
Genome-Wide Insight into Profound Effect of Carbon Catabolite Repressor (Cre1) on the Insect-Pathogenic Lifecycle of Beauveriabassiana




Abstract
:1. Introduction
2. Materials and Methods
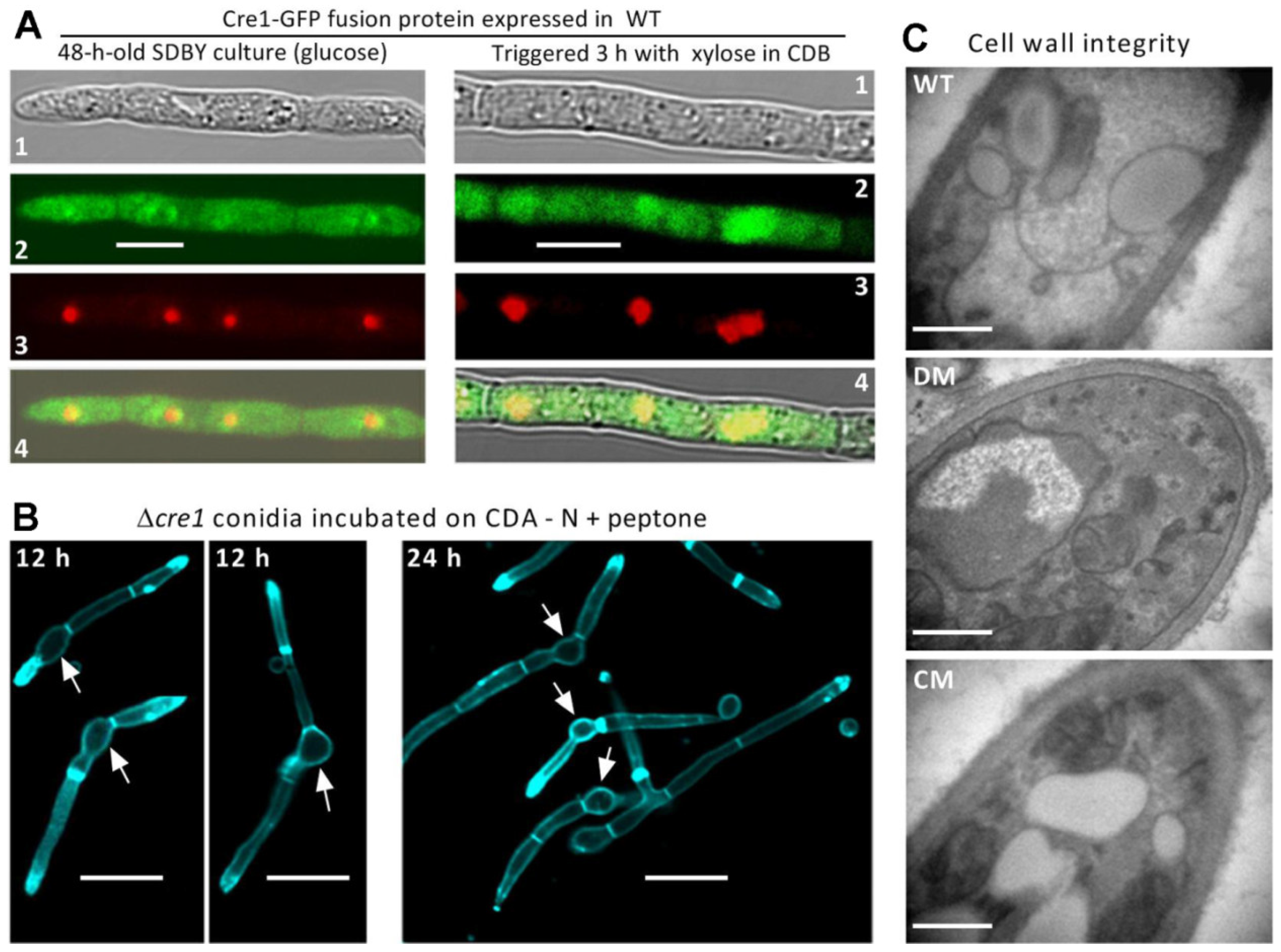
2.1. Subcellular Localization of Cre1
2.2. Generation of Cre1 Mutants
2.3. Examination of Cell-Lytic Phenotype
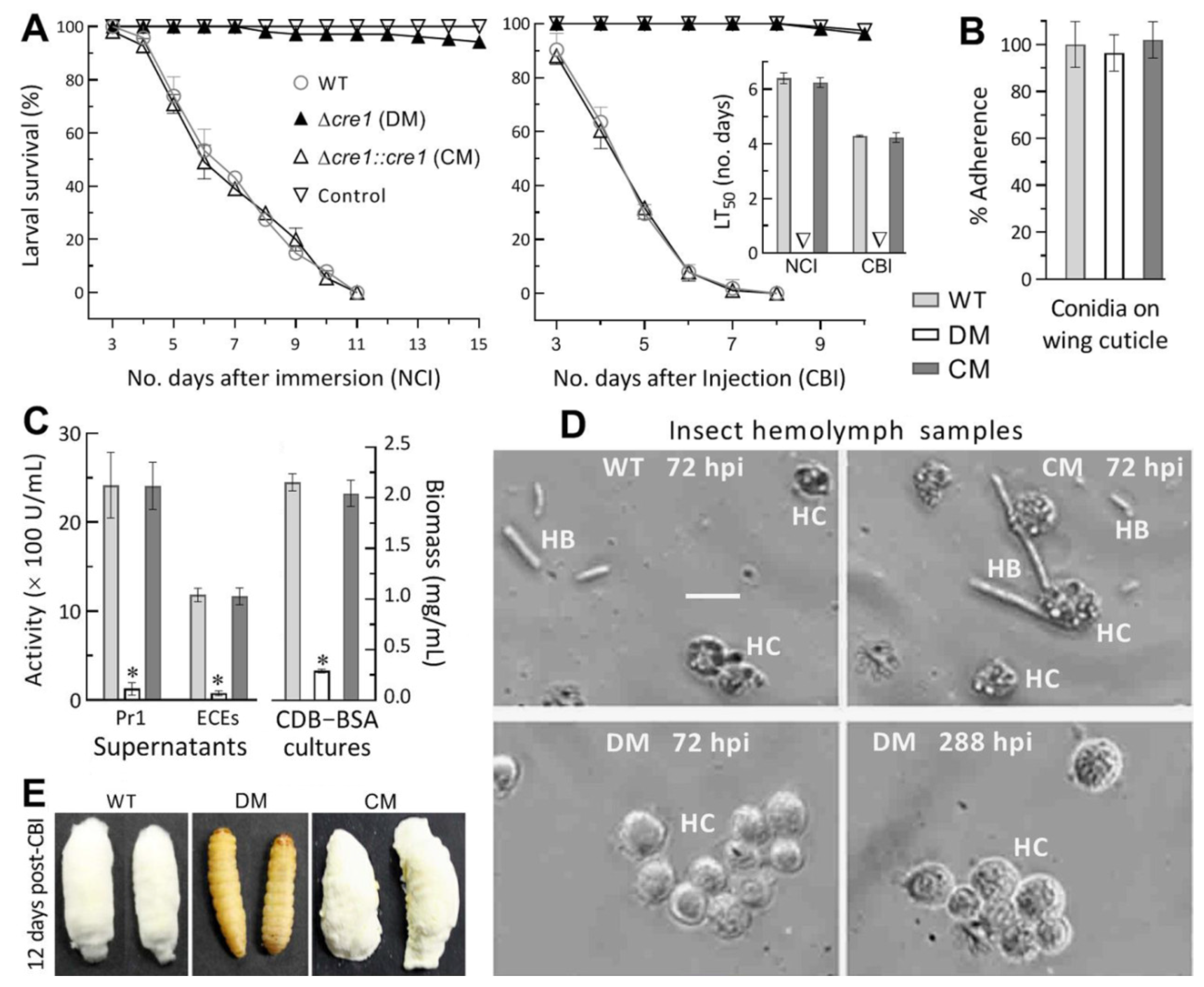
2.4. Assays for Virulence and Virulence-Related Cellular Events
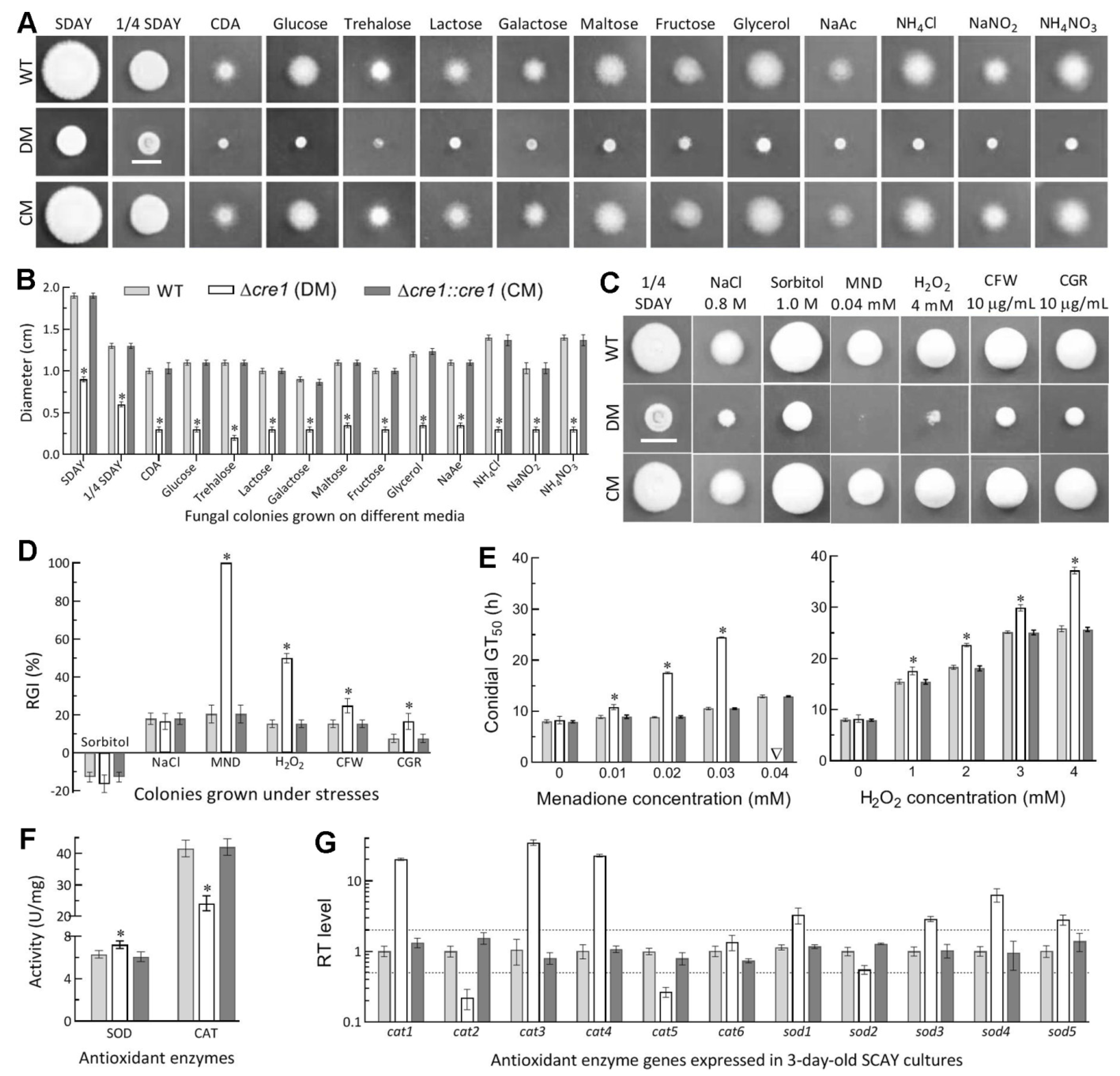
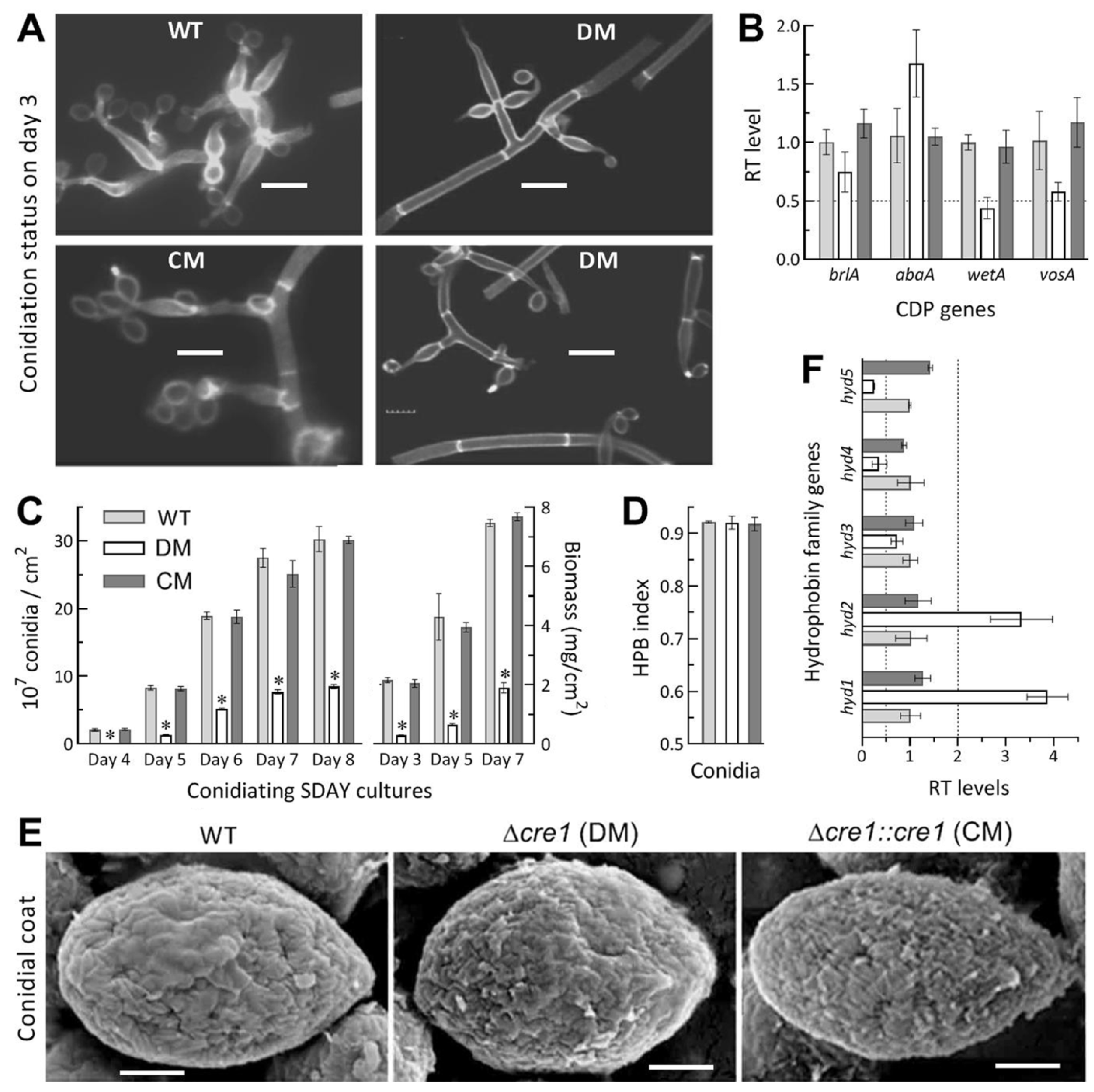
2.5. Assays for Radial Growth, Stress Response, Conidiation Capacity and Conidial Quality
2.6. Transcriptomic Analysis
2.7. Real-Time Quantitative PCR (qPCR) Analysis
2.8. Statistical Analysis
3. Results
3.1. Subcellular Localization of Cre1
3.2. No Cell-Lytic Phenotype Occurs in the Absence of Cre1
3.3. Indispensability of Cre1 for NCI and Hemocoel Colonization
3.4. Essential Role of Cre1 in Radial Growth and Antioxidant Response
3.5. Impact of Cre1 Deletion on Aerial Conidiation and Conidial Hydrophobicity
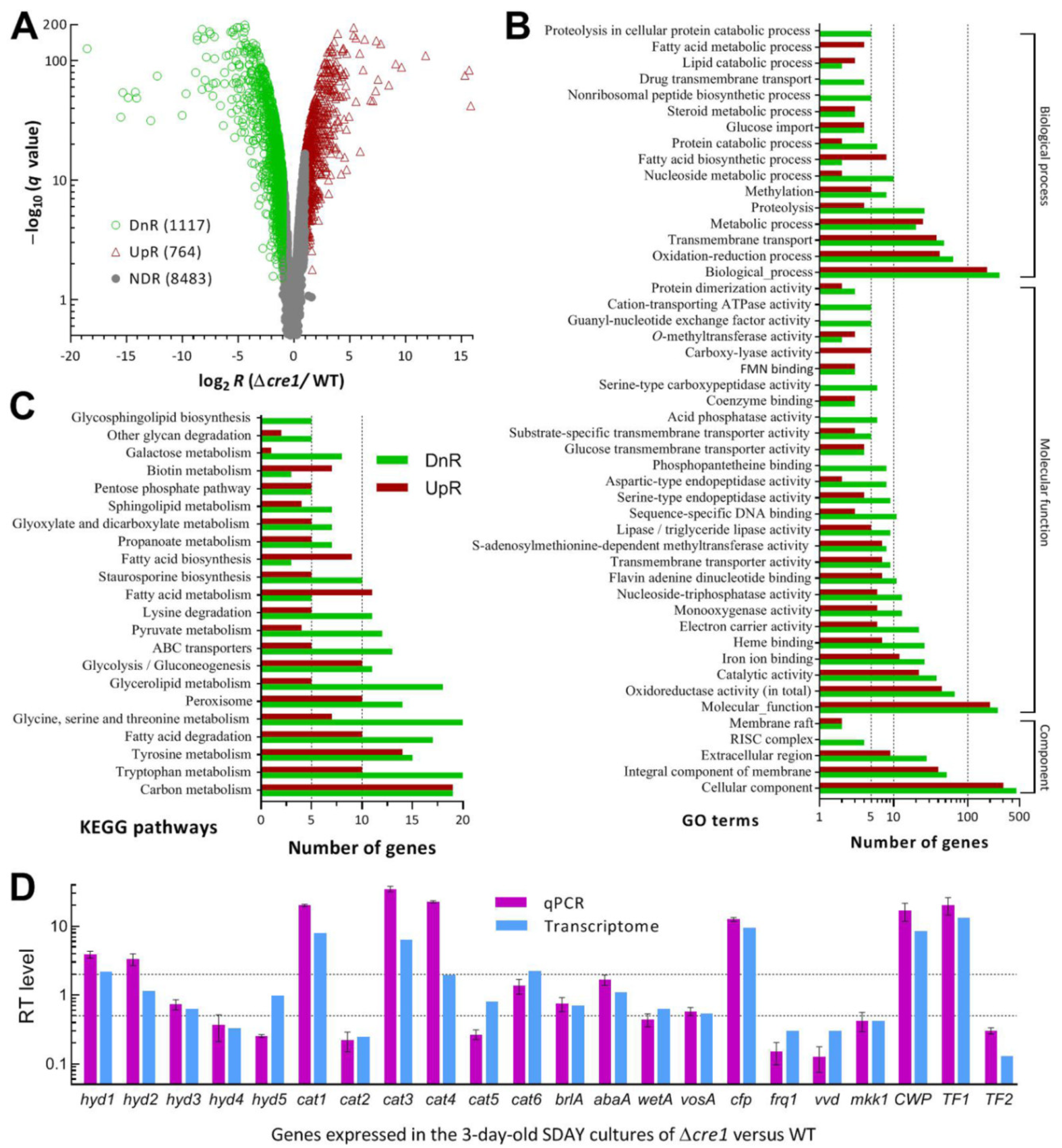
3.6. Genome-Wide Insight into Pleiotropic Effect of Cre1
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nehlin, J.O.; Ronne, H. Yeast MIG1 repressor is related to the mammalian early growth response and Wilms’ tumour finger proteins. EMBO J. 1990, 9, 2891–2898. [Google Scholar] [CrossRef] [PubMed]
- Lutfiyya, L.L.; Iyer, V.R.; DeRisi, J.; DeVit, M.J.; Brown, P.O.; Johnston, M. Characterization of three related glucose repressors and genes they regulate in Saccharomyces cerevisiae. Genetics 1998, 150, 1377–1391. [Google Scholar] [CrossRef]
- Westholm, J.O.; Nordberg, N.; Muren, E.; Ameur, A.; Komorowski, J.; Ronne, H. Combinatorial control of gene expression by the three yeast repressors Mig1, Mig2 and Mig3. BMC Genom. 2008, 9, 601. [Google Scholar] [CrossRef] [Green Version]
- Santangelo, G.M. Glucose signaling in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2006, 70, 253–282. [Google Scholar] [CrossRef] [Green Version]
- McCartney, R.R.; Schmidt, M.C. Regulation of Snf1 Kinase. Activation requires phosphorylation of threonine 210 by an upstream kinase as well as a distinct step mediated by the Snf4 subunit. J. Biol. Chem. 2001, 276, 36460–36466. [Google Scholar] [CrossRef] [Green Version]
- Hedbacker, K.; Carlson, M. SNF1/AMPK pathways in yeast. Front. Biosci. 2008, 13, 2408. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, G.W.; Welkenhuysen, N.; Ye, T.; Cvijovic, M.; Hohmann, S. Mig1 localization exhibits biphasic behavior which is controlled by both metabolic and regulatory roles of the sugar kinases. Mol. Genet. Genom. 2020, 295, 1489–1500. [Google Scholar] [CrossRef]
- Vassiliadis, D.; Wong, K.H.; Andrianopoulos, A.; Monahan, B.J. A genome-wide analysis of carbon catabolite repression in Schizosaccharomyces pombe. BMC Genom. 2019, 20, 251. [Google Scholar] [CrossRef] [PubMed]
- Dowzer, C.E.; Kelly, J.M. Analysis of the creA gene, a regulator of carbon catabolite repression in Aspergillus nidulans. Mol. Cell. Biol. 1991, 11, 5701–5709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scazzocchio, C.; Gavrias, V.; Cubero, B.; Panozzo, C.; Mathieu, M.; Felenbok, B. Carbon catabolite repression in Aspergillus nidulans—A review. Can. J. Bot.-Rev. Can. Bot. 1995, 73, S160–S166. [Google Scholar] [CrossRef]
- Panozzo, C.; Cornillot, E.; Felenbok, B. The CreA repressor is the sole DNA-binding protein responsible for carbon catabolite repression of the alcAgene in Aspergillus nidulans via its binding to a couple of specific sites. J. Biol. Chem. 1998, 273, 6367–6372. [Google Scholar] [CrossRef] [Green Version]
- Strauss, J.; Horvath, H.K.; Abdallah, B.M.; Kindermann, J.; Mach, R.L.; Kubicek, C.P. The function of CreA, the carbon catabolite repressor of Aspergillus nidulans, is regulated at the transcriptional and post-transcriptional level. Mol. Microbiol. 1999, 32, 169–178. [Google Scholar] [CrossRef]
- Mogensen, J.; Nielsen, H.B.; Hofmann, G.; Nielsen, J. Transcription analysis using high-density microarrays of Aspergillus nidulans wild-type and creA mutant during growth on glucose or ethanol. Fungal Genet. Biol. 2006, 43, 593–603. [Google Scholar] [CrossRef]
- Roy, P.; Lockington, R.A.; Kelly, J.M. CreA-mediated repression in Aspergillus nidulans does not require transcriptional auto- regulation, regulated intracellular localisation or degradation of CreA. Fungal Genet. Biol. 2008, 45, 657–670. [Google Scholar] [CrossRef]
- Ries, L.N.A.; Beattie, S.R.; Espeso, E.A.; Cramer, R.A.; Goldman, G.H. Diverse regulation of the CreA carbon catabolite repressor in Aspergillus nidulans. Genetics 2016, 203, 335–352. [Google Scholar] [CrossRef] [Green Version]
- De Assis, L.J.; Ulas, M.; Ries, L.N.A.; El Ramli, N.A.M.; Sarikaya-Bayram, O.; Braus, G.H.; Bayram, O.; Goldman, G.H. Regulation of Aspergillus nidulans CreA-mediated catabolite repression by the F-Box proteins Fbx23 and Fbx47. mBio 2018, 9, e00840-18. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.A.; Kamlangdee, N.; Kelly, J.M. The CreB deubiquitinating enzyme does not directly target the CreA repressor protein in Aspergillus nidulans. Curr. Genet. 2016, 63, 647–667. [Google Scholar] [CrossRef]
- Ribeiro, L.F.C.; Chelius, C.; Boppidi, K.R.; Naik, N.S.; Hossain, S.; Ramsey, J.J.J.; Kumar, J.; Ribeiro, L.F.; Ostermeier, M.; Tran, B.; et al. Comprehensive analysis of Aspergillus nidulans PKA phosphorylome identifies a novel mode of CreA regulation. mBio 2019, 10, e02825-18. [Google Scholar] [CrossRef] [Green Version]
- De Assis, L.J.; Silva, L.P.; Bayram, O.; Dowling, P.; Kniemeyer, O.; Kruger, T.; Brakhage, A.A.; Chen, Y.Y.; Dong, L.G.; Tan, K.L.; et al. Carbon catabolite repression in filamentous fungi is regulated by phosphorylation of the transcription factor CreA. mBio 2021, 12, e03146-20. [Google Scholar] [CrossRef]
- De Assis, L.J.; Silva, L.P.; Bayram, O.; Liu, L.; Schmitt, K.; Valerius, O.; Braus, G.H.; Ries, L.N.A.; Godman, G.H. The high osmolarity glycerol mitogen-activated protein kinase regulates glucose catabolite repression in filamentous fungi. PLoS Genet. 2020, 16, e1008996. [Google Scholar] [CrossRef]
- Mach, R.L.; Strauss, J.; Zeilinger, S.; Schindler, M.; Kubicek, C.P. Carbon catabolite repression of xylanase I (xyn1) gene expression in Trichoderma reesei. Mol. Microbiol. 1996, 21, 1273–1281. [Google Scholar] [CrossRef] [PubMed]
- Takashima, S.; Iikura, H.; Nakamura, A.; Masaki, H.; Uozumi, T. Analysis of Cre1 binding sites in the Trichoderma reesei cbh1 upstream region. FEMS Microbiol. Lett. 1996, 145, 361–366. [Google Scholar] [CrossRef]
- Cziferszky, A.; Mach, R.L.; Kubicek, C.P. Phosphorylation positively regulates DNA binding of the carbon catabolite repressor Cre1 of Hypocrea jecorina (Trichoderma reesei). J. Biol. Chem. 2002, 277, 14688–14694. [Google Scholar] [CrossRef] [Green Version]
- Portnoy, T.; Margeot, A.; Linke, R.; Atanasova, L.; Fekete, E.; Sandor, E.; Hartl, L.; Karaffa, L.; Druzhinina, I.S.; Seiboth, B.; et al. The CRE1 carbon catabolite repressor of the fungus Trichoderma reesei: A master regulator of carbon assimilation. BMC Genom. 2011, 12, 269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichius, A.; Seidl-Seiboth, V.; Seiboth, B.; Kubicek, C.P. Nucleo-cytoplasmic shuttling dynamics of the transcriptional regulators XYR1 and CRE1 under conditions of cellulase and xylanase gene expression in Trichoderma reesei. Mol. Microbiol. 2014, 94, 1162–1178. [Google Scholar] [CrossRef]
- Vautard, G.; Cotton, P.; Fevre, M. The glucose repressor CRE1 from Sclerotinia sclerotiorum is functionally related to CREA from Aspergillus nidulans but not to the Mig proteins from Saccharomyces cerevisiae. FEBS Lett. 1999, 453, 54–58. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.; Kim, K.H.; La Rota, M.; Scott, D.; Santopietro, G.; Callihan, M.; Mitchell, T.K.; Lawrence, C.B. Identification of novel virulence factors associated with signal transduction pathways in Alternaria brassicicola. Mol. Microbiol. 2009, 72, 1316–1333. [Google Scholar] [CrossRef]
- Jonkers, W.; Rep, M. Mutation of CRE1 in Fusarium oxysporum reverts the pathogenicity defects of the FRP1 deletion mutant. Mol. Microbiol. 2009, 74, 1100–1113. [Google Scholar] [CrossRef] [PubMed]
- Randhawa, A.; Ogunyewo, O.A.; Eqbal, D.; Gupta, M.; Yazdani, S.S. Disruption of zinc finger DNA binding domain in catabolite repressor Mig1 increases growth rate, hyphal branching, and cellulase expression in hypercellulolytic fungus Penicillium funiculosum NCIM1228. Biotechnol. Biofules 2018, 11, 15. [Google Scholar] [CrossRef]
- Xu, X.X.; Fan, C.; Song, L.Y.; Li, J.Y.; Chen, Y.; Zhang, Y.H.; Liu, B.; Zhang, W. A novel CreA-mediated regulation mechanism of cellulase expression in the thermophilic fungus Humicola insolens. Int. J. Mol. Sci. 2019, 20, 3693. [Google Scholar] [CrossRef] [Green Version]
- Tong, S.M.; Feng, M.G. Insights into regulatory roles of MAPK-cascaded pathways in multiple stress responses and life cycles of insect and nematode mycopathogens. Appl. Microbiol. Biotechnol. 2019, 103, 577–587. [Google Scholar] [CrossRef]
- Tong, S.M.; Feng, M.G. Phenotypic and molecular insights into heat tolerance of formulated cells as active ingredients of fungal insecticides. Appl. Microbiol. Biotechnol. 2020, 104, 5711–5724. [Google Scholar] [CrossRef]
- Tong, S.M.; Feng, M.G. Molecular basis and regulatory mechanisms underlying fungal insecticides’ resistance to solar ultraviolet irradiation. Pest Manag. Sci. 2021. [Google Scholar] [CrossRef]
- Screen, S.; Bailey, A.; Charnley, K.; Cooper, R.; Clarkson, J. Carbon regulation of the cuticledegrading enzyme PR1 from Metarhizium anisopliae may involve a trans-acting DNA binding protein CRR1, a functional equivalent of the Aspergillus nidulans CREA protein. Curr. Genet. 1997, 31, 511–518. [Google Scholar] [CrossRef]
- Jin, K.; Luo, Z.B.; Jiang, X.D.; Zhang, Y.J.; Zhou, Y.H.; Pei, Y. Carbon catabolite repressor gene BbCre1 influences carbon source uptake but does not have a big impact on virulence in Beauveriabassiana. J. Invertebr. Pathol. 2011, 106, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.J.; Mou, Y.N.; Tong, S.M.; Ying, S.H.; Feng, M.G. Subtilisin-like Pr1 proteases marking evolution of pathogenicity in a wide-spectrum insect-pathogenic fungus. Virulence 2020, 11, 365–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Z.B.; Qin, Y.Q.; Pei, Y.; Keyhani, N.O. Ablation of the creA regulator results in amino acid toxicity, temperature sensitivity, pleiotropic effects on cellular development and loss of virulence in the filamentous fungus Beauveria bassiana. Environ. Microbiol. 2014, 16, 1122–1136. [Google Scholar] [CrossRef]
- Lai, Y.L.; Cao, X.; Chen, J.J.; Wang, L.L.; Wei, G.; Wang, S.B. Coordinated regulation of infection-related morphogenesis by the KMT2-Cre1-Hyd4 regulatory pathway to facilitate fungal infection. Sci. Adv. 2020, 6, eaaz1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.Z.; Xia, Y.X.; Kim, B.; Keyhani, N.O. Two hydrophobins are involved in fungal spore coat rodlet layer assembly and each playdistinct roles in surface interactions, development and pathogenesis in the entomopathogenic fungus, Beauveria bassiana. Mol. Microbiol. 2011, 80, 811–826. [Google Scholar] [CrossRef] [PubMed]
- Ren, K.; Mou, Y.N.; Yu, L.; Tong, S.M.; Ying, S.H.; Feng, M.G. SET1/KMT2-governed histone H3K4 methylation coordinates the lifecycle in vivo and in vitro of the fungal insect pathogen Beauveria bassiana. Environ. Microbiol. 2021, 23, 5541–5554. [Google Scholar] [CrossRef]
- Zhou, Q.; Yu, L.; Ying, S.H.; Feng, M.G. Comparative roles of three adhesin genes (adh1–3) in insect-pathogenic lifecycle of Beauveria bassiana. Appl. Microbiol. Biotechnol. 2021, 105, 5491–5502. [Google Scholar] [CrossRef]
- Ortiz-Urquiza, A.; Keyhani, N.O. Action on the surface: Entomopathogenic fungi versus the insect cuticle. Insects 2013, 4, 357–374. [Google Scholar] [CrossRef]
- Pedrini, N.; Ortiz-Urquiza, A.; Huarte-Bonnet, C.; Zhang, S.Z.; Keyhani, N.O. Targeting of insect epicuticular lipids by the entomopathogenic fungus Beauveriabassiana: Hydrocarbon oxidation within the context of a host-pathogen interaction. Front. Microbiol. 2013, 4, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.Y.; Fu, B.; Tong, S.M.; Ying, S.H.; Feng, M.G. Two photolyases repair distinct DNA lesions and reactivate UVB-inactivated conidia of an insect mycopathogen under visible light. Appl. Environ. Microbiol. 2019, 85, e02459-18. [Google Scholar] [CrossRef] [Green Version]
- Mou, Y.N.; Fu, B.; Ren, K.; Ying, S.H.; Feng, M.G. A small cysteine-free protein acts as a novel regulator of fungal insect-pathogenic lifecycle and genomic expression. mSystems 2021, 6, e00098-21. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.; Ying, S.H.; Zheng, P.; Wang, Z.L.; Zhang, S.; Xie, X.Q.; Shang, Y.; Leger, R.J.S.; Zhao, G.P.; Wang, C.; et al. Genomic perspectives on the evolution of fungal entomopathogenicity in Beauveria bassiana. Sci. Rep. 2012, 2, 483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Ying, S.H.; Hu, Y.; Feng, M.G. Mas5, a homologue of bacterial DnaJ, is indispensable for the host infection and environmental adaptation of a filamentous fungal insect pathogen. Environ. Microbiol. 2016, 18, 1037–1047. [Google Scholar] [CrossRef] [Green Version]
- Ren, K.; Mou, Y.N.; Tong, S.M.; Ying, S.H.; Feng, M.G. DIM5/KMT1 controls fungal insect pathogenicity and genome stability by methylation of histone H3K4, H3K9 and H3K36. Virulence 2021, 12, 1306–1322. [Google Scholar] [CrossRef]
- Guo, C.T.; Peng, H.; Tong, S.M.; Ying, S.H.; Feng, M.G. Distinctive role of fluG in the adaptation of Beauveria bassiana to insect- pathogenic lifecycle and environmental stresses. Environ. Microbiol. 2021, 23, 5184–5199. [Google Scholar] [CrossRef]
- Shao, W.; Cai, Q.; Tong, S.M.; Ying, S.H.; Feng, M.G. Nuclear Ssr4 is required for the in vitro and in vivo asexual cycles and global gene activity of Beauveria bassiana. mSystems 2020, 5, e00677-19. [Google Scholar] [CrossRef] [Green Version]
- Mou, Y.N.; Ren, K.; Tong, S.M.; Ying, S.H.; Feng, M.G. Essential role of COP9 signalosome subunit 5 (Csn5) in insect pathogenicity and asexual development of Beauveria bassiana. J. Fungi 2021, 7, 642. [Google Scholar] [CrossRef]
- Zhang, L.B.; Feng, M.G. Antioxidant enzymes and their contributions to biological control potential of fungal insect pathogens. Appl. Microbiol. Biotechnol. 2018, 102, 4995–5004. [Google Scholar] [CrossRef]
- Shao, W.; Cai, Q.; Tong, S.M.; Ying, S.H.; Feng, M.G. Rei1-like protein regulates nutritional metabolism and transport required for the asexual cycle in vitro and in vivo of a fungal insect pathogen. Environ. Microbiol. 2019, 21, 2772–2786. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.L.; Zhang, L.B.; Ying, S.H.; Feng, M.G. Catalases play differentiated roles in the adaptation of a fungal entomopathogen to environmental stresses. Environ. Microbiol. 2013, 15, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.Q.; Li, F.; Ying, S.H.; Feng, M.G. Additive contributions of two manganese-cored superoxide dismutases (MnSODs) to anti-oxidation, UV tolerance and virulence of Beauveria bassiana. PLoS ONE 2012, 7, e30298. [Google Scholar]
- Zhang, A.X.; Mouhoumed, A.Z.; Tong, S.M.; Ying, S.H.; Feng, M.G. BrlA and AbaA govern virulence-required dimorphic switch, conidiation and pathogenicity in a fungal insect pathogen. mSystems 2019, 4, e00140-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Shi, H.Q.; Ying, S.H.; Feng, M.G. WetA and VosA are distinct regulators of conidiation capacity, conidial quality, and biological control potential of a fungal insect pathogen. Appl. Microbiol. Biotechnol. 2015, 99, 10069–10081. [Google Scholar] [CrossRef]
- Xu, Y.Q.; Rozco, R.; Wijeratne, E.M.K.; Espinosa-Artiles, P.; Gunatilaka, A.A.L.; Stock, S.P.; Stock, S.; Molnar, I. Biosynthesis of the cyclooligomer depsipeptide bassianolide, an insecticidal virulence factor of Beauveriabassiana. Fungal Genet. Biol. 2009, 46, 353–364. [Google Scholar] [CrossRef]
- Tong, S.M.; Zhang, A.X.; Guo, C.T.; Ying, S.H.; Feng, M.G. Daylight length-dependent translocation of VIVID photoreceptor in cells and its essential role in conidiation and virulence of Beauveria bassiana. Environ.Microbiol. 2018, 20, 169–185. [Google Scholar] [CrossRef]
- Tong, S.M.; Wang, D.Y.; Cai, Q.; Ying, S.H.; Feng, M.G. Opposite nuclear dynamics of two FRH-dominated frequency proteins orchestrate non-rhythmic conidiation of Beauveria bassiana. Cells 2020, 9, 626. [Google Scholar] [CrossRef] [Green Version]
- Ding, J.L.; Hou, J.; Feng, M.G.; Ying, S.H. Transcriptomic analyses reveal comprehensive responses of insect hemocytes to mycopathogen Beauveria bassiana, and fungal virulence-related cell wall protein assists pathogen to evade host cellular defense. Virulence 2020, 11, 1352–1365. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.H.; Cai, R.L.; Guo, J.Y.; Zhong, Z.H.; Bao, J.D.; Wang, Z.H.; Chen, X.F.; Zhou, J.; Lu, G.D. Carbon catabolite repressor MoCreA is required for the asexual development and pathogenicity of the rice blast fungus. Fungal Genet. Biol. 2021, 146, 103496. [Google Scholar] [CrossRef] [PubMed]
- Beattie, S.R.; Mark, K.M.K.; Thammahong, A.; Ries, L.N.A.; Dhingra, S.; Caffrey-Carr, A.K.; Cheng, C.; Black, C.C.; Bowyer, P.; Bromley, M.J.; et al. Filamentous fungal carbon catabolite repression supports metabolic plasticity and stress responses essential for disease progression. PLoS Pathog. 2017, 13, e1006340. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cellular Processes and Events | Number of Genes | ||
|---|---|---|---|
| Down | Up | Total | |
| Carbon/nitrogen metabolisms and energy conversion | 103 | 63 | 166 |
| Host infection- and virulence-related events | 31 | 15 | 46 |
| Asexual development (aerial conidiation) | 5 | 1 | 6 |
| Response to oxidative stress | 22 | 16 | 38 |
| Cell wall composition/integrity | 11 | 20 | 31 |
| Thermal tolerance (response to heat) | 7 | 0 | 7 |
| Cellular transport and homeostasis | 103 | 55 | 158 |
| Transcriptional regulation (transcription factors) | 52 | 23 | 75 |
| Postranslational modifications and chromatin remodeling | 31 | 18 | 49 |
| DNA splicing, repair, reverse transcription and translation | 13 | 6 | 19 |
| Cellular signaling | 22 | 9 | 31 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohamed, R.A.; Ren, K.; Mou, Y.-N.; Ying, S.-H.; Feng, M.-G. Genome-Wide Insight into Profound Effect of Carbon Catabolite Repressor (Cre1) on the Insect-Pathogenic Lifecycle of Beauveriabassiana. J. Fungi 2021, 7, 895. https://doi.org/10.3390/jof7110895
Mohamed RA, Ren K, Mou Y-N, Ying S-H, Feng M-G. Genome-Wide Insight into Profound Effect of Carbon Catabolite Repressor (Cre1) on the Insect-Pathogenic Lifecycle of Beauveriabassiana. Journal of Fungi. 2021; 7(11):895. https://doi.org/10.3390/jof7110895
Chicago/Turabian StyleMohamed, Rehab Abdelmonem, Kang Ren, Ya-Ni Mou, Sheng-Hua Ying, and Ming-Guang Feng. 2021. "Genome-Wide Insight into Profound Effect of Carbon Catabolite Repressor (Cre1) on the Insect-Pathogenic Lifecycle of Beauveriabassiana" Journal of Fungi 7, no. 11: 895. https://doi.org/10.3390/jof7110895
APA StyleMohamed, R. A., Ren, K., Mou, Y.-N., Ying, S.-H., & Feng, M.-G. (2021). Genome-Wide Insight into Profound Effect of Carbon Catabolite Repressor (Cre1) on the Insect-Pathogenic Lifecycle of Beauveriabassiana. Journal of Fungi, 7(11), 895. https://doi.org/10.3390/jof7110895