
Bioprocesses with Reduced Ecological Footprint by Marine Debaryomyces hansenii Strain for Potential Applications in Circular Economy



Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strain
2.2. Media Composition
2.3. Fermentation Conditions
2.4. Quantification of Biomass and Compounds
2.5. Phytase Sequence
2.6. Determination of Phytase Activity
3. Results and Discussion
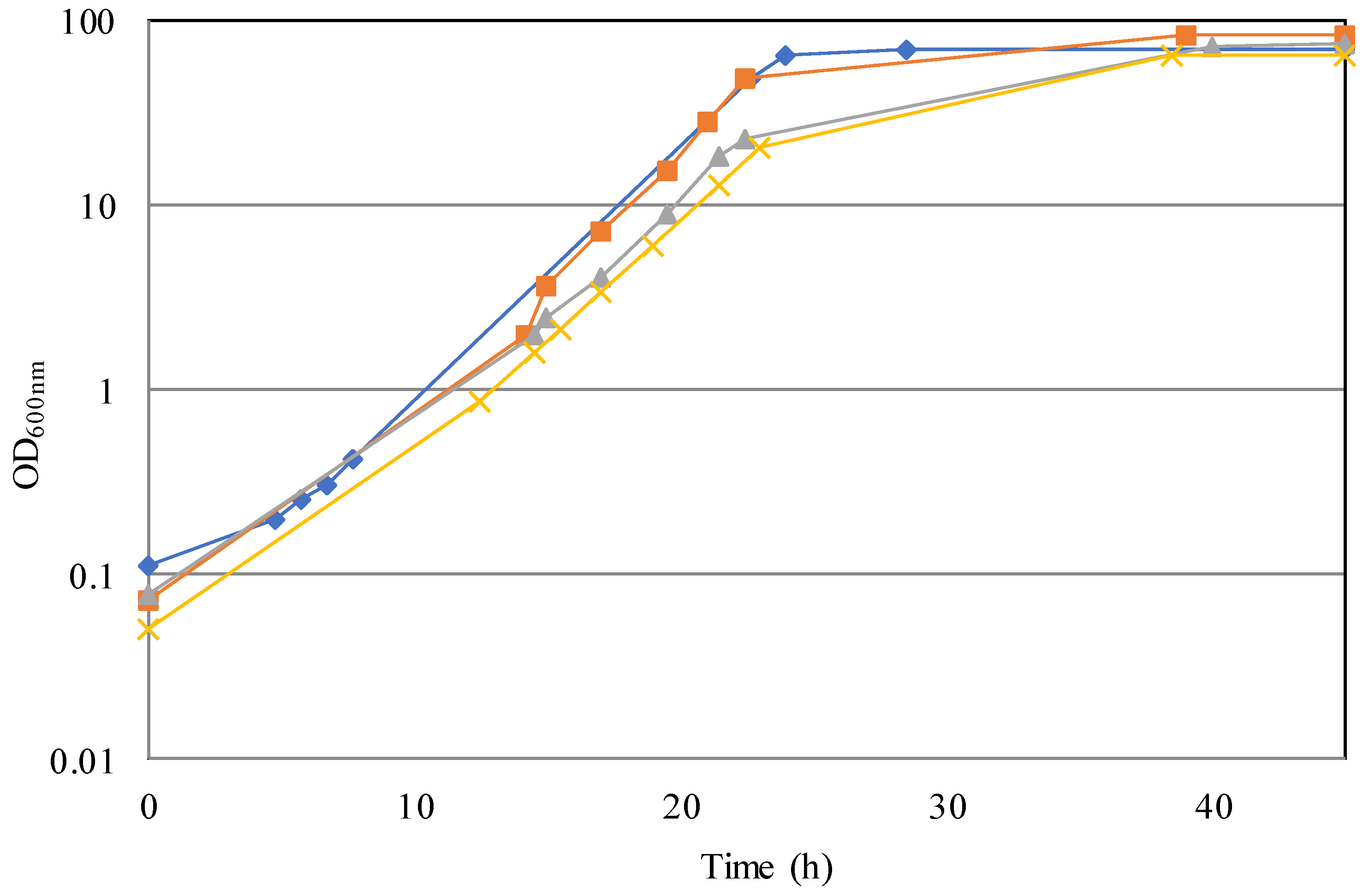
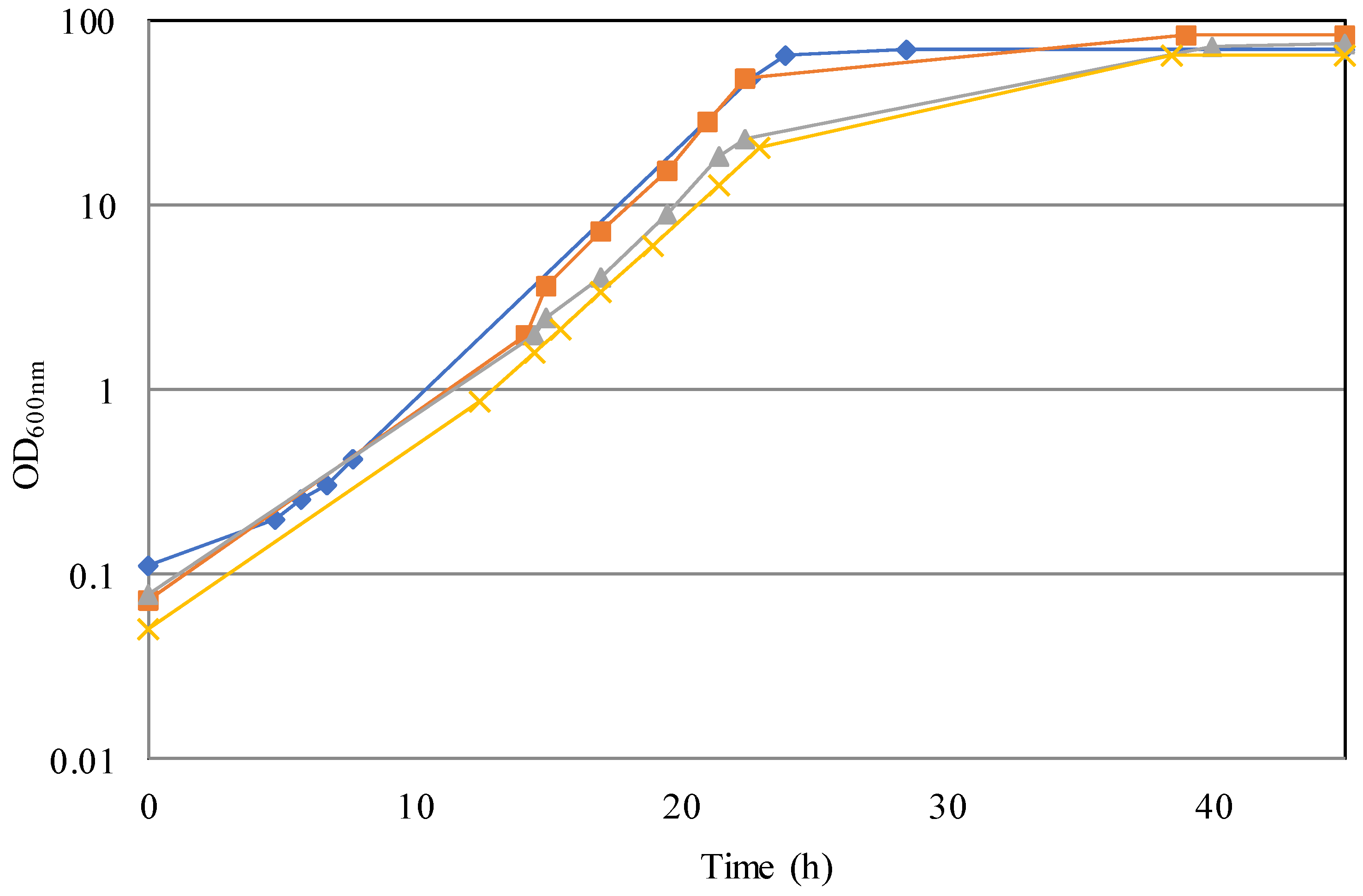
3.1. Cultivation in Bioreactor: Optimization of Growth Conditions
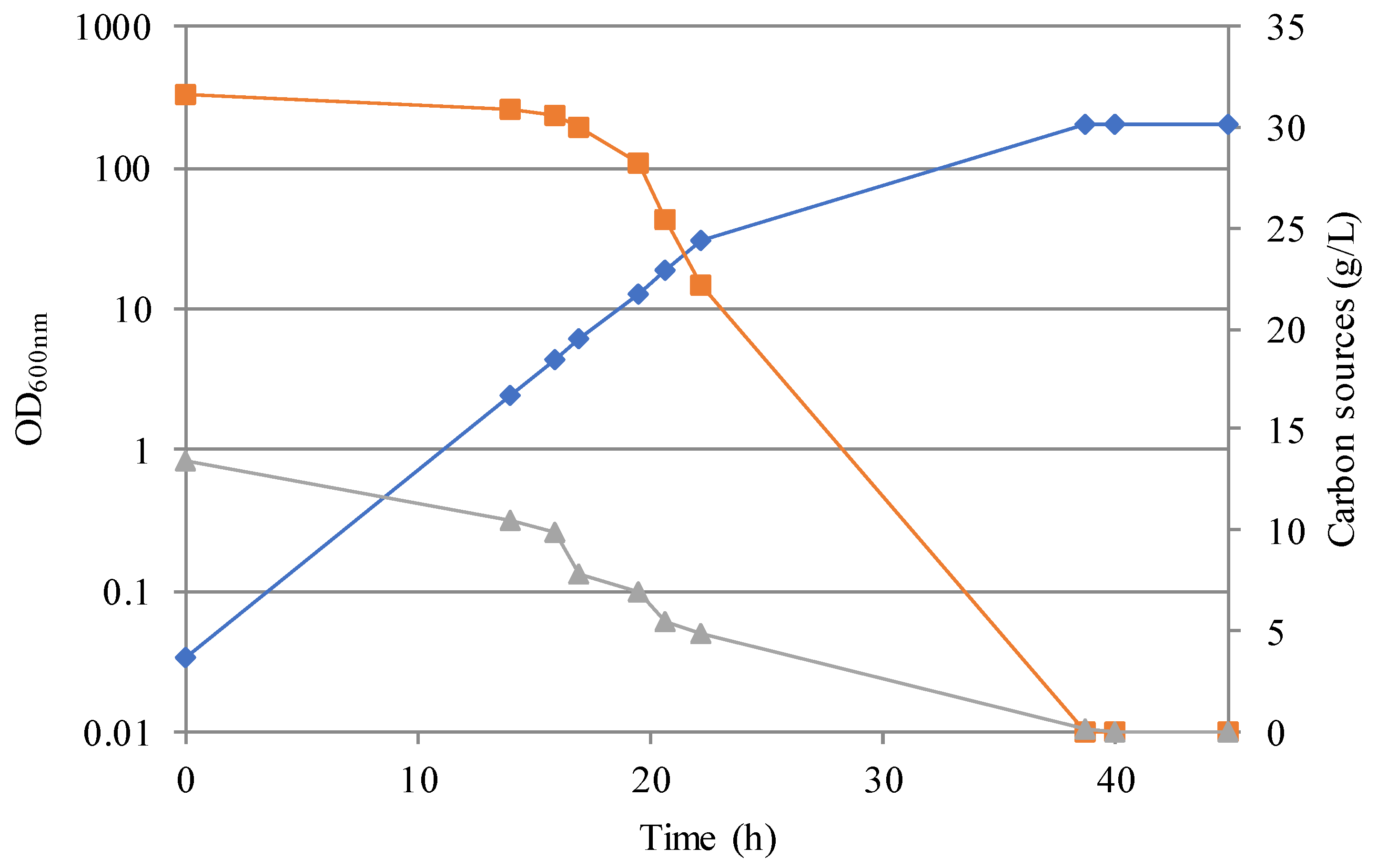
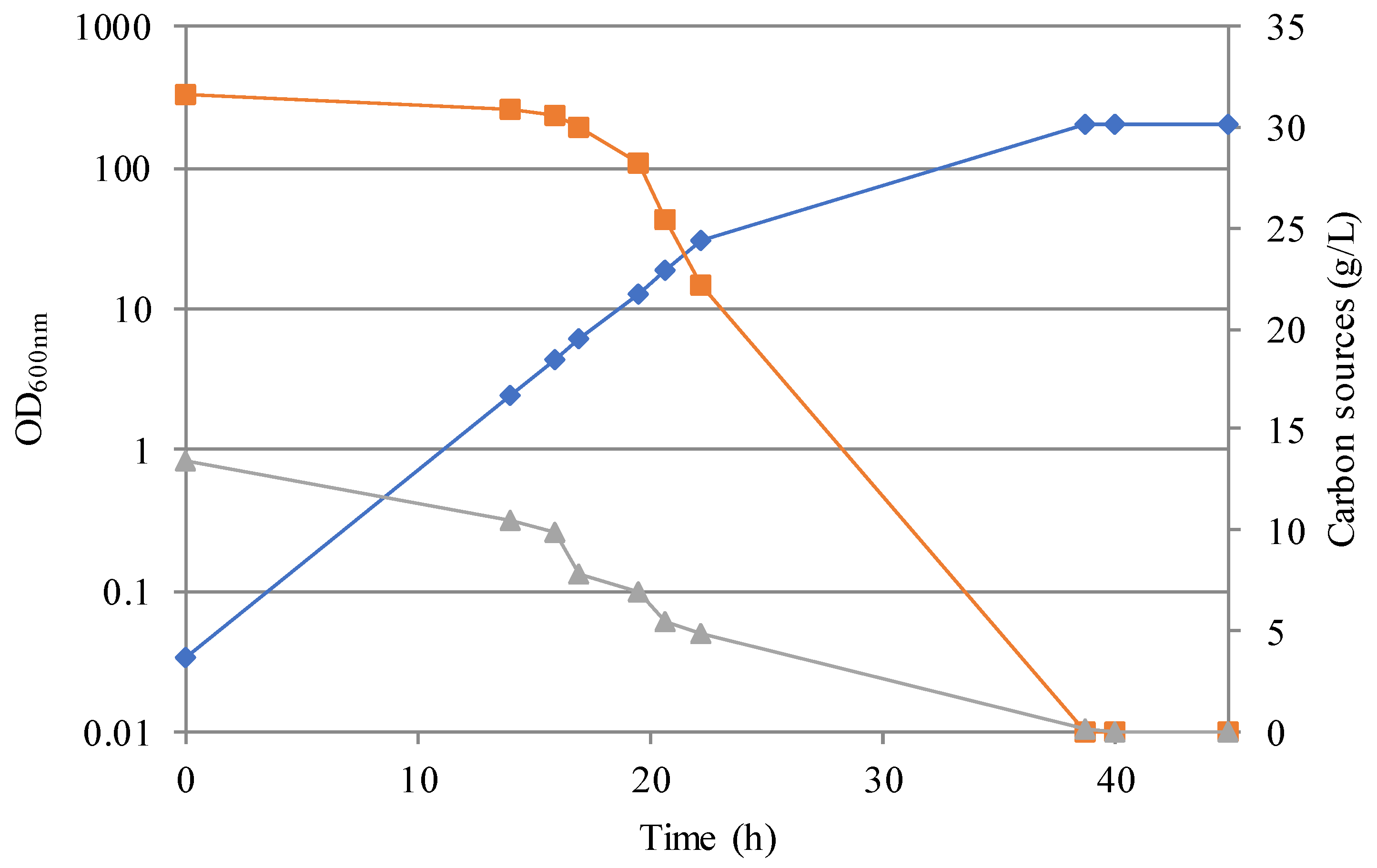
3.2. Simulation of an Industrial Bioprocess with Reduced Ecological Footprint
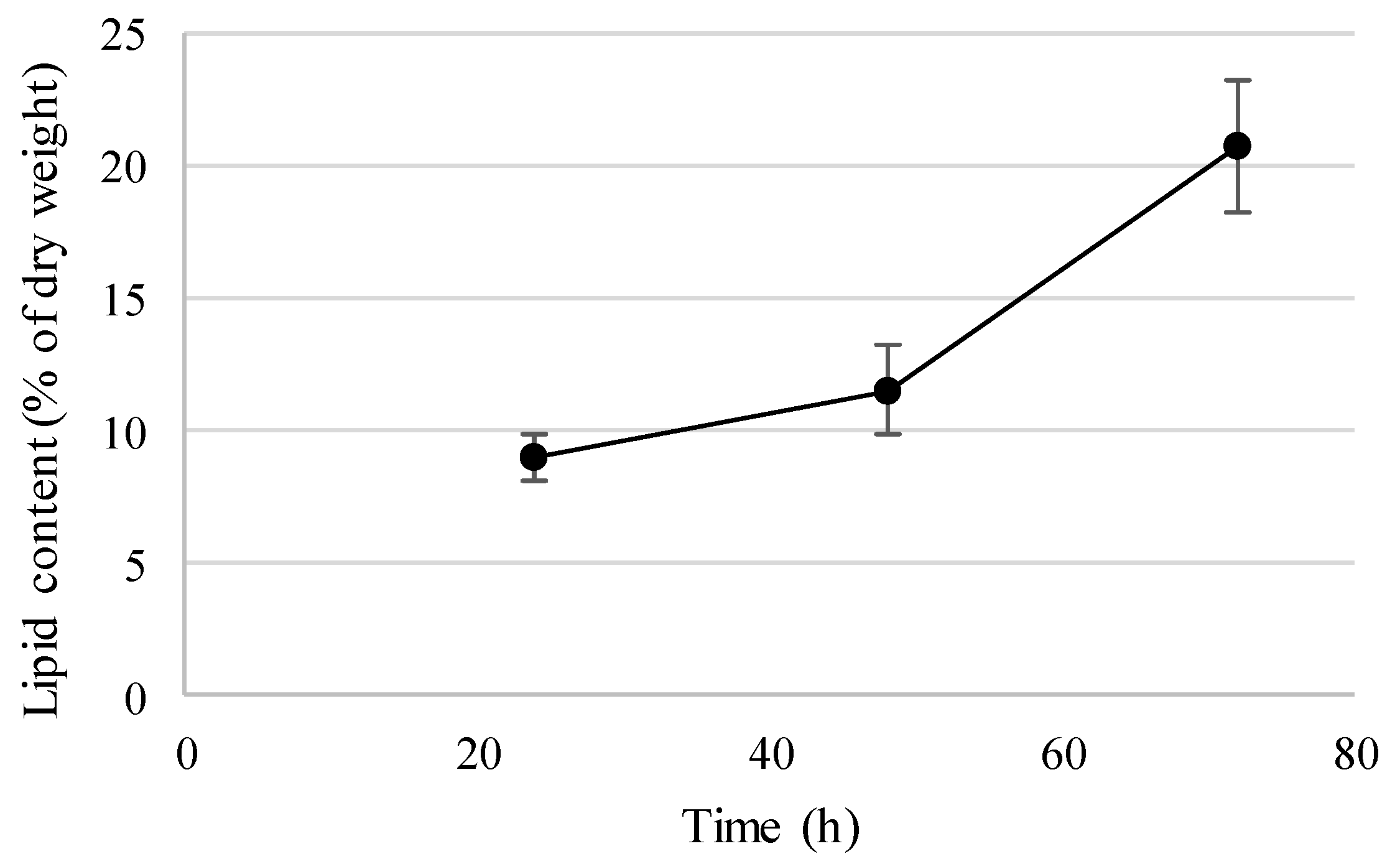
3.3. SCP and SCO Production
3.4. Phytase Activity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- D’Amato, D.; Droste, N.; Allen, B.; Kettunen, M.; Lähtinen, K.; Korhonen, J.; Leskinen, P.; Matthies, B.D.; Toppinen, A. Green, circular, bio economy: A comparative analysis of sustainability avenues. J. Clean. Prod. 2017, 168, 716–734. [Google Scholar] [CrossRef]
- Heer, D.; Sauer, U. Identification of furfural as a key toxin in lignocellulosic hydrolysates and evolution of a tolerant yeast strain. Microb. Biotechnol. 2008, 1, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Gerbens-Leenes, W.; Hoekstra, A.Y. The water footprint of sweeteners and bio-ethanol. Environ. Int. 2012, 40, 202–211. [Google Scholar] [CrossRef] [PubMed]
- Gerbens-Leenes, W.; Hoekstra, A.Y.; van der Meer, T.H. The water footprint of bioenergy. Proc. Natl. Acad. Sci. USA 2009, 106, 10219–10223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.S.; Zhang, F.; Miao, L.P.; Zhou, C. Novel Seawater Fermentation Strain Generating Biosurfactant. Patent CN104087525A, 24 August 2016. [Google Scholar]
- Fenical, W.; Jensen, P.; Mincer, T. Marine Actinomycete Taxon for Drug and Fermentation Product Discovery. Patent US20080070273, 1 February 2011. [Google Scholar]
- Trincone, A. Marine biocatalysts: Enzymatic features and applications. Mar. Drugs 2011, 9, 478–499. [Google Scholar] [CrossRef] [Green Version]
- Zaky, A.S.; Tucker, G.A.; Daw, Z.Y.; Du, C. Marine yeast isolation and industrial application. FEMS Yeast Res. 2014, 14, 813–825. [Google Scholar] [CrossRef]
- Andreu, C.; del Olmo, M. Biotransformation using halotolerant yeast in seawater: A sustainable strategy to produce R-(−)-phenylacetylcarbinol. Appl. Microbiol. Biotechnol. 2018, 102, 4717–4727. [Google Scholar] [CrossRef]
- Andreu, C.; del Olmo, M. Whole-cell biocatalysis in seawater: New halotolerant yeast strains for the regio- and stereoselectivity reduction of 1-phenylpropane-1,2-dione in saline-rich media. Chembiochem 2020, 21, 621–1628. [Google Scholar] [CrossRef]
- Prista, C.; Loureiro-Dias, M.C.; Montiel, V.; Garcia, R.; Ramos, J. Mechanisms underlying the halotolerant way of Debaryomyces hansenii. FEMS Yeast Res. 2005, 5, 693–701. [Google Scholar] [CrossRef] [Green Version]
- Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Hilbert, F.; Lindqvist, R.; et al. Update of the list of QPS-recommended biological agents intentionally added to food or feed as notified to EFSA 10: Suitability of taxonomic units notified to EFSA until March 2019. EFSA J. 2019, 17, e05753. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Moreno, L.; Ruiz-Pérez, F.; Rodríguez-Castro, E.; Ramos, J. Debaryomyces hansenii Is a Real Tool to Improve a Diversity of Characteristics in Sausages and Dry-Meat Products. Microorganisms 2021, 9, 1512. [Google Scholar] [CrossRef] [PubMed]
- Ochangco, H.S.; Gamero, A.; Smith, I.M.; Christensen, J.E.; Jespersen, L.; Arneborg, N. In vitro investigation of Debaryomyces hansenii strains for potential probiotic properties. World J. Microbiol. Biotechnol. 2016, 32, 141. [Google Scholar] [CrossRef] [PubMed]
- Iacumin, L.; Manzano, M.; Andyanto, D.; Comi, G. Biocontrol of ochratoxigenic moulds (Aspergillus ochraceus and Penicillium nordicum) by Debaryomyces hansenii and Saccharomycopsis fibuligera during speck production. Food Microbiol. 2017, 62, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Breuer, U.; Harms, H. Debaryomyces hansenii—An extremophilic yeast with biotechnological potential. Yeast 2006, 23, 415–437. [Google Scholar] [CrossRef]
- Gori, K.; Sorensen, L.M.; Petersen, M.A.; Jespersen, L.; Arneborg, N. Debaryomyces hansenii strains differ in their production of flavor compounds in a cheese-surface model. Microbiol. Open 2012, 1, 161–168. [Google Scholar] [CrossRef]
- Prista, C.; Michan, C.; Miranda, I.M.; Ramos, J. The halotolerant Debaryomyces hansenii, the cinderella of non-conventional yeasts. Yeast 2016, 33, 523–533. [Google Scholar] [CrossRef] [Green Version]
- Tavares, J.M.; Duarte, L.C.; Amaral-Collaço, M.T.; Gìrio, F.M. Phosphate limitation stress induces xylitol overproduction by Debaryomyces hansenii. FEMS Microbiol. Lett. 2016, 29, 1887–1891. [Google Scholar]
- Pal, S.; Choudhary, V.; Kumar, A.; Biswas, D.; Mondal, A.K.; Sahoo, D.K. Studies on xylitol production by metabolic pathway engineered Debaryomyces hansenii. Bioresour. Technol. 2013, 147, 449–455. [Google Scholar] [CrossRef]
- Capusoni, C.; Arioli, S.; Donzella, S.; Guidi, B.; Serra, I.; Compagno, C. Hyper-osmotic stress elicits membrane depolarization and decreased permeability in halotolerant marine Debaryomyces hansenii strains and in Saccharomyces cerevisiae. Front. Microbiol. 2019, 10, 64–70. [Google Scholar] [CrossRef] [Green Version]
- Burgaud, G.; Arzur, D.; Durand, L.; Cambon-Bonavita, M.A.; Barbier, G. Marine culturable yeasts in deep-sea hydrothermal vents: Species richness and association with fauna. FEMS Microbiol. Ecol. 2010, 73, 121–133. [Google Scholar] [CrossRef] [Green Version]
- Merico, A.; Sulo, P.; Piškur, J.; Compagno, C. Fermentative lifestyle in yeasts belonging to the Saccharomyces complex. FEBS J. 2007, 274, 976–989. [Google Scholar] [CrossRef]
- Lapeña, D.; Kosa, G.; Hansen, L.D.; Mydland, L.T.; Passoth, V.; Horn, S.J.; Eijsink, V.G.H. Production and characterization of yeasts grown on media composed of spruce-derived sugars and protein hydrolysates from chicken by-products. Microb. Cell Factories 2020, 19, 19. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Commission Regulation (EC) No 152/2009 of 27 January 2009 laying down the methods of sampling and analysis for the official control of feed. OJL 2009, 54, 1. [Google Scholar]
- Hogenboom, J.A.; D’Incecco, P.; Fuselli, F.; Pellegrino, L. Ion-exchange chromatographic method for the determination of the free amino acid composition of cheese and other dairy products: An inter-laboratory validation study. Food Anal. Methods 2017, 10, 3137–3148. [Google Scholar] [CrossRef] [Green Version]
- Donzella, S.; Cucchetti, D.; Capusoni, C.; Rizzi, A.; Galafassi, S.; Gambaro, C.; Compagno, C. Engineering cytoplasmic acetyl-CoA synthesis decouples lipid production from nitrogen starvation in the oleaginous yeast Rhodosporidium azoricum. Microb. Cell. Factories 2019, 18, 199. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M. Purification and characterization of Phytase from Bacillus suhtilis (natto) N–77. Biosci. Biotechnol. Biochem. 1992, 56, 1266–1269. [Google Scholar] [CrossRef]
- Sánchez, N.S.; Calahorra, M.; Ramírez, J.; Peña, A. Salinity and high pH affect energy pathways and growth in Debaryomyces hansenii. Fungal Biol. 2018, 122, 977–990. [Google Scholar] [CrossRef]
- Sukumaran, P.; Nulit, R.; Halimoon, N.; Simoh, S.; Omar, H.; Ismail, A. Formulation of cost-effective medium using urea as a nitrogen source for Arthrospira platensis cultivation under real environment. Annu. Res. Rev. Biol. 2018, 22, 1–12. [Google Scholar] [CrossRef]
- Øverland, M.; Karlsson, A.; Mydland, L.T.; Romarheim, O.H.; Skrede, A. Evaluation of Candida utilis, Kluyveromyces marxianus and Saccharomyces cerevisiae yeasts as protein sources in diets for Atlantic salmon (Salmon salar). Aquaculture 2013, 402–403, 1–7. [Google Scholar] [CrossRef]
- Øverland, M.; Skrede, A. Yeast derived from lignocellulosic biomass as a sustainable feed resource for use in aquaculture. J. Sci. Food Agric. 2017, 97, 733–742. [Google Scholar] [CrossRef]
- Koutinas, A.; Chatzifragkou, A.; Kopsahelis, N.; Papanikolaou, S.; Kookos, I. Design and techno-economic evaluation of microbial oil production as a renewable resource for biodiesel and oleochemical production. Fuel 2014, 116, 566–577. [Google Scholar] [CrossRef]
- Béligon, V.; Christophe, G.; Fontanille, P.; Larroche, C. Microbial lipids as potential source to food supplements. Curr. Opin. Food Sci. 2016, 7, 35–42. [Google Scholar] [CrossRef]
- Blomqvist, J.; Pickova, J.; Tilami, S.K.; Sampels, S.; Mikkelsen, N.; Brandenburg, J.; Sandgren, M.; Passoth, V. Oleaginous yeast as a component in fish feed. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [Green Version]
- Uemura, H. Synthesis and production of unsaturated and polyunsaturated fatty acids in yeast: Current state and perspectives. Appl. Microbiol. Biotechnol. 2012, 95, 1–12. [Google Scholar] [CrossRef]
- Chattopadhyay, A.; Mitra, M.; Maiti, M.K. Recent advances in lipid metabolic engineering of oleaginous yeasts. Biotechnol. Adv. 2021. [CrossRef]
- Upcraft, T.; Tu, W.; Johnson, R.; Finnigan, T.; Van Hung, N.; Hallett, J.; Guo, M. Protein from renewable resources: Mycoprotein production from agricultural residues. Green Chem. 2021, 23, 5150–5165. [Google Scholar] [CrossRef]
- Hellwig, C.; Gmoser, R.; Lundin, M.; Taherzadeh, M.J.; Rousta, K. Fungi Burger from Stale Bread? A Case Study on Perceptions. Of a Novel Protein-Rich Food Product Made from an Edible Fungus. Foods 2020, 9, 1112. [Google Scholar] [CrossRef]
- Andreu, C.; del Olmo, M. Improved biocatalytic activity of the Debaryomyces species in seawater. ChemCatChem 2019, 11, 3085. [Google Scholar] [CrossRef]
- Vasudevan, U.M.; Jaiswal, A.K.; Krishna, S.; Pandey, A. Thermostable phytase in feed and fuel industries. Bioresour. Technol. 2019, 278, 400–407. [Google Scholar] [CrossRef]
- Coban, H.B.; Demirci, A. Phytase as a diet ingredient: From microbial production to its 356 applications in food and feed industry. In Handbook of Food Bioengineering, Microbial Production of Food Ingredients and Additives; Holban, A.M., Grumezescu, A.M., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 33–55. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Nionelli, L.; Coda, R.; De Angelis, M.; Gobbetti, M. Effect of sourdough fermentation on stabilisation, and chemical and nutritional characteristics of wheat germ. Food Chem. 2010, 119, 1079–1089. [Google Scholar] [CrossRef]
- Leenhardt, F.; Levrat-Verny, M.A.; Chanliaud, E.; Rémésy, C. Moderate decrease of pH by sourdough fermentation is sufficient to reduce phytate content of whole wheat flour through endogenous phytase activity. J. Agric. Food Chem. 2005, 53, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Capusoni, C.; Serra, I.; Donzella, S.; Compagno, C. Screening for yeast Phytase leads to the identification of a new cell-bound and secreted activity in Cyberlindnera jadinii CJ2. Front. Bioeng. Biotechnol. 2021, 9, 662598. [Google Scholar] [CrossRef] [PubMed]
- Jacques, N.; Mallet, S.; Casaregola, S. Delimitation of the species of the Debaryomyces hansenii complex by intron sequence analysis. Int. J. Syst. Evol. Microbiol. 2009, 59, 1242–1251. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| µ max | Final Dry Weight | Biomass Yield | q Glucose | q Xylose | ||
|---|---|---|---|---|---|---|
| [h−1] | [g/L] | [gd.w./gc.s.] | [mmolglc/gdw/h] | [mmolxiyl/gdw/h] | ||
| pH 4.5 | MMP | 0.32 ± 0.010 | 13.5 ± 0.5 | 0.61 ± 0.016 | 2.83 ± 0.098 | - |
| MMPSS | 0.30 ± 0.008 | 12.3 ± 0.6 | 0.55 ± 0.015 | 2.47 ± 0.088 | - | |
| IMSS | 0.30 ± 0.011 | 28.5 ± 0.9 | 0.62 ± 0.018 | 2.56 ± 0.144 | 2.30 ± 0.100 | |
| pH 6 | MMP | 0.31 ± 0.012 | 13.8 ± 0.4 | 0.63 ± 0.012 | 2.57 ± 0.121 | - |
| MMPSS | 0.34 ± 0.013 | 13.4 ± 0.5 | 0.62 ± 0.015 | 3.25 ± 0.132 | - |
| mg/gDW | g/100 g Protein | mg/gDW (g/100 g Protein) Reported by Lapeña et al. [22] | |
|---|---|---|---|
| Asp | 20 | 10.2 | 37–48 (7–10) |
| Thr | 14 | 5.9 | 21–26 (4–5) |
| Ser | 13 | 6.4 | 21–28 (4–6) |
| Glu | 27 | 13.2 | 64–76 (13–15) |
| Gly | 9.6 | 4.8 | 19–30 (4–6) |
| Ala | 12 | 5.9 | 23–29 (5–6) |
| Val | 13 | 5.7 | 20–29 (4–6) |
| Cys a | 5 | 2.5 | 3–6 (0.6–1.2) |
| Met b | 2.5 | 1.1 | 4–7 (0.8–1.4) |
| Ile | 11 | 4.7 | 16–24 (3–5) |
| Leu | 18 | 8.1 | 29–38 (6–8) |
| Tyr | 6 | 3.0 | 11–18 (2–4) |
| Phe | 10 | 4.6 | 14–19 (3–4) |
| Lys | 16 | 7.2 | 27–43 (5–9) |
| His | 4.5 | 2.0 | 9–13 (2–3) |
| Arg | 10 | 4.4 | 21–32 (4–6) |
| Pro | 8 | 4.0 | 17–23 (3–5) |
| Total | 94.7 |
| Cell-Bound | Extracellular | |
|---|---|---|
| 60 °C | 5.03 ± 0.513 | BDL |
| 37 °C | 0.57 ± 0.071 | BDL |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Donzella, S.; Capusoni, C.; Pellegrino, L.; Compagno, C. Bioprocesses with Reduced Ecological Footprint by Marine Debaryomyces hansenii Strain for Potential Applications in Circular Economy. J. Fungi 2021, 7, 1028. https://doi.org/10.3390/jof7121028
Donzella S, Capusoni C, Pellegrino L, Compagno C. Bioprocesses with Reduced Ecological Footprint by Marine Debaryomyces hansenii Strain for Potential Applications in Circular Economy. Journal of Fungi. 2021; 7(12):1028. https://doi.org/10.3390/jof7121028
Chicago/Turabian StyleDonzella, Silvia, Claudia Capusoni, Luisa Pellegrino, and Concetta Compagno. 2021. "Bioprocesses with Reduced Ecological Footprint by Marine Debaryomyces hansenii Strain for Potential Applications in Circular Economy" Journal of Fungi 7, no. 12: 1028. https://doi.org/10.3390/jof7121028
APA StyleDonzella, S., Capusoni, C., Pellegrino, L., & Compagno, C. (2021). Bioprocesses with Reduced Ecological Footprint by Marine Debaryomyces hansenii Strain for Potential Applications in Circular Economy. Journal of Fungi, 7(12), 1028. https://doi.org/10.3390/jof7121028