Spodoptera frugiperda (Lepidoptera: Noctuidae) to chlorantraniliprole and cross-resistance to other diamide insecticides. Pest Manag. Sci. 2019, 75, 2682–2689. [Google Scholar] [CrossRef]
- Safavi, S. Isolation, identification and pathogenicity assessment of a new isolate of entomopathogenic fungus, Beauveria bassiana in Iran. J. Plant Prot. Res. 2010, 50, 158–163. [Google Scholar] [CrossRef]
- Imoulan, A.; Hussain, M.; Kirk, P.M.; El Meziane, A.; Yao, Y.J. Entomopathogenic fungus Beauveria: Host specificity, ecology and significance of morpho-molecular characterization in accurate taxonomic classification. J. Asia Pac. Entomol. 2017, 20, 1204–1212. [Google Scholar] [CrossRef]
- Hegedus, D.D.; Khachatourians, G.G. Construction of cloned DNA probes for the specific detection of the entomopathogenic fungus Beauveria bassiana in grasshoppers. J. Invertebr. Pathol. 1993, 62, 233–240. [Google Scholar] [CrossRef]
- Hassan, F.R.; Abdullah, S.K.; Assaf, L.H. Pathogenicity of the entomopathogenic fungus, Beauveria bassiana (Bals.) Vuill. endophytic and a soil isolate against the squash beetle, Epilachna chrysomelina (F.) (Coleoptera: Coccinellidae). Egypt. J. Biol. Pest Control 2019, 29, 1–7. [Google Scholar] [CrossRef]
- Talaei-Hassanloui, R.; Kharazi-Pakdel, A.; Goettel, M.; Mozaffari, J. Variation in virulence of Beauveria bassiana isolates and its relatedness to some morphological characteristics. Biocontrol Sci. Technol. 2006, 16, 525–534. [Google Scholar] [CrossRef]
- Oreste, M.; Bubici, G.; Poliseno, M.; Triggiani, O.; Tarasco, E. Pathogenicity of Beauveria bassiana (Bals.-Criv.) Vuill. and Metarhizium anisopliae (Metschn.) sorokin against Galleria mellonella L. and Tenebrio molitor L. in laboratory assays. Redia 2012, 95, 43–48. [Google Scholar]
- Yasin, M.; Wakil, W.; Ghazanfar, M.U.; Qayyum, M.A.; Tahir, M.; Bedford, G.O. Virulence of entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae against red palm weevil, Rhynchophorus ferrugineus (Olivier). Entomol. Res. 2019, 49, 3–12. [Google Scholar] [CrossRef]
- Ekesi, S.; Akpa, A.D.; Onu, I.; Ogunlana, M.O. Entomopathogenicity of Beauveria bassiana and Metarhizium anisopliae to the cowpea aphid, Aphis craccivora koch (Homoptera: Aphididae). Arch. Phytopathol. Plant Prot. 2000, 33, 171–180. [Google Scholar] [CrossRef]
- Macuphe, N.; Oguntibeju, O.O.; Nchu, F. Evaluating the endophytic activities of Beauveria bassiana on the physiology, growth, and antioxidant activities of extracts of lettuce (Lactuca sativa L.). Plants 2021, 10, 1178. [Google Scholar] [CrossRef]
- Barra-Bucarei, L.; González, M.G.; Iglesias, A.F.; Aguayo, G.S.; Peñalosa, M.G.; Vera, P.V. Beauveria bassiana multifunction as an endophyte: Growth promotion and biologic control of Trialeurodes vaporariorum, (Westwood) (Hemiptera: Aleyrodidae) in tomato. Insects 2020, 11, 591. [Google Scholar] [CrossRef]
- Russo, M.L.; Scorsetti, A.C.; Vianna, M.F.; Allegrucci, N.; Ferreri, N.A.; Cabello, M.N.; Pelizza, S.A. Effects of endophytic Beauveria bassiana (Ascomycota: Hypocreales) on biological, reproductive parameters and food preference of the soybean pest Helicoverpa gelotopoeon. J. King Saud Univ.-Sci. 2019, 31, 1077–1082. [Google Scholar] [CrossRef]
- Xiao, G.; Ying, S.H.; Zheng, P.; Wang, Z.L.; Zhang, S.; Xie, X.Q.; Shang, Y.; Leger, R.J.; Zhao, G.P.; Feng, M.G. Genomic perspectives on the evolution of fungal entomopathogenicity in Beauveria bassiana. Sci. Rep. 2012, 2, 483. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Chen, H.; Wei, G.; Wang, G.; Li, F.; Wang, S. In vivo gene expression profiling of the entomopathogenic fungus Beauveria bassiana elucidates its infection stratagems in Anopheles mosquito. Sci. China Life Sci. 2017, 60, 839–851. [Google Scholar] [CrossRef] [PubMed]
- Ávila-Hernández, J.G.; Carrillo-Inungaray, M.L.; Cruz-Quiroz, R.D.; Wong-Paz, J.E.; Muñiz-Márquez, D.B.; Parra, R.; Aguilar, C.N.; Aguilar-Zárate, P. Beauveria bassiana secondary metabolites: A review inside their production systems, biosynthesis, and bioactivities. Mex. J. Biotechnol. 2020, 5, 1–33. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Name | Primer Sequence | Reference |
|---|---|---|---|
| ITS | ITS1F | CTTGGTCATTTAGAGGAAGTAA | [24] |
| ITS4 | TCCTCCGCTTATTGATATGC | [25] | |
| TEF-1 | 983F | GCTCCYGGHCAYCGTGAYTTYAT | [26] |
| 2218R | ATGACACCRACRGCRACRGTYTG | [26] | |
| RPB1 | RPB1-Af | GAR TGYCCDGGDCAYTTYGG | [27] |
| RPB1-Cr | CCNGCDATNTCRTTRTCCATRTA | [27] | |
| RPB2 | fRPB2-5f2 | GAYGAYMGWGATCAYTTYGG | [28] |
| fRPB2-7cR | CCCATRGCTTGYTTRCCCAT | [28] |
| Strain and Voucher No. | Taxon | Country | Host/substratum | GenBank Accession Number | Reference | |||
|---|---|---|---|---|---|---|---|---|
| ITS | TEF-1 | RPB1 | RPB2 | |||||
| BCMU1 | Beauveria bassiana | Thailand | Bactocera dorsalis | OL375165 | OL410297 | OL410303 | OL410309 | This study |
| BCMU2 | Beauveria bassiana | Thailand | Coffeeberry borer | OL375167 | OL410298 | OL410304 | OL410310 | This study |
| BCMU3 | Beauveria bassiana | Thailand | Ant | OL375168 | OL410299 | OL410305 | OL410311 | This study |
| BCMU4 | Beauveria bassiana | Thailand | Coffeeberry borer | OL375169 | OL410300 | OL410306 | OL410312 | This study |
| BCMU5 | Beauveria bassiana | Thailand | Coffee stem borer | OL375170 | OL410301 | OL410307 | OL410313 | This study |
| BCMU6 | Beauveria bassiana | Thailand | Bactocera dorsalis | OL375173 | OL410302 | OL410308 | OL410314 | This study |
| ARSEF 1564 T | Beauveria bassiana | Italy | Hyphantria cunea | GU734762 | EF222318 | HQ880833 | HQ880905 | [35] |
| CHE-CNRCB 168 | Beauveria bassiana | Mexico | Diaphorina citri | KU725691 | KU725693 | KU725699 | KU725703 | [36] |
| Isolate 4511 | Beauveria bassiana | China | Soil | KX901310 | KX901322 | KX901328 | KY464981 | [37] |
| Isolate 4508 | Beauveria bassiana | China | Soil | KX901307 | KX901319 | KX901325 | KY464978 | [37] |
| 2898 T | Beauveria medogensis | China | Soil | KU994837 | KU994833 | KU994835 | KU994834 | [37] |
| BUB426 | Beauveria medogensis | China | Soil | MG642832 | MG642904 | MG642859 | MG642874 | [37] |
| RCEF5500 T | Beauveria lii | China | Henosepilachna vigintioctopunctata | JN689372 | JN689371 | JN689374 | JN689370 | [38] |
| ARSEF 8257 T | Beauveria verroae | France | Varroa destructor | NR111599 | HQ881002 | HQ880872 | HQ880944 | [39] |
| ARSEF 2694 | Beauveria verroae | Switzerland | Larinus sp. | HQ880802 | HQ881004 | HQ880874 | HQ880946 | [39] |
| ARSEF 4598 T | Beauveria australis | Australia | Soil | NR111597 | HQ880995 | HQ880861 | HQ880933 | [39] |
| ARSEF 4580 | Beauveria australis | Australia | Orthoptera: Acrididae | HQ880788 | HQ880994 | HQ880860 | HQ880932 | [39] |
| ARSEF 7032 T | Beauveria kipukae | USA | Not provided | NR111600 | HQ881005 | HQ880875 | HQ880947 | [39] |
| ARSEF 7760 T | Beauveria malawiensis | Malawi | Phoracantha semipunctata | DQ376247 | DQ376246 | HQ880897 | HQ880969 | [40] |
| Bwetak89 | Beauveria malawiensis | New Zealand | Not provided | MW027837 | MW030946 | MW027830 | MW027829 | [35] |
| ARSEF 4850 T | Beauveria asiastica | Korea | Coleoptera: Cerambycidae | NR111596 | KJ523141 | HQ880859 | HQ880931 | [39] |
| BCC13243 | Beauveria asiastica | Thailand | NR | MN401629 | MN401455 | MN401553 | NR | [41] |
| ARSEF 2567 T | Beauveria caledonica | Scotland | Soil | HQ880817 | EF469057 | HQ880889 | HQ880961 | [39] |
| BUB421 | Beauveria caledonica | China | Soil | MG642831 | MG642903 | MG642858 | MG642873 | [39] |
| GZU12141 T | Beauveria majiangensis | China | Coleoptera | MG052642 | MG052640 | MG052644 | NR | [42] |
| GZU12142 | Beauveria majiangensis | China | Coleoptera | MG052643 | MG052641 | MG052645 | NR | [42] |
| ARSEF 617 T | Beauveria brongniartii | France | Coleoptera: Scarabaeidae | NR111595 | HQ880991 | HQ880854 | HQ880926 | [39] |
| ARSEF 7516 | Beauveria brongniartii | Japan | Coleoptera: Scarabaeidae | HQ880766 | HQ880976 | HQ880838 | HQ880910 | [39] |
| Bt99 | Beauveria hoplocheli | Reunion Island | Coleoptera: Melolonthidae | KC339698 | KC339710 | KM453949 | KM453958 | [43] |
| ARSEF 3405 T | Beauveria pseudobassiana | Kentucky, USA | Lepidoptera: Tortricidae | NR111598 | NR | HQ880864 | HQ880936 | [39] |
| ARSEF 1855 | Beauveria pseudobassiana | Canada | Coleoptera: Scolytidae | HQ880796 | HQ880999 | HQ880868 | HQ880940 | [39] |
| ARSEF 2922 T | Beauveria vermiconia | Chile | Soil | NR151832 | NR | HQ880894 | HQ880966 | [44] |
| ARSEF 2641 T | Beauveria amorpha | Brazil | Hymenoptera: Formicidae | NR111601 | NR | HQ880880 | HQ880952 | [39] |
| ARSEF 7542 | Beauveria amorpha | Colorado, USA | Hymenoptera: Formicidae | HQ880805 | HQ881007 | HQ880877 | HQ880949 | [39] |
| CBS 350.85 | Lecanicillium antillanum | Cuba | Hymenomycete: Agaric | MH861888 | DQ522350 | DQ522396 | DQ522450 | [45] |
| ARSEF 4029 | Isaria farinosa | Denmark | Coleoptera: Carabidae | HQ880828 | HQ881019 | HQ880900 | HQ880972 | [39] |
| Isolates | Time (Days) | |||
|---|---|---|---|---|
| 3 | 6 | 9 | 12 | |
| BCMU1 108 | 3.33 ± 1.67 ab* | 5.00 ± 0.00 a | 6.67 ± 1.67 a | 41.67 ± 3.33 c |
| BCMU1 106 | 3.33 ± 3.33 ab | 3.33 ± 3.33 a | 3.33 ± 3.33 a | 10.00 ± 2.89 a |
| BCMU2 108 | 15.00 ± 0.00 bc | 20.00 ± 2.87 b | 31.67 ± 3.33 cd | 41.67 ± 3.33 c |
| BCMU2 106 | 0.00 ± 0.00 a | 3.33 ± 3.33 a | 5.00 ± 2.88 a | 35.00 ± 2.89 bc |
| BCMU3 108 | 36.67 ± 3.33 ef | 40.00 ± 2.87 c | 43.33 ± 3.33 e | 73.33 ± 3.33 ef |
| BCMU3 106 | 3.33 ± 1.67 ab | 11.67 ± 1.67 ab | 11.67 ± 1.67 ab | 55.00 ± 2.89 d |
| BCMU4 108 | 30.00 ± 2.89 de | 53.33 ± 3.33 d | 55.00 ± 2.87 f | 60.00 ± 2.89 d |
| BCMU4 106 | 21.67 ± 1.67 cd | 21.67 ± 1.67 b | 21.67 ± 1.67 bc | 23.33 ± 1.67 b |
| BCMU5 108 | 28.33 ± 4.41 de | 56.67 ± 7.26 d | 66.67 ± 4.41 g | 83.33 ± 6.01 fg |
| BCMU5 106 | 3.33 ± 1.67 ab | 8.33 ± 1.67 ab | 8.33 ± 1.67 a | 35.00 ± 0.00 bc |
| BCMU6 108 | 43.33 ± 6.00 f | 71.67 ± 4.41 e | 76.67 ± 3.33 g | 91.67 ± 1.67 g |
| BCMU6 106 | 15.00 ± 2.87 bc | 23.33 ± 3.33 b | 33.33 ± 1.67 de | 61.67 ± 3.33 de |
| Control | 0.00 a | 0.00 a | 0.00 a | 0.00 a |
| df1 | 12 | 12 | 12 | 12 |
| df2 | 26 | 26 | 26 | 26 |
| F-test 0.05 | 27.798 | 50.594 | 91.074 | 81.081 |
| Fungal Isolate | Length (bp) | GenBank Accession Number | Closeted Species/Accession Number | Similarity (%) |
|---|---|---|---|---|
| BCMU1 | 346 | OL469003 | Beauveria bassiana ARSEF 2860/XM008599737 | 100 |
| BCMU2 | 358 | OL469004 | Beauveria bassiana ARSEF 2860/XM008599737 | 100 |
| BCMU3 | 356 | OL469005 | Beauveria bassiana ARSEF 2860/XM008599737 | 100 |
| BCMU4 | 340 | OL469006 | Beauveria bassiana ARSEF 2860/XM008599737 | 100 |
| BCMU5 | 346 | OL469007 | Beauveria bassiana ARSEF 2860/XM008599737 | 100 |
| BCMU6 | 350 | OL469008 | Beauveria bassiana ARSEF 2860/XM008599737 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajula, J.; Pittarate, S.; Suwannarach, N.; Kumla, J.; Ptaszynska, A.A.; Thungrabeab, M.; Mekchay, S.; Krutmuang, P. Evaluation of Native Entomopathogenic Fungi for the Control of Fall Armyworm (Spodoptera frugiperda) in Thailand: A Sustainable Way for Eco-Friendly Agriculture. J. Fungi 2021, 7, 1073. https://doi.org/10.3390/jof7121073
Rajula J, Pittarate S, Suwannarach N, Kumla J, Ptaszynska AA, Thungrabeab M, Mekchay S, Krutmuang P. Evaluation of Native Entomopathogenic Fungi for the Control of Fall Armyworm (Spodoptera frugiperda) in Thailand: A Sustainable Way for Eco-Friendly Agriculture. Journal of Fungi. 2021; 7(12):1073. https://doi.org/10.3390/jof7121073
Chicago/Turabian StyleRajula, Julius, Sarayut Pittarate, Nakarin Suwannarach, Jaturong Kumla, Aneta A. Ptaszynska, Malee Thungrabeab, Supamit Mekchay, and Patcharin Krutmuang. 2021. "Evaluation of Native Entomopathogenic Fungi for the Control of Fall Armyworm (Spodoptera frugiperda) in Thailand: A Sustainable Way for Eco-Friendly Agriculture" Journal of Fungi 7, no. 12: 1073. https://doi.org/10.3390/jof7121073
APA StyleRajula, J., Pittarate, S., Suwannarach, N., Kumla, J., Ptaszynska, A. A., Thungrabeab, M., Mekchay, S., & Krutmuang, P. (2021). Evaluation of Native Entomopathogenic Fungi for the Control of Fall Armyworm (Spodoptera frugiperda) in Thailand: A Sustainable Way for Eco-Friendly Agriculture. Journal of Fungi, 7(12), 1073. https://doi.org/10.3390/jof7121073