
Fungal Laccases: The Forefront of Enzymes for Sustainability

 ,
,  , ,
, , 
Abstract
1. Introduction
2. Laccase: A Case Study
2.1. Laccase: Overview, Sources
2.2. Physicochemical and Catalytic Properties, Enzyme Stability
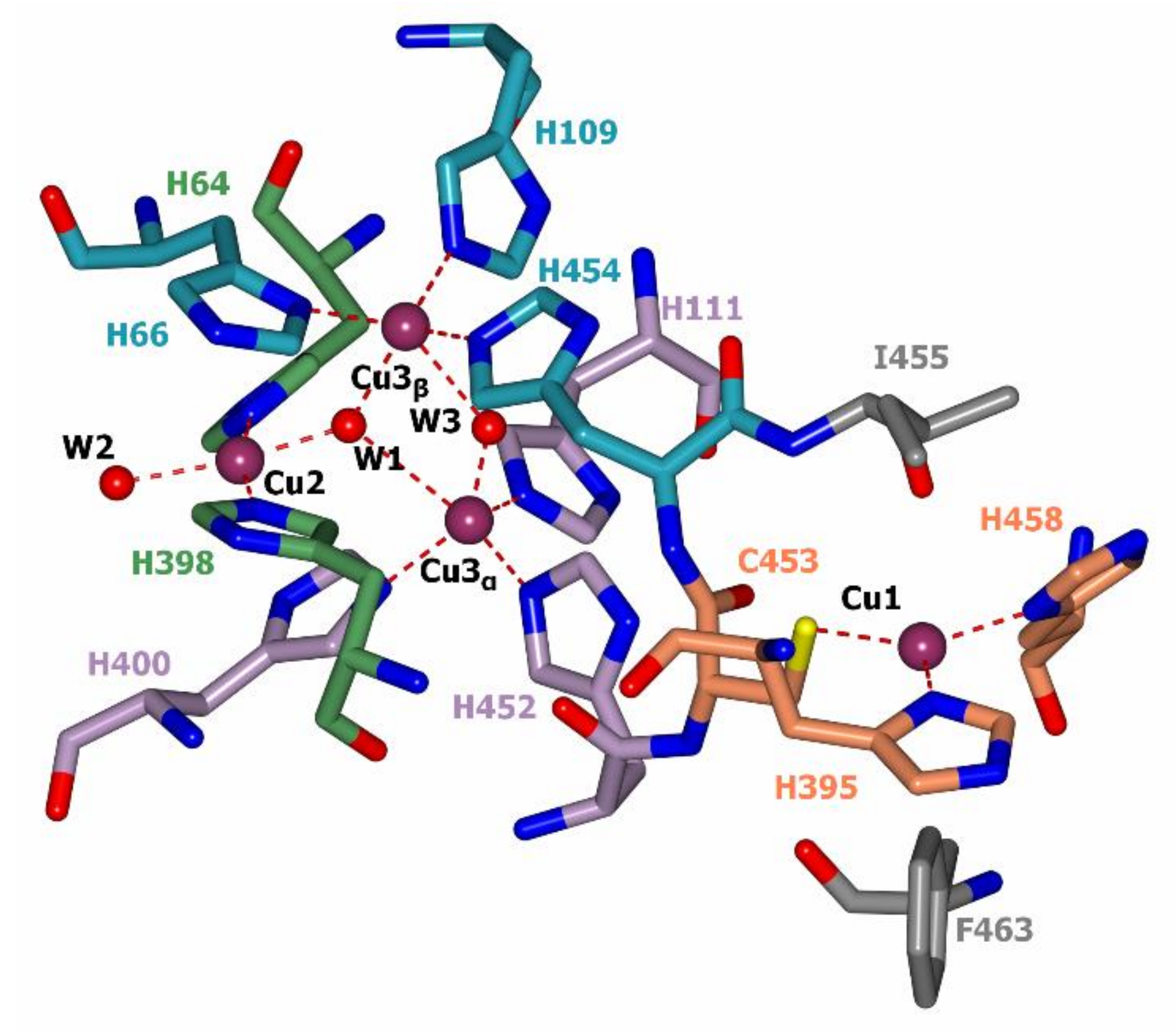
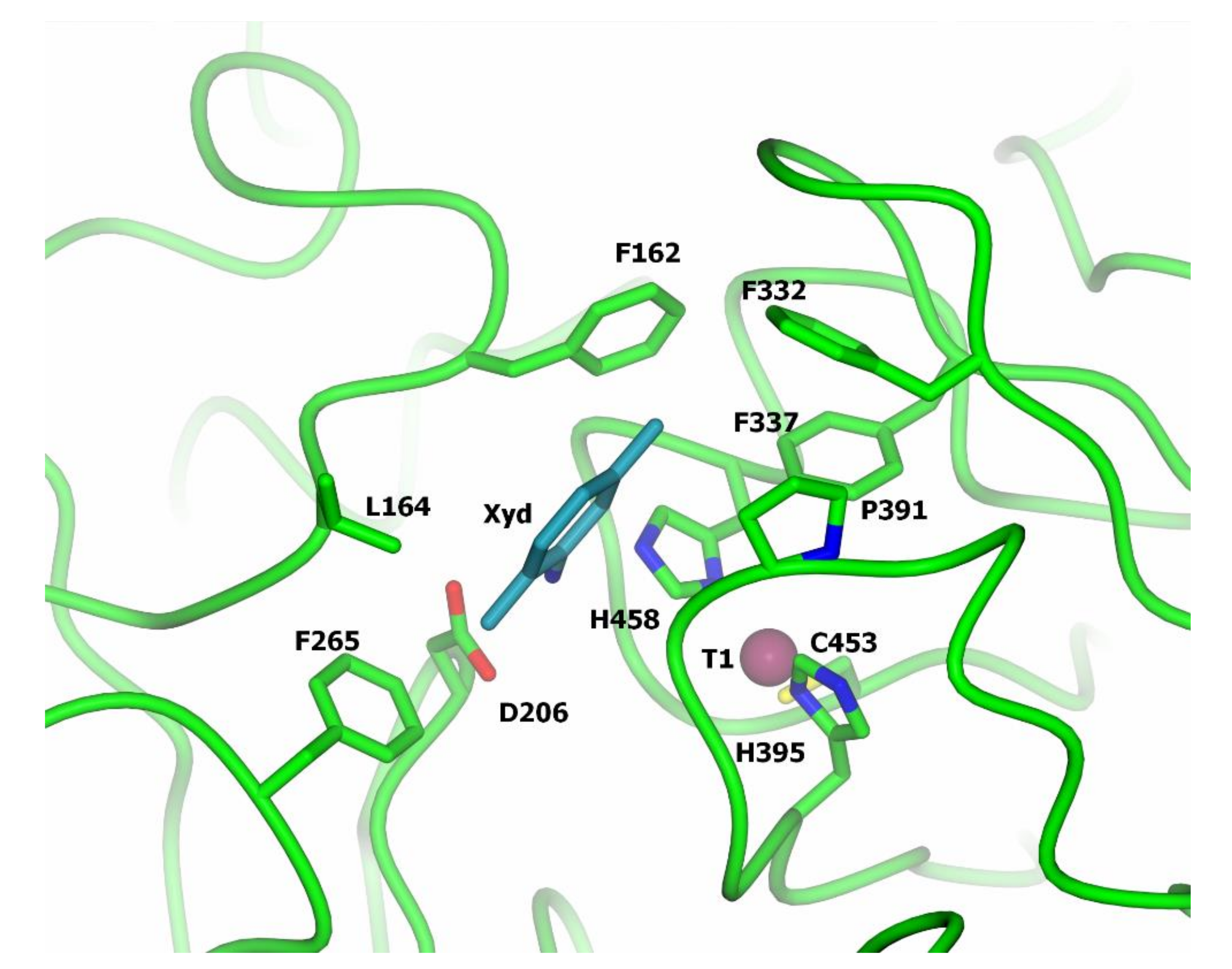
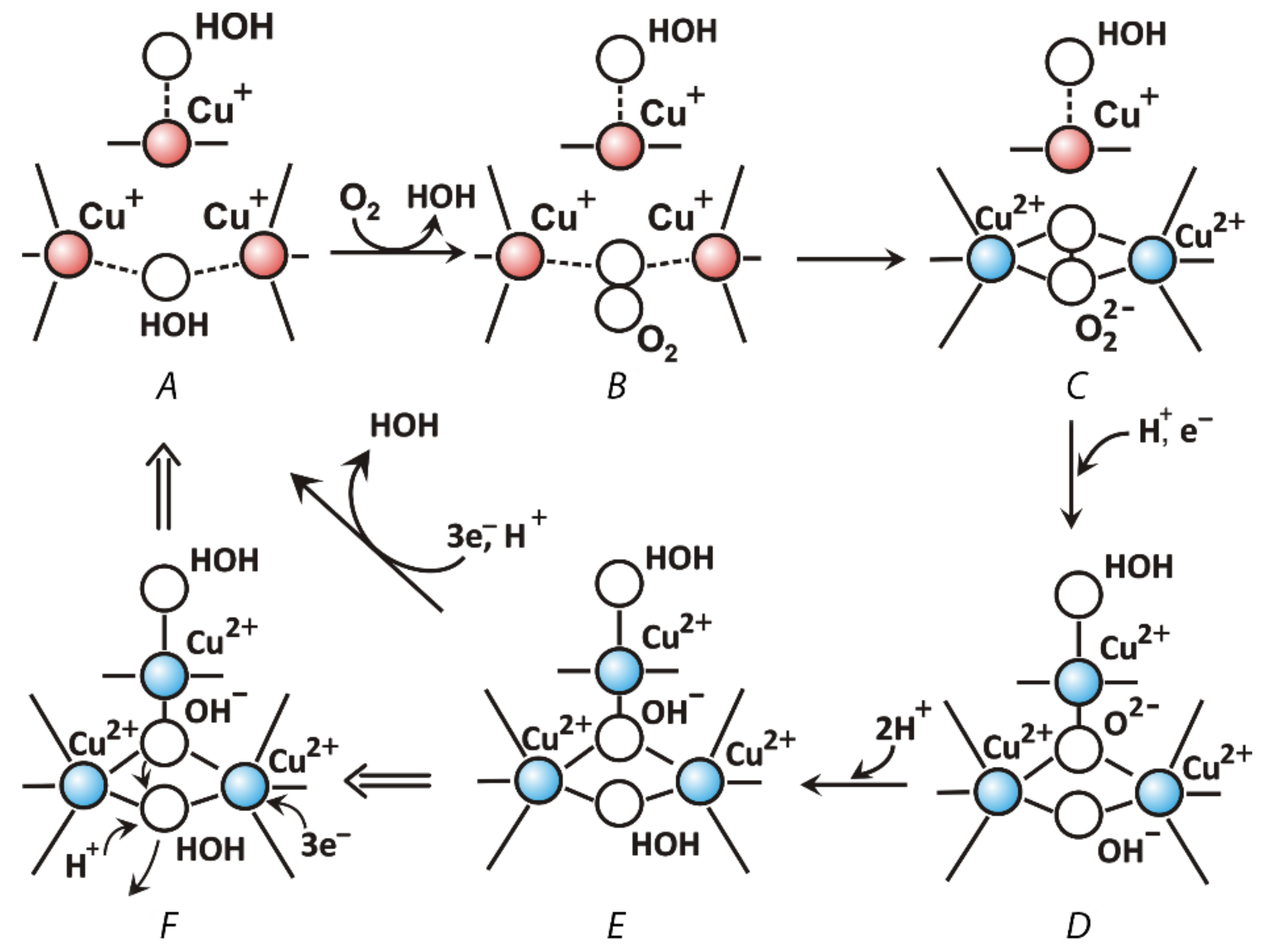
2.3. Laccase Structure and Mechanism of Catalysis
2.4. Laccase Industrial Application
3. Re-Use of Agro-Wastes for Laccase Production
3.1. Laccase Production by Solid State Fermentation Using Agro-Wastes
3.2. Laccase Immobilization on Agro Industrial Wastes
3.3. Valorizarion of Agro-Wastes by Laccase Pretreatment
4. New Trends and Challenges for Laccase Application
4.1. Cold-Active Laccases
4.2. Alkaline Active Laccase
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chapman, J.; Ismail, A.E.; Dinu, C.Z. Industrial applications of enzymes: Recent advances, techniques, and outlooks. Catalysts 2018, 8, 238. [Google Scholar] [CrossRef]
- European Commission. Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions Next Steps for a Sustainable European Future European Action for Sustainability. 2016. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:52016DC0739&from=EN (accessed on 25 February 2021).
- Mika, L.; Csefalvay, E.; Nemeth, A. Catalytic Conversion of Carbohydrates to Initial Platform Chemicals: Chemistry and Sustainability. Chem. Rev. 2018, 118, 505–613. [Google Scholar] [CrossRef] [PubMed]
- Grand View Research. Available online: https://www.grandviewresearch.com/industry-analysis/enzymes-industry (accessed on 8 April 2021).
- Global Market Insight. Available online: https://www.gminsights.com/industry-analysis/enzymes-market (accessed on 8 April 2021).
- Fasim, A.; More, V.S.; More, S.S. Large-scale production of enzymes for biotechnology uses. Curr. Opin. Biotechnol. 2021, 69, 68–76. [Google Scholar] [CrossRef]
- Contesini, F.J.; Frandsen, R.J.N.; Damasio, A. CAZymes in Biorefinery: From Genes to Application. Front. Bioeng. Biotechnol. 2021, 9, 86. [Google Scholar] [CrossRef]
- Malhotra, M.; Suman, S.K. Laccase-mediated delignification and detoxification of lignocellulosic biomass: Removing obstacles in energy generation. Environ. Sci. Pollut. Res. 2021, 28, 58929–58944. [Google Scholar] [CrossRef] [PubMed]
- FAO. The State of Food and Agriculture Moving Forward on Food Loss and Waste Reduction. 2019. Available online: http://www.fao.org/3/ca6030en/ca6030en.pdf (accessed on 2 April 2021).
- Lange, L.; Connor, K.O.; Arason, S.; Bundgård-Jørgensen, U.; Canalis, A.; Carrez, D.; Gallagher, J.; Gøtke, N.; Huyghe, C.; Jarry, B.; et al. Developing a Sustainable and Circular Bio-Based Economy in EU: By Partnering Across Sectors, Upscaling and Using New Knowledge Faster, and For the Benefit of Climate, Environment & Biodiversity, and People & Business. Front. Bioeng. Biotechnol. 2021, 8, 1456. [Google Scholar] [CrossRef]
- Cannatelli, M.D.; Ragauskas, A.J. Two Decades of Laccases: Advancing Sustainability in the Chemical Industry. Chem. Rec. 2017, 17, 122–140. [Google Scholar] [CrossRef]
- Zucca, P.; Cocco, G.; Sollai, F.; Sanjust, E. Fungal laccases as tools for biodegradation of industrial dyes. Biocatalysis 2016, 1, 82–108. [Google Scholar] [CrossRef]
- Moiseenko, K.V.; Savinova, O.S.; Vasina, D.V.; Kononikhin, A.S.; Tyazhelova, T.V.; Fedorova, T.V. Laccase Isoenzymes of Trametes hirsuta LE-BIN072: Degradation of industrial dyes and secretion under the different induction conditions. Appl. Biochem. Biotechnol. 2018, 54, 834–841. [Google Scholar] [CrossRef]
- Zhang, Y.; Lin, D.F.; Hao, J.; Zhao, Z.H.; Zhang, Y.J. The crucial role of bacterial laccases in the bioremediation of petroleum hydrocarbons. World J. Microbiol. Biotechnol. 2020, 36, 1–10. [Google Scholar] [CrossRef]
- Barrios-Estrada, C.; de Jesús Rostro-Alanis, M.; Muñoz-Gutiérrez, B.D.; Iqbal, H.M.; Kannan, S.; Parra-Saldívar, R. Emergent contaminants: Endocrine disruptors and their laccase-assisted degradation—A review. Sci. Total. Environ. 2018, 612, 1516–1531. [Google Scholar] [CrossRef]
- Loi, M.; Fanelli, F.; Zucca, P.; Liuzzi, V.C.; Quintieri, L.; Cimmarusti, M.T.; Monaci, L.; Haidukowski, M.; Logrieco, A.F.; Sanjust, E.; et al. Aflatoxin B1 and M1 degradation by Lac2 from Pleurotus pulmonarius and redox mediators. Toxins 2016, 8, 245. [Google Scholar] [CrossRef]
- Yoshida, H. Chemistry of lacquer (urushi). J. Chem. Soc. 1883, 43, 472–486. [Google Scholar] [CrossRef]
- Bertrand, G. Simultaneous occurence of laccase and tyrosinase in the juice of some mushrooms. C. R. Hebd. Séances Acad. Sci. 1896, 123, 463–465. [Google Scholar]
- Giardina, P.; Faraco, V.; Pezzella, C.; Piscitelli, A.; Vanhulle, S.; Sannia, G. Laccases: A never-ending story. Cell. Mol. Life Sci. 2010, 67, 369–385. [Google Scholar] [CrossRef]
- Hoegger, P.J.; Kilaru, S.; James, T.Y.; Thacker, J.R.; K??es, U. Phylogenetic comparison and classification of laccase and related multicopper oxidase protein sequences. FEBS J. 2006, 273, 2308–2326. [Google Scholar] [CrossRef] [PubMed]
- Munk, L.; Sitarz, A.K.; Kalyani, D.C.; Mikkelsen, J.D.; Meyer, A.S. Can laccases catalyze bond cleavage in lignin? Biotechnol. Adv. 2015, 33, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Alcalde, M. Laccases: Biological functions, molecular structure and industrial applications. In Industrial Enzymes; Springer: Dordrecht, The Netherlands, 2007; pp. 461–476. ISBN 978-1-4020-5376-4. [Google Scholar]
- Sharma, P.; Goel, R.; Capalash, N. Bacterial laccases. World J. Microbiol. Biotechnol. 2007, 23, 823–832. [Google Scholar] [CrossRef]
- Nordberg, H.; Cantor, M.; Dusheyko, S.; Hua, S.; Poliakov, A.; Shabalov, I.; Dubchak, I. The genome portal of the Department of Energy Joint Genome Institute: 2014 updates. Nucleic Acids Res. 2014, 42, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Piscitelli, A.; Giardina, P.; Lettera, V.; Pezzella, C.; Sannia, G.; Faraco, V. Induction and transcriptional regulation of laccases in fungi. Curr. Genom. 2011, 12, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.M.; Solomon, E.I. Electron transfer and reaction mechanism of laccases. Cell. Mol. Life Sci. 2015, 72, 869–883. [Google Scholar] [CrossRef]
- Xu, F. Oxidation of phenols, anilines, and benzenethiols by fungal laccases: Correlation between activity and redox potentials as well as halide inhibition. Biochemistry 1996, 35, 7608–7614. [Google Scholar] [CrossRef] [PubMed]
- Glazunova, O.A.; Trushkin, N.A.; Moiseenko, K.V.; Filimonov, I.S.; Fedorova, T.V. Catalytic efficiency of basidiomycete laccases: Redox potential versus substrate-binding pocket structure. Catalysts 2018, 8, 152. [Google Scholar] [CrossRef]
- Frasconi, M.; Favero, G.; Boer, H.; Koivula, A.; Mazzei, F. Kinetic and biochemical properties of high and low redox potential laccases from fungal and plant origin. Biochim. Biophys. Acta 2010, 1804, 899–908. [Google Scholar] [CrossRef] [PubMed]
- Lahtinen, M.; Kruus, K.; Boer, H.; Kemell, M.; Andberg, M.; Viikari, L. The effect of lignin model compound structure on the rate of oxidation catalyzed by two different fungal laccases. J. Mol. Catal. 2009, 57, 204–210. [Google Scholar] [CrossRef]
- Xu, F.; Berka, R.M.; Wahleithner, J.; Nelson, B. Site-directed mutations in fungal laccase: Effect on redox potential, activity and pH profile. Biochem. J. 1998, 70, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Hoyos, C.M.; Morales-Alvarez, E.D.; Poutou-Pinales, R.A.; Pedroza-Rodriguez, A.M.; Rodriguez-Vazquez, R.; Delgado-Boada, J.M. Fungal laccases. Fungal Biol. Rev. 2013, 27, 67–82. [Google Scholar] [CrossRef]
- Mate, D.M.; Alcalde, M. Laccase engineering: From rational design to directed evolution. Biotechnol. Adv. 2015, 33, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Glazunova, O.A.; Polyakov, K.M.; Moiseenko, K.V.; Kurzeev, S.A.; Fedorova, T.V. Structure-function study of two new middle-redox potential laccases from basidiomycetes Antrodiella faginea and Steccherinum murashkinskyi. Int. J. Biol. Macromol. 2018, 118, 406–418. [Google Scholar] [CrossRef]
- Rodgers, C.J.; Blanford, C.F.; Giddens, S.R.; Skamnioti, P.; Armstrong, F.A.; Gurr, S.J. Designer laccases: A vogue for high-potential fungal enzymes? Trends Biotechnol. 2010, 28, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Shleev, S.; Christenson, A.; Serezhenkov, V.; Burbaev, D.; Yaropolov, A.; Gorton, L.; Ruzgas, T. Electrochemical redox transformations of T1 and T2 copper sites in native Trametes hirsuta laccase at gold electrode. Biochem. J. 2005, 385, 745–754. [Google Scholar] [CrossRef]
- Reinhammar, B.R.M. Oxidation-reduction potentials of the electron acceptors in laccases and stellacyanin. BBA—Bioenerg. 1972, 275, 245–259. [Google Scholar] [CrossRef]
- Farver, O.; Wherland, S.; Koroleva, O.; Loginov, D.S.; Pecht, I. Intramolecular electron transfer in laccases. FEBS J. 2011, 278, 3463–3471. [Google Scholar] [CrossRef]
- Morozova, O.V.; Shumakovich, G.P.; Gorbacheva, M.A.; Shleev, S.V.; Yaropolov, A.I. “Blue” laccases. Biochemistry 2007, 72, 1136–1150. [Google Scholar] [CrossRef] [PubMed]
- Baldrian, P. Fungal laccases—Occurrence and properties. FEMS Microbiol. Rev. 2006, 30, 215–242. [Google Scholar] [CrossRef] [PubMed]
- Claus, H. Laccases: Structure, reactions, distribution. Micron 2004, 35, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Hildén, K.; Hakala, T.K.; Lundell, T. Thermotolerant and thermostable laccases. Biotechnol. Lett. 2009, 31, 1117–1128. [Google Scholar] [CrossRef]
- Li, K.; Xu, F.; Eriksson, K.E.L. Comparison of fungal laccases and redox mediators in oxidation of a nonphenolic lignin model compound. Appl. Environ. Microbiol. 1999, 65, 2654–2660. [Google Scholar] [CrossRef]
- Xu, Y.; Hong, Y.; Xiao, Y.; Fang, W. Preparation and application of polyclonal antibody against a recombinant laccase. Cell. Mol. Immunol. 2007, 4, 315–317. [Google Scholar] [PubMed]
- Vite-Vallejo, O.; Palomares, L.A.; Dantán-González, E.; Ayala-Castro, H.G.; Martínez-Anaya, C.; Valderrama, B.; Folch-Mallol, J. The role of N-glycosylation on the enzymatic activity of a Pycnoporus sanguineus laccase. Enzyme Microb. Technol. 2009, 45, 233–239. [Google Scholar] [CrossRef]
- Maestre-Reyna, M.; Liu, W.C.; Jeng, W.Y.; Lee, C.C.; Hsu, C.A.; Wen, T.N.; Wang, A.H.J.; Shyur, L.F. Structural and functional roles of glycosylation in fungal laccase from Lentinus sp. PLoS ONE 2015, 10, e0120601. [Google Scholar] [CrossRef]
- Glazunova, O.A.; Moiseenko, K.V.; Kamenihina, I.A.; Isaykina, T.U.; Yaropolov, A.I.; Fedorova, T.V. Laccases with variable properties from different strains of Steccherinum ochraceum: Does glycosylation matter? Int. J. Mol. Sci. 2019, 20, 2008. [Google Scholar] [CrossRef]
- Lisova, Z.A.; Lisov, A.V.; Leontievsky, A.A. Two laccase isoforms of the basidiomycete Cerrena unicolor VKMF-3196. Induction, isolation and properties. J. Basic Microbiol. 2010, 50, 72–82. [Google Scholar] [CrossRef]
- Васина, Д.В.; Логинов, Д.С.; Королева, О.В. Сравнительный анализ протеома базидиального гриба Trametes hirsuta при культивировании на средах различного состава. Биохимия 2013, 78, 627–636. [Google Scholar]
- Glazunova, O.A.; Moiseenko, K.V.; Savinova, O.S.; Fedorova, T.V. Purification and Characterization of Two Novel Laccases from Peniophora lycii. J. Fungi 2020, 6, 340. [Google Scholar] [CrossRef] [PubMed]
- Garzillo, A.M.; Colao, M.C.; Caruso, C. Laccase from the white-rot fungus Trametes trogii. Appl. Microbiol. Biotechnol. 1998, 49, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Reiss, R.; Ihssen, J.; Richter, M.; Eichhorn, E.; Schilling, B.; Thöny-Meyer, L. Laccase versus laccase-like multi-copper oxidase: A comparative study of similar enzymes with diverse substrate spectra. PLoS ONE 2013, 8, e65633. [Google Scholar] [CrossRef]
- Astolti, P.; Brandi, P.; Galli, C.; Gentili, P.; Gerini, M.F.; Greci, L.; Lanzalunga, O. New mediators for the enzyme laccase: Mechanistic features and selectivity in the oxidation of non-phenolic substrates. New J. Chem. 2005, 29, 1308–1317. [Google Scholar] [CrossRef]
- Lange, H.; Decina, S.; Crestini, C. Oxidative upgrade of lignin—Recent routes reviewed. Eur. Polym. J. 2013, 49, 1151–1173. [Google Scholar] [CrossRef]
- D’Acunzo, F.; Galli, C.; Gentili, P.; Sergi, F. Mechanistic and steric issues in the oxidation of phenolic and non-phenolic compounds by laccase or laccase-mediator systems. the case of bifunctional substrates. New J. Chem. 2006, 30, 583–591. [Google Scholar] [CrossRef]
- Munk, L.; Andersen, M.L.; Meyer, A.S. Influence of mediators on laccase catalyzed radical formation in lignin. Enzyme Microb. Technol. 2018, 116, 48–56. [Google Scholar] [CrossRef]
- Kumar, S.V.S.; Phale, P.S.; Durani, S.; Wangikar, P.P. Combined sequence and structure analysis of the fungal laccase family. Biotechnol. Bioeng. 2003, 83, 386–394. [Google Scholar] [CrossRef]
- Ducros, V.; Brzozowski, A.M.; Wilson, K.S.; Brown, S.H.; Ostergaard, P.; Shneider, P.; Yaver, D.S.; Pedersen, A.H.; Davies, G.J. Crystal structure of type-2 Cu depleted laccase from Coprinus Cinereus at 2,2 А resolution. Nat. Struct. Biol. 1998, 5, 310–316. [Google Scholar] [CrossRef]
- Hakulinen, N.; Rouvinen, J. Three-dimensional structures of laccases. Cell. Mol. Life Sci. 2015, 72, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Glazunova, O.A.; Polyakov, K.M.; Fedorova, T.V.; Dorovatovskii, P.V.; Koroleva, O.V. Elucidation of the crystal structure of Coriolopsis caperata laccase: Restoration of the structure and activity of the native enzyme from the T2-depleted form by copper ions. Acta Crystallogr. Sect. D Biol. Crystallogr. 2015, 71, 854–861. [Google Scholar] [CrossRef]
- Polyakov, K.M.; Fedorova, T.V.; Stepanova, E.V.; Cherkashin, E.A.; Kurzeev, S.A.; Strokopytov, B.V.; Lamzin, V.S.; Koroleva, O.V. Structure of native laccase from Trametes hirsuta at 1.8 A resolution. Acta Crystallogr. D Biol. Crystallogr. 2009, 65, 611–617. [Google Scholar] [CrossRef] [PubMed]
- Durão, P.; Chen, Z.; Silva, C.S.; Soares, C.M.; Pereira, M.M.; Todorovic, S.; Hildebrandt, P.; Bento, I.; Lindley, P.F.; Martins, L.O. Proximal mutations at the type 1 copper site of CotA laccase: Spectroscopic, redox, kinetic and structural characterization of I494A and L386A mutants. Biochem. J. 2008, 412, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Osipov, E.; Polyakov, K.; Kittl, R.; Shleev, S.; Dorovatovsky, P.; Tikhonova, T.; Hann, S.; Ludwig, R.; Popov, V. Effect of the L499M mutation of the ascomycetous Botrytis aclada laccase on redox potential and catalytic properties. Acta Crystallogr. Sect. D Biol. Crystallogr. 2014, 70, 2913–2923. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Palmer, A.E.; Yaver, D.S.; Berka, R.M.; Gambetta, G.A.; Brown, S.H.; Solomon, E.I. Targeted mutations in a Trametes villosa laccase. J. Biol. Chem. 1999, 274, 12372–12375. [Google Scholar] [CrossRef]
- Matera, I.; Gullotto, A.; Tilli, S.; Ferraroni, M.; Scozzafava, A.; Briganti, F. Crystal structure of the blue multicopper oxidase from the white-rot fungus Trametes trogii complexed with p-toluate. Inorg. Chim. Acta 2008, 361, 4129–4137. [Google Scholar] [CrossRef]
- Macellaro, G.; Baratto, M.C.; Piscitelli, A.; Pezzella, C.; Fabrizi De Biani, F.; Palmese, A.; Piumi, F.; Record, E.; Basosi, R.; Sannia, G. Effective mutations in a high redox potential laccase from Pleurotus ostreatus. Appl. Microbiol. Biotechnol. 2014, 98, 4949–4961. [Google Scholar] [CrossRef]
- Kallio, J.; Auer, S.; Jänis, J.; Andberg, M.; Kruus, K.; Rouvinen, J.; Koivula, A.; Hakulinen, N. Structure–function studies of a Melanocarpus albomyces laccase suggest a pathway for oxidation of phenolic compounds. J. Mol. Biol. 2009, 392, 895–909. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, T.; Jolivalt, C.; Briozzo, P.; Caminade, E.; Joly, N.; Madzak, C.; Mougin, C. Crystal structure of a four-copper laccase complexed with an arylamine: Insights into substrate recognition and correlation with kinetics. Biochemistry 2002, 41, 7325–7333. [Google Scholar] [CrossRef] [PubMed]
- Xie, T.; Liu, Z.; Liu, Q.; Wang, G. Structural insight into the oxidation of sinapic acid by CotA laccase. J. Struct. Biol. 2015, 190, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Enguita, F.J.; Marçal, D.; Martins, L.O.; Grenha, R.; Henriques, A.O.; Lindley, P.F.; Carrondo, M.A. Substrate and dioxygen binding to the endospore coat laccase from Bacillus subtilis. J. Biol. Chem. 2004, 279, 23472–23476. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Xie, T.; Zhong, Q.; Wang, G. Crystal structure of CotA laccase complexed with 2,2-azinobis-(3-ethylbenzothiazoline-6-sulfonate) at a novel binding site. Acta Crystallogr. Sect. Struct. Biol. Commun. 2016, 72, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.L.; Clark, P.A.; Solomon, E.I. Spectroscopic and chemical studies of the laccase trinuclear copper active site: Geometric and electronic structure. J. Am. Chem. Soc. 1990, 112, 9534–9548. [Google Scholar] [CrossRef]
- Komori, H.; Sugiyama, R.; Kataoka, K.; Miyazaki, K.; Higuchi, Y.; Sakurai, T. New insights into the catalytic active-site structure of multicopper oxidases. Acta Crystallogr. Sect. D Biol. Crystallogr. 2014, 70, 772–779. [Google Scholar] [CrossRef] [PubMed]
- Polyakov, K.M.M.; Gavryushov, S.; Ivanova, S.; Fedorova, T.V.V.; Glazunova, O.A.A.; Popov, A.N.N.; Koroleva, O.V.V. Structural study of the X-ray-induced enzymatic reduction of molecular oxygen to water by Steccherinum murashkinskyi laccase: Insights into the reaction mechanism. Acta Crystallogr. Sect. D Struct. Biol. 2017, 73, 388–401. [Google Scholar] [CrossRef] [PubMed]
- Polyakov, K.M.; Gavryushov, S.; Fedorova, T.V.; Glazunova, O.A.; Popov, A.N. The subatomic resolution study of laccase inhibition by chloride and fluoride anions using single-crystal serial crystallography: Insights into the enzymatic reaction mechanism. Acta Crystallogr. Sect. D Struct. Biol. 2019, 75, 804–816. [Google Scholar] [CrossRef] [PubMed]
- Moreno, A.D.; Ibarra, D.; Eugenio, M.E.; Tomás-Pejó, E. Laccases as versatile enzymes: From industrial uses to novel applications. J. Chem. Technol. Biotechnol. 2020, 95, 481–494. [Google Scholar] [CrossRef]
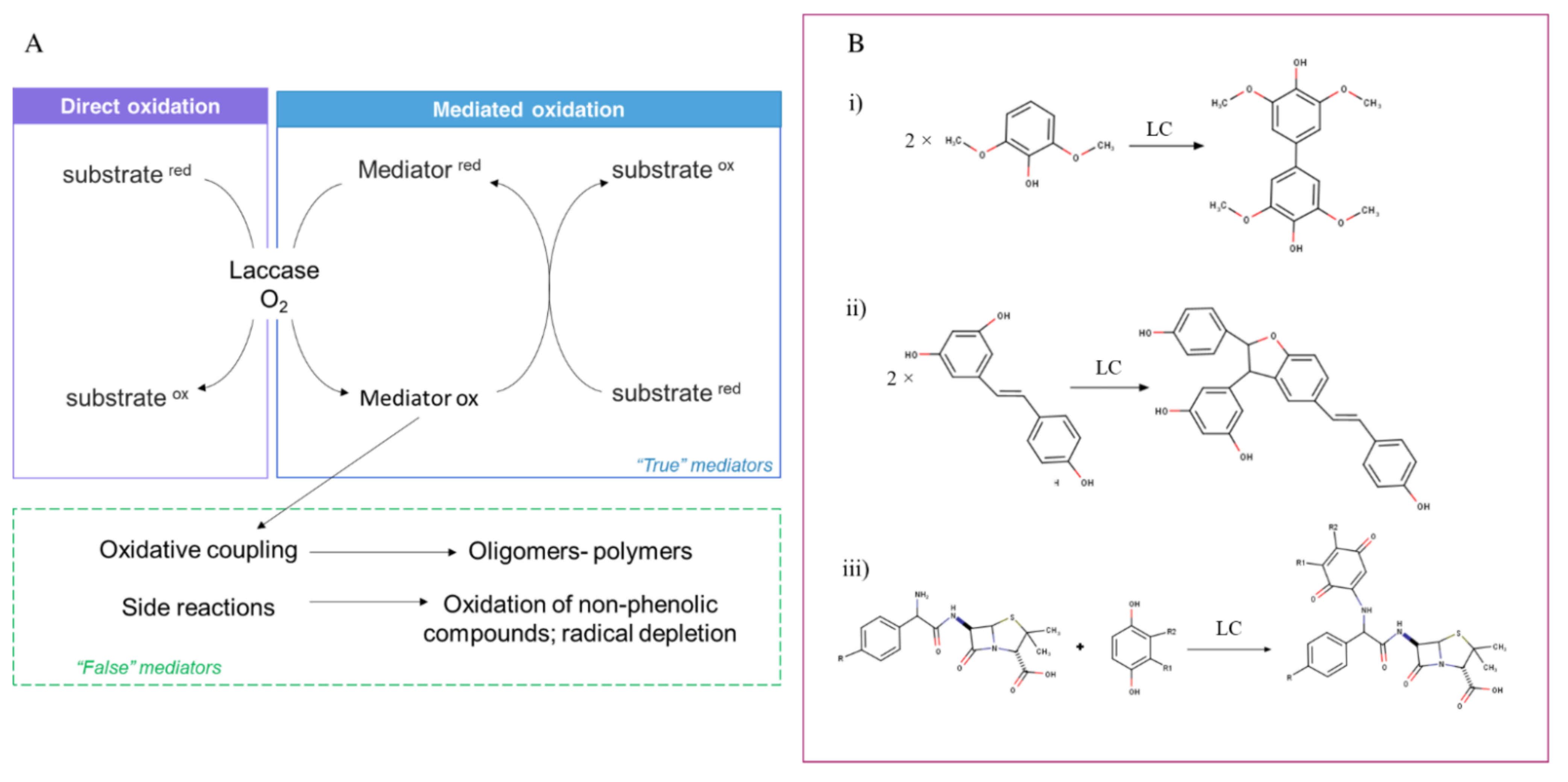
- Morozova, O.V.; Shumakovich, G.P.; Shleev, S.V.; Yaropolov, Y.I. Laccase-mediator systems and their applications: A review. Appl. Biochem. Microbiol. 2007, 43, 523–535. [Google Scholar] [CrossRef]
- Cañas, A.I.; Camarero, S. Laccases and their natural mediators: Biotechnological tools for sustainable eco-friendly processes. Biotechnol. Adv. 2010, 28, 694–705. [Google Scholar] [CrossRef]
- Yesilada, O.; Birhanli, E.; Geckil, H. Bioremediation and Decolorization of Textile Dyes by White Rot Fungi and Laccase Enzymes. In Mycoremediation and Environmental Sustainability, 1st ed.; Prasad, R., Ed.; Springer: Cham, Switzerland, 2018; Volume 2, pp. 121–153. [Google Scholar] [CrossRef]
- Dhillon, G.S.; Kaur, S.; Brar, S.K.; Verma, M. Flocculation and haze removal from crude beer using in-house produced laccase from Trametes versicolor cultured on brewer’s spent grain. J. Agric. Food Chem. 2012, 60, 7895–7904. [Google Scholar] [CrossRef] [PubMed]
- Lettera, V.; Pezzella, C.; Cicatiello, P.; Piscitelli, A.; Giacobelli, V.G.; Galano, E.; Amoresano, A.; Sannia, G. fficient immobilization of a fungal laccase and its exploitation in fruit juice clarification. Food Chem. 2016, 196, 1272–1278. [Google Scholar] [CrossRef]
- Loi, M.; Quintieri, L.; Fanelli, F.; Caputo, L.; Mulè, G. Application of a recombinant laccase-chlorogenic acid system in protein crosslink and antioxidant properties of the curd. Food Res. Int. 2018, 106, 763–770. [Google Scholar] [CrossRef]
- Loi, M.; Quintieri, L.; De Angelis, E.; Monaci, L.; Logrieco, A.F.; Caputo, L.; Mule, G. Yield improvement of the Italian fresh Giuncata cheese by laccase–induced protein crosslink. Int. Dairy J. 2020, 100, 104555. [Google Scholar] [CrossRef]
- Li, M.; Karboune, S.; Light, K.; Kermasha, S. Oxidative cross-linking of potato proteins by fungal laccases: Reaction kinetics and effects on the structural and functional properties. Innov. Food Sci. Emerg. Technol. 2021, 71, 102723. [Google Scholar] [CrossRef]
- Zhu, Y.; Tao, H.; Janaswamy, S.; Zou, F.; Cui, B.; Guo, L. The functionality of laccase-or peroxidase-treated potato flour: Role of interactions between protein and protein/starch. Food Chem. 2021, 341, 128082. [Google Scholar] [CrossRef]
- Manhivi, V.E.; Amonsou, E.O.; Kudanga, T. Laccase-mediated crosslinking of gluten-free amadumbe flour improves rheological properties. Food Chem. 2018, 264, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Loi, M.; Fanelli, F.; Cimmarusti, M.T.; Mirabelli, V.; Haidukowski, M.; Logrieco, A.F.; Caliandro, R.; Mule, G. In vitro single and combined mycotoxins degradation by Ery4 laccase from Pleurotus eryngii and redox mediators. Food Control 2018, 90, 401–406. [Google Scholar] [CrossRef]
- Bilal, M.; Rasheed, T.; Nabeel, F.; Iqbal, H.M.; Zhao, Y. Hazardous contaminants in the environment and their laccase-assisted degradation–a review. J. Env Manag. 2019, 234, 253–264. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Du, H.; Peng, X.; Tang, Y.; Zhou, Y.; Chen, X.; Fei, J.; Meng, Y.; Yuan, L. Degradation of several polycyclic aromatic hydrocarbons by laccase in reverse micelle system. Sci. Total Environ. 2020, 708, 134970. [Google Scholar] [CrossRef]
- Kudanga, T.; Nemadziva, B.; Le Roes-Hill, M. Laccase catalysis for the synthesis of bioactive compounds. Appl. Microbiol. Biotechnol. 2017, 101, 13–33. [Google Scholar] [CrossRef]
- Bhusainahalli, V.M.; Spatafora, C.; Chalal, M.; Vervandier-Fasseur, D.; Meunier, P.; Latruffe, N.; Tringali, C. Resveratrol-Related Dehydrodimers: Laccase—Mediated Biomimetic Synthesis and Antiproliferative Activity. Eur. J. Org. Chem. 2012, 27, 5217–5224. [Google Scholar] [CrossRef]
- Sun, K.; Cheng, X.; Yu, J.; Chen, L.; Wei, J.; Chen, W.; Wang, J.; Li, S.; Liu, Q.; Si, Y. Isolation of Trametes hirsuta La-7 with high laccase-productivity and its application in metabolism of 17β-estradiol. Environ. Pollut. 2020, 263, 114381. [Google Scholar] [CrossRef] [PubMed]
- Mikolasch, A.; Manda, K.; Schlüter, R.; Lalk, M.; Witt, S.; Seefeldt, S.; Lindequist, U. Comparative analyses of laccase-catalyzed amination reactions for production of novel β-lactam antibiotics. Biotechnol. Appl. Biochem. 2012, 59, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Mikolasch, A.; Hammer, E.; Witt, S.; Lindequist, U. Laccase-catalyzed derivatization of 6-aminopenicillanic, 7-aminocephalosporanic and 7-aminodesacetoxycephalosporanic acid. AMB Express 2020, 10, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lapinsonnière, L.; Picot, M.; Barrière, F. Enzymatic versus Microbial Bio-Catalyzed Electrodes in Bio-Electrochemical Systems. ChemSusChem 2012, 5, 995–1005. [Google Scholar] [CrossRef] [PubMed]
- Rangelov, S.; Nicell, J.A. Modelling the transient kinetics of laccase-catalyzed oxidation of four aqueous phenolic substrates at low concentrations. Biochem. Eng. J. 2018, 132, 233–243. [Google Scholar] [CrossRef]
- Slagman, S.; Zuilhof, H.; Franssen, M.C. Laccase-mediated grafting on biopolymers and synthetic polymers: A critical review. ChemBioChem 2018, 19, 288. [Google Scholar] [CrossRef] [PubMed]
- Meza, J.C.; Auria, R.; Lomascolo, A.; Sigoillot, J.C.; Casalot, L. Role of ethanol on growth, laccase production and protease activity in Pycnoporus cinnabarinus ss3. Enzym. Microb. Technol. 2007, 41, 162–168. [Google Scholar] [CrossRef]
- Rodríguez-Couto, S. Solid-State Fermentation for Laccases Production and Their Applications. In Current Developments in Biotechnology and Bioengineering, 1st ed.; Pandey, A., Larroche, C., Socco, C.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 211–234. [Google Scholar] [CrossRef]
- Rodrigues, E.M.; Karp, S.G.; Malucelli, L.C.; Helm, C.V.; Alvarez, T.M. Evaluation of laccase production by Ganoderma lucidum in submerged and solid-state fermentation using different inducers. J. Basic Microb. 2019, 59, 784–791. [Google Scholar] [CrossRef]
- Wang, F.; Xu, L.; Zhao, L.; Ding, Z.; Ma, H.; Terry, N. Fungal Laccase Production from Lignocellulosic Agricultural Wastes by Solid-State Fermentation: A Review. Microorganisms 2019, 7, 665. [Google Scholar] [CrossRef] [PubMed]
- Del Cerro, C.; Erickson, E.; Dong, T.; Wong, A.R.; Eder, E.K.; Purvine, S.O.; Mitchell, H.D.; Weitz, K.K.; Markillie, K.M.; Burnet, M.C.; et al. Intracellular pathways for lignin catabolism in white-rot fungi. Proc. Natl. Acad. Sci. USA 2021, 118. [Google Scholar] [CrossRef]
- De Castro, R.J.S.; Sato, H.H. Enzyme production by solid state fermentation: General aspects and an analysis of the physicochemical characteristics of substrates for agro-industrial wastes valorization. Waste Biomass Valoriz. 2015, 6, 1085–1093. [Google Scholar] [CrossRef]
- Isanapong, J.; Krailoekpaiboon, T.; Noiniyom, W.; Panchal, S. Utilization of Organic Wastes for Laccase Production by Pleurotus ostreatus. Appl. Sci. Eng. Progr. 2017, 10, 239–244. [Google Scholar] [CrossRef][Green Version]
- Sánchez, Ó.J.; Montoya, S. Assessment of polysaccharide and biomass production from three white-rot fungi by solid-state fermentation using wood and agro-industrial residues: A kinetic approach. Forests 2020, 11, 1055. [Google Scholar] [CrossRef]
- Arora, S.; Rani, R.; Ghosh, S. Bioreactors in solid state fermentation technology: Design, applications and engineering aspects. J. Biotech. 2018, 269, 16–34. [Google Scholar] [CrossRef] [PubMed]
- Tišma, M.; Jurić, A.; Bucić-Kojić, A.; Panjičko, M.; Planinić, M. Biovalorization of brewers’ spent grain for the production of laccase and polyphenols. J. Inst. Brew. 2018, 124, 182–186. [Google Scholar] [CrossRef]
- Dhillon, G.S.; Kaur, S.; Brar, S.K. In-vitro decolorization of recalcitrant dyes through an ecofriendly approach using laccase from Trametes versicolor grown on brewer’s spent grain. Int. Biodeter. Biodegr. 2012, 72, 67–75. [Google Scholar] [CrossRef]
- Philippoussis, A.; Diamantopoulou, P.; Papadopoulou, K.; Lakhtar, H.; Roussos, S.; Parissopoulos, G.; Papanikolaou, S. Biomass, laccase and endoglucanase production by Lentinula edodes during solid state fermentation of reed grass, bean stalks and wheat straw residues. World J. Microb. Biotechnol. 2011, 27, 285–297. [Google Scholar] [CrossRef]
- Adekunle, A.E.; Zhang, C.; Guo, C.; Liu, C.Z. Laccase production from Trametes versicolor in solid-state fermentation of steam-exploded pretreated cornstalk. Waste Biomass Valoriz. 2017, 8, 153–159. [Google Scholar] [CrossRef]
- Li, G.; Fu, Y.; Dang, W.; Hu, R.; Xue, H. The effects of aqueous ammonia-pretreated rice straw as solid substrate on laccase production by solid-state fermentation. Bioprocess Biosyst. Eng. 2019, 42, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Patel, H.; Gupte, A. Optimization of different culture conditions for enhanced laccase production and its purification from Tricholoma giganteum AGHP. Bioresour. Bioprocess. 2016, 3, 1–10. [Google Scholar] [CrossRef]
- Mishra, V.; Jana, A.K. Sweet sorghum bagasse pretreatment by Coriolus versicolor in mesh tray bioreactor for selective delignification and improved saccharification. Waste Biomass Valoriz. 2019, 10, 2689–2702. [Google Scholar] [CrossRef]
- Xu, L.; Sun, K.; Wang, F.; Zhao, L.; Hu, J.; Ma, H.; Ding, Z. Laccase production by Trametes versicolor in solid-state fermentation using tea residues as substrate and its application in dye decolorization. J. Environ. Manag. 2020, 270, 110904. [Google Scholar] [CrossRef]
- Pourkhanali, K.; Khayati, G.; Mizani, F.; Raouf, F. Isolation, identification and optimization of enhanced production of laccase from Galactomyces geotrichum under solid-state fermentation. Prep. Biochem. Biotechnol. 2021, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Bernal, C.; Rodriguez, K.; Martinez, R. Integrating enzyme immobilization and protein engineering: An alternative path for the development of novel and improved industrial biocatalysts. Biotech. Adv. 2018, 36, 1470–1480. [Google Scholar] [CrossRef] [PubMed]
- Sigurdardóttir, S.B.; Lehmann, J.; Ovtar, S.; Grivel, J.C.; Negra, M.D.; Kaiser, A.; Pinelo, M. Enzyme immobilization on inorganic surfaces for membrane reactor applications: Mass transfer challenges, enzyme leakage and reuse of materials. Adv. Synth. Catal. 2018, 360, 2578–2607. [Google Scholar] [CrossRef]
- Daronch, N.A.; Kelbert, M.; Pereira, C.S.; de Araújo, P.H.H.; de Oliveira, D. Elucidating the choice for a precise matrix for laccase immobilization: A review. Chem. Eng. J. 2020, 397, 125506. [Google Scholar] [CrossRef]
- Huang, P.S.; Boyken, S.E.; Baker, D. The coming of age of de novo protein design. Nature 2016, 537, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Alvarado-Ramírez, L.; Rostro-Alanis, M.; Rodríguez-Rodríguez, J.; Castillo-Zacarías, C.; Sosa-Hernández, J.E.; Barceló, D.; Iqbal, H.M.N.; Parra-Saldívar, R. Exploring current tendencies in techniques and materials for immobilization of laccases–A review. Int J. Biol. Macromol. 2021, 181, 683–696. [Google Scholar] [CrossRef] [PubMed]
- Girelli, A.M.; Astolfi, M.L.; Scuto, F.R. Agro-industrial wastes as potential carriers for enzyme immobilization: A review. Chemosphere 2020, 244, 125368. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, A.M.; Tavares, A.P.; Rocha, C.M.; Cristóvão, R.O.; Teixeira, J.A.; Macedo, E.A. Immobilization of commercial laccase on spent grain. Process. Biochem. 2012, 47, 1095–1101. [Google Scholar] [CrossRef]
- Girelli, A.M.; Scuto, F.R. Spent grain as a sustainable and low-cost carrier for laccase immobilization. Waste Manag. 2021, 128, 114–121. [Google Scholar] [CrossRef]
- Ahmed, T.A.; Suso, H.P.; Hincke, M.T. In-depth comparative analysis of the chicken eggshell membrane proteome. J. Proteom. 2017, 155, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Girelli, A.M.; Scuto, F.R. Eggshell membrane as feedstock in enzyme immobilization. J. Biotechnol. 2021, 325, 241–249. [Google Scholar] [CrossRef]
- De Souza Bezerra, T.M.; Bassan, J.C.; de Oliveira Santos, V.T.; Ferraz, A.; Monti, R. Covalent immobilization of laccase in green coconut fiber and use in clarification of apple juice. Process. Biochem. 2015, 50, 417–423. [Google Scholar] [CrossRef]
- Cristóvão, R.O.; Silvério, S.C.; Tavares, A.P.M.; Jose, M.L.; Boaventura, R.A.L.; Macedo, E.A.; Coelho, M.A.Z. Green coconut fiber: A novel carrier for the immobilization of commercial laccase by covalent attachment for textile dyes decolourization. World J. Microbiol. Biotechnol. 2012, 28, 2827–2838. [Google Scholar] [CrossRef] [PubMed]
- Cristóvão, R.O.; Tavares, A.P.; Brígida, A.I.; Loureiro, J.M.; Boaventura, R.A.; Macedo, E.A.; Coelho, M.A.Z. Immobilization of commercial laccase onto green coconut fiber by adsorption and its application for reactive textile dyes degradation. J. Mol. Catal. B Enzym. 2011, 72, 6–12. [Google Scholar] [CrossRef]
- Li, N.; Xia, Q.; Niu, M.; Ping, Q.; Xiao, H. Immobilizing laccase on different species wood biochar to remove the chlorinated biphenyl in wastewater. Sci. Rep. 2018, 8, 13947. [Google Scholar] [CrossRef]
- Imam, A.; Suman, S.K.; Singh, R.; Vempatapu, B.P.; Ray, A.; Kanaujia, P.K. Application of laccase immobilized rice straw biochar for anthracene degradation. Environ. Pollut. 2021, 268, 115827. [Google Scholar] [CrossRef]
- Lonappan, L.; Liu, Y.; Rouissi, T.; Brar, S.K.; Verma, M.; Surampalli, R.Y. Adsorptive immobilization of agro-industrially produced crude laccase on various micro-biochars and degradation of diclofenac. Sci. Total Environ. 2018, 640, 1251–1258. [Google Scholar] [CrossRef] [PubMed]
- Tesfaye, T.; Sithole, B.; Ramjugernath, D. Valorisation of chicken feathers: Recycling and recovery routes. In Proceedings of the Sixteenth International Waste Management and Landfill Symposium S, Cagliari, Italy, 2–6 October 2017. [Google Scholar]
- Suman, S.K.; Patnam, P.L.; Ghosh, S.; Jain, S.L. Chicken feather derived novel support material for immobilization of laccase and its application in oxidation of veratryl alcohol. ACS Sustain. Chem. Eng. 2018, 7, 3464–3474. [Google Scholar] [CrossRef]
- Baig, K.S.; Wu, J.; Turcotte, G. Future prospects of delignification pretreatments for the lignocellulosic materials to produce second generation bioethanol. Int. J. Energy Res. 2019, 43, 1411–1427. [Google Scholar] [CrossRef]
- Shanmugam, S.; Hari, A.; Ulaganathan, P.; Yang, F.; Krishnaswamy, S.; Wu, Y.R. Potential of biohydrogen generation using the delignified lignocellulosic biomass by a newly identified thermostable laccase from Trichoderma asperellum strain BPLMBT1. Int. J. Hydrog. 2018, 43, 3618–3628. [Google Scholar] [CrossRef]
- Giacobbe, S.; Pezzella, C.; Lettera, V.; Sannia, G.; Piscitelli, A. Laccase pretreatment for agrofood wastes valorization. Bioresour. Technol. 2018, 265, 59–65. [Google Scholar] [CrossRef]
- Banerjee, R.; Chintagunta, A.D.; Ray, S. Laccase mediated delignification of pineapple leaf waste: An ecofriendly sustainable attempt towards valorization. BMC Chem. 2019, 13, 58. [Google Scholar] [CrossRef]
- Sherpa, K.C.; Ghangrekar, M.M.; Banerjee, R. A green and sustainable approach on statistical optimization of laccase mediated delignification of sugarcane tops for enhanced saccharification. J. Environ. Manag. 2018, 217, 700–709. [Google Scholar] [CrossRef]
- Rencoret, J.; Pereira, A.; del Río, J.C.; Martínez, A.T.; Gutiérrez, A. Delignification and saccharification enhancement of sugarcane byproducts by a laccase-based pretreatment. ACS Sustain. Chem. Eng. 2017, 5, 7145–7154. [Google Scholar] [CrossRef]
- Suman, S.K.; Malhotra, M.; Kurmi, A.K.; Narani, A.; Bhaskar, T.; Ghosh, S.; Iata Jain, S. Jute sticks biomass delignification through laccase-mediator system for enhanced saccharification and sustainable release of fermentable sugar. Chemosphere 2022, 286, 131687. [Google Scholar] [CrossRef] [PubMed]
- Rajak, R.C.; Banerjee, R. An eco-friendly process integration for second generation bioethanol production from laccase delignified Kans grass. Energy Conv. Manag. 2018, 157, 364–371. [Google Scholar] [CrossRef]
- Rajak, R.C.; Banerjee, R. Enzyme mediated biomass pretreatment and hydrolysis: A biotechnological venture towards bioethanol production. RSC Adv. 2016, 6, 61301–61311. [Google Scholar] [CrossRef]
- Lu, H.; Lou, H.; Hu, J.; Liu, Z.; Chen, Q. Macrofungi: A review of cultivation strategies, bioactivity, and application of mushrooms. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2333–2356. [Google Scholar] [CrossRef]
- Olivieri, G.; Wijffels, R.H.; Marzocchella, A.; Russo, M.E. Bioreactor and bioprocess design issues in enzymatic hydrolysis of lignocellulosic biomass. Catalysts 2021, 11, 680. [Google Scholar] [CrossRef]
- Branà, M.T.; Sergio, L.; Haidukowski, M.; Logrieco, A.F.; Altomare, C. Degradation of aflatoxin B1 by a sustainable enzymatic extract from spent mushroom substrate of Pleurotus eryngii. Toxins 2020, 12, 49. [Google Scholar] [CrossRef]
- Nazilah, K.R.; Koentjoro, M.P.; Isdiantoni, E.I.; Muslihatin, W.; Prasetyo, E.N. Effect of laccase oxidation pre-treatment on coffee (Coffea arabica) bean processing waste for composting substrate. In AIP Conference Proceedings; AIP Publishing LLC: Melville, NY, USA, 2020; Volume 2215, p. 070008. [Google Scholar] [CrossRef]
- Kuddus, M. Cold-active enzymes in food biotechnology: An updated mini review. J. Appl. Biol. Biotechnol. 2018, 6, 58–63. [Google Scholar] [CrossRef]
- Hamid, B.; Mohiddin, F.A. Cold-Active Enzymes in Food Processing. In Enzymes in Food Technology; Kuddus, M., Ed.; Springer Nature: Singapore, 2018. [Google Scholar] [CrossRef]
- Shi, Y.; Kong, D.; Liu, J.; Lu, J.; Yin, X.; Zhou, Q. Transformation of triclosan by a novel cold-adapted laccase from Botrytis sp. FQ. Front. Environ. Sci Eng. 2017, 11, 6. [Google Scholar] [CrossRef]
- Wisniewska, K.M.; Twarda-Clapa, A.; Białkowska, A.M. Screening of Novel Laccase Producers—Isolation and Characterization of Cold-Adapted Laccase from Kabatiella bupleuri G3 Capable of Synthetic Dye Decolorization. Biomolecules 2021, 11, 828. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.; Dou, X.; Huang, L.; Wang, L.; Meng, D.; Zhai, L.; Liao, X. Characterization of a robust cold-adapted and thermostable laccase from Pycnoporus sp. SYBC-L10 with a strong ability for the degradation of tetracycline and oxytetracycline by laccase-mediated oxidation. J. Hazard. Mater. 2020, 382, 121084. [Google Scholar] [CrossRef]
- Novoa, C.; Dhoke, G.V.; Mate, D.M.; Martínez, R.; Haarmann, T.; Schreiter, M.; Eidner, J.; Schwerdtfeger, R.; Lorents, P.; Davari, M.D.; et al. KnowVolution of a fungal laccase toward alkaline pH. ChemBioChem 2019, 20, 1458–1466. [Google Scholar] [CrossRef]
- Yin, Q.; Zhou, G.; Peng, C.; Zhang, Y.; Kües, U.; Liu, J.; Xiao, Y.; Fang, Z. The first fungal laccase with an alkaline pH optimum obtained by directed evolution and its application in indigo dye decolorization. AMB Express 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Xu, F. Effects of redox potential and hydroxide inhibition on the pH activity profile of fungal laccases. J. Biolog. Chem. 1997, 272, 924–928. [Google Scholar] [CrossRef]
- Hämäläinen, V.; Grönroos, T.; Suonpää, A.; Heikkilä, M.W.; Romein, B.; Ihalainen, P.; Malandra, R.; Birikh, K.R. Enzymatic processes to unlock the lignin value. Front. Bioeng. Biotechnol. 2018, 6, 20. [Google Scholar] [CrossRef]
- Du, W.; Zuo, D.; Gan, H.; Yi, C. Comparative study on the effects of laser bleaching and conventional bleaching on the physical properties of indigo kapok/cotton denim fabrics. Appl. Sci. 2019, 9, 4662. [Google Scholar] [CrossRef]
- Mate, D.M.; Gonzalez-Perez, D.; Falk, M.; Kittl, R.; Pita, M.; De Lacey, A.L.; Ludwig, R.; Shleev, S.; Alcalde, M. Blood tolerant laccase by directed evolution. Chem. Biol. 2013, 20, 223–231. [Google Scholar] [CrossRef]
- Stanzione, I.; Pezzella, C.; Giardina, P.; Sannia, G.; Piscitelli, A. Beyond natural laccases: Extension of their potential applications by protein engineering. Appl. Microb. Biotechnol. 2020, 104, 915–924. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SSF Substrate | Species | Substrate Composition and Growth Parameters | LC Activity | Reference |
|---|---|---|---|---|
| Brewers’ spent grains (BSG) | Trametes versicolor | Lignin 8.53%, cellulose 16.1%, hemicellulose 20%, ash 5.3% DW. Growth at 27 °C for 14 days | 560 U/L after 7 days | [108] |
| BSG added with LC inducers | T. versicolor | Lignin 12.4%, cellulose 13.8%, hemicellulose 30%, ash 2.6% DW. Growth at 30 °C for 16 days | 13,506 IU/g using 10 mg/kg phenol as inducer after 12 days | [109] |
| Wheat straw (WS), bean stalk (BS), and red grass (RG) | Lentinula edodes | WS: Lignin 7.58%, cellulose 68.93%, hemicellulose 11.16% DW. | 579 U/g DW after 25 days | [110] |
| BS: Lignin 11.27%, cellulose 64.65%, hemicellulose 11.27% DW. | 258 U/g DW after 25 days | |||
| RG: Lignin 7.5%, cellulose 69.22%, hemicellulose 10.69% DW. Growth at 26 °C for 40 days | 390 U/g DW after 35 days | |||
| Corn stalk (raw and steam exploded) | T. versicolor | Lignin 22.43%, cellulose 33.96%, hemicellulose 13.95% DW. Growth at 30 °C for 7 days | 2600 U/g after 15 days | [111] |
| Rice straw (raw and ammonia-treated) | Funalia trogii | - | 172 U/g after 14 days | [112] |
| Wheat and rice straw and bran | Tricholoma giganteum | - Growth at 30 °C for 20 days | 89,800 U/g after 16 days | [113] |
| Sorghum Bagasse | Coriolus versicolor | Lignin 25.14%, cellulose 38.02%, hemicellulose 25.01% DW. Growth at 27.5 °C for 20 days | 115 U/g after 20 days | [114] |
| Tea residues | T. versicolor | Lignin 13.60%, cellulose 11.60%, hemicellulose 32.50% DW. Growth at 26 °C for 8 days | 31.2 U/g after 8 days | [115] |
| Olive leaves and wheat straw | Galactomyces geotrichum | - Growth at 30 °C for 26 days | 56 U/g after 14 days | [116] |
| Carrier Material | LC Origin | Immobilization Method | Results | Application | Reference |
|---|---|---|---|---|---|
| Brewer’s spent grain | Commercial LC (DeniLite base) from Aspergillus spp. | Adsorption to acid/base treated spent grain | Recovered activity 99%, immobilization yield 95%, 75% activity retained after 10 cycles at pH 10 | - | [123] |
| Spent grains | Commercial LC (S igma-Aldrich) from T. versicolor | Imine binding with acid/base treated spent grain and adsorption to spent grain | Recovered activity 39%, immobilization yield 1.3%, 58% activity retained after 6 cycles | 58% removal of syringic acid after 6 cycles and complete removal in 4 h | [124] |
| Egg shell membrane | Commercial LC (S igma-Aldrich) from T. versicolor | Covalent bonding and adsorption | Recovered activity 57%, immobilization yield 41%, 40% activity retained after 6 cycles | 57% removal of syringic acid after 24 h | [126] |
| Green Coconut fibre (CF) | Commercial LC (DeniLite base) from Aspergillus spp. | Acid/oxidative pretreatment; Covalent attachment to functionalized CF | Immobilization yield 74%, 55% activity retained after 10 cycles | Up to 70% degradation of textile dyes. Operational activity is significantly reduced in following cycles. | [127] |
| CF | T. versicolor | CF was subjected to acid/base pretreatment associated with thermal decompression. Adsorption to functionalized CF; covalent bonding via glutaradehyde. | Recovered activity 59%, immobilization yield 98%, 16.5-fold increase in thermal stability, 80% activity retained after 10 cycles in operational conditions | 65% reduction of phenolic compounds from apple juice | [129] |
| Maple biochar | Coprinus comatus | Adsorption to maple biochar | Recovered activity 66.5%, immobilization yield 64%, 66% activity retained after 7 cycles. | 71.4% degradation of chlorinated biphenyls after 5 h of treatment (2.5 times higher than the free enzyme) | [130] |
| Rice straw biochar | T. maxima | Adsorption to acid-treated rice straw biochar | Recovered activity 66%, immobilization yield 100% at pH 3.1, 60% activity retained after 6 cycles | 98% degradation of anthracene after 24 h (comparable to the free enzyme) | [131] |
| Pine wood biochar | T. versicolor | Covalent immobilization via glutaraldehyde to citric acid pretreated biochars | Recovered activity 20.1%, 46% activity retained after 5 cycles | 98.9% removal of diclofenac after 5 h of treatment | [132] |
| Pig manure biochar | Recovered activity 40.2%, 40% activity retained after 5 cycles | 98.9% removal of diclofenac after 2 h of treatment | |||
| Almond shell biochar | Recovered activity 31.8%, 43% activity retained after 5 cycles | 98.9% removal of diclofenac after 4 h of treatment | |||
| Chicken feathers (Chf) | T. maxima | Covalent immobilization via glutaraldehyde to Chf functionalized with amino 3- aminopropyltrimethoxysilane | Recovered activity up to 93%, immobilization yield up to 74.24%, no significant loss of activity after 8 cycles | Complete oxidation of veratryl alcohol after 48 h | [134] |
| LC Origin | Application | Results | Reference |
|---|---|---|---|
| Trichoderma asperellum | Pretreatment of sweet sorghum stover for biohydrogen production | Removal of up to 77% of lignin; 3.26-fold increase in biohydrogen production. | [136] |
| Pleurotus ostreatus | Pretreatment of apple pomace, potato peels, and coffee silverskin for energy production | Up to 83% saccharification yields and ∼70% phenol reduction using 2.5% vanillin as redox mediator | [137] |
| P. djamor | Valorization of pineapple leaf waste for biofuel production | Removal of up to 78.57% of lignin, 2.6-fold increase in reducing sugars | [138] |
| P. djamor | Pretreatment of sugarcane tops | Decrease of up to 79.1% of lignin content; increase of 3.3 fold in fermentable sugars | [139] |
| Pycnoporus cinnabarinus | Valorization of sugarcane bagasse for energy production | Decrease of up to 27% of lignin content; increase of 39% of glucose release | [140] |
| Valorization of straw for energy production | Decrease of up to 31% of lignin content; increase of 46% of glucose release | ||
| Trametes maxima | Pretreatment of jute sticks for energy production | Decrease of up to 21.8% of lignin content; increase of 19.5% of glucose release using 5% HBT as redox mediator | [141] |
| Lentinus squarrosulus MR13 | Pretreatment of kans grass (Saccharum spontaneum) | Decrease of up to 87.8% of lignin content; increase of 9% of bioethanol production | [142] |
| Decrease of up to 81.2% of lignin content; production of up to 500 mg/g of fermentable sugars | [143] | ||
| T. versicolor | Pretreatment of coffee bean processing waste for composting | Increase in total plate count values | [144] |
| LC origin | Application | Results | Reference |
|---|---|---|---|
| Botrytis sp. FQ | Removal of antibiotics | 70% activity at temperatures between 0 °C and 30 °C and an optimal temperature of 15 °C on dimethoxyphenol and removal of up to 60% of triclosan at pH 6.8, 20 °C | [149] |
| Kabatiella bupleuri | Dye decolorization | Retention of 60% of the maximum activity at 10 °C and over 40% in ice; up to 48.6% degradation of crystal violet after 1-h reaction with ABTS | [150] |
| Lac-Q from Pycnoporus spp. | Removal of antibiotics | Retention of 51% of the maximum activity at in ice; degradation of 50 mg L−1 of oxytetracycline at pH 6.0 and 0 °C after 5 min with ABTS | [151] |
| Melanocarpus albomyces | Oxidation of dimethoxyphenol | Optimum pH of 9 for the substrate 2,6-dimethoxyphenol | [152] |
| Coprinopsis cinerea | Dye decolorization | High activity towards 2,6-dimethoxyphenol at pH 8-8.5; decolorization of 87% of indigo dye at pH 7.0–7.5 | [153] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loi, M.; Glazunova, O.; Fedorova, T.; Logrieco, A.F.; Mulè, G. Fungal Laccases: The Forefront of Enzymes for Sustainability. J. Fungi 2021, 7, 1048. https://doi.org/10.3390/jof7121048
Loi M, Glazunova O, Fedorova T, Logrieco AF, Mulè G. Fungal Laccases: The Forefront of Enzymes for Sustainability. Journal of Fungi. 2021; 7(12):1048. https://doi.org/10.3390/jof7121048
Chicago/Turabian StyleLoi, Martina, Olga Glazunova, Tatyana Fedorova, Antonio F. Logrieco, and Giuseppina Mulè. 2021. "Fungal Laccases: The Forefront of Enzymes for Sustainability" Journal of Fungi 7, no. 12: 1048. https://doi.org/10.3390/jof7121048
APA StyleLoi, M., Glazunova, O., Fedorova, T., Logrieco, A. F., & Mulè, G. (2021). Fungal Laccases: The Forefront of Enzymes for Sustainability. Journal of Fungi, 7(12), 1048. https://doi.org/10.3390/jof7121048