
A Snapshot Picture of the Fungal Composition of Bee Bread in Four Locations in Bulgaria, Differing in Anthropogenic Influence

 , ,
, ,  ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
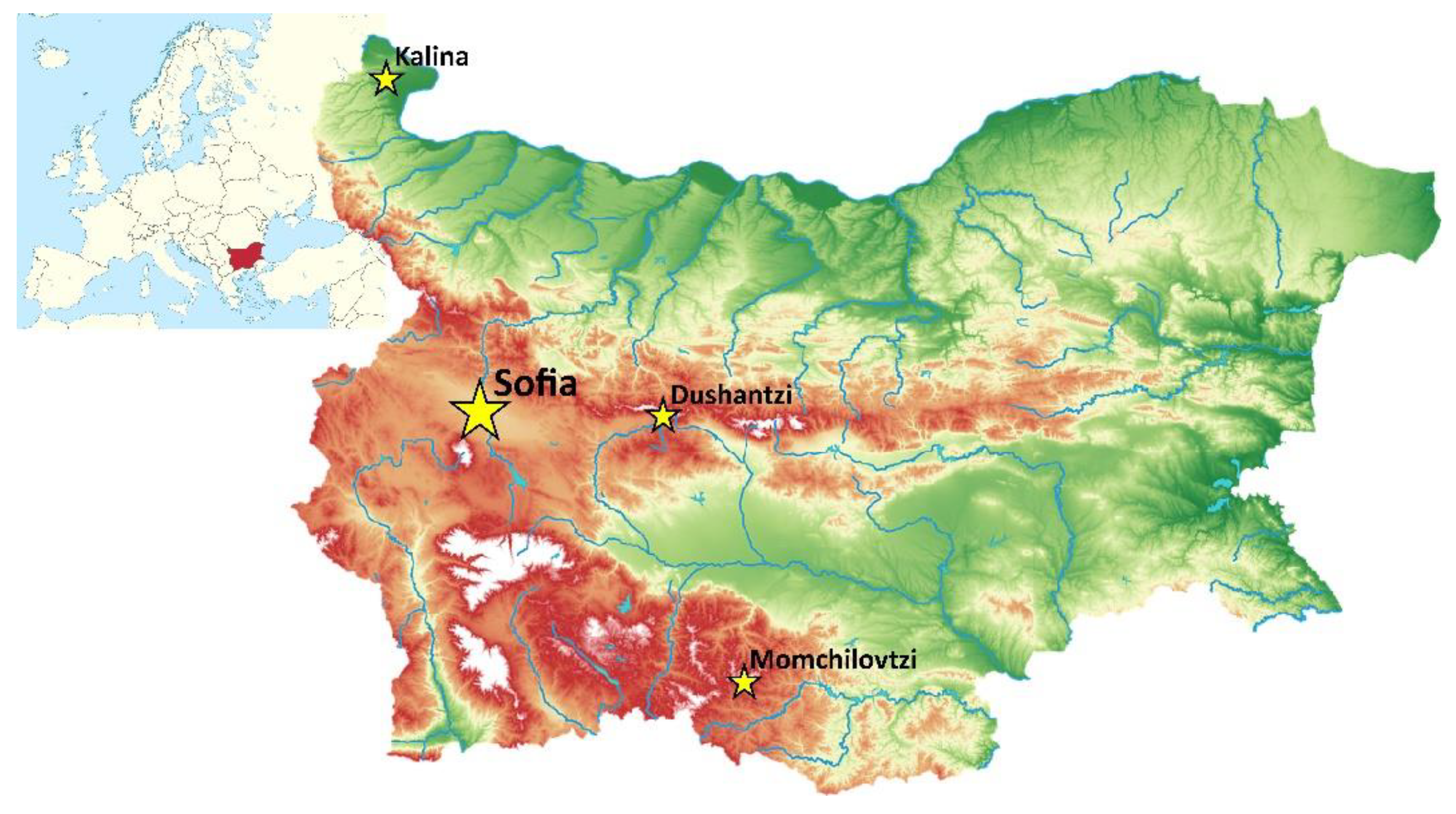
2.1. Sampling Locations
2.2. Bee Bread Sampling
2.3. DNA Isolation
2.4. Metagenomic Analyses
2.4.1. NGS-Sequencing
2.4.2. Sequences Processing
2.4.3. Otus Analyses
2.4.4. Alpha Diversity Analyses
3. Results
3.1. NGS Sequencing Results
3.2. OTUs Counts
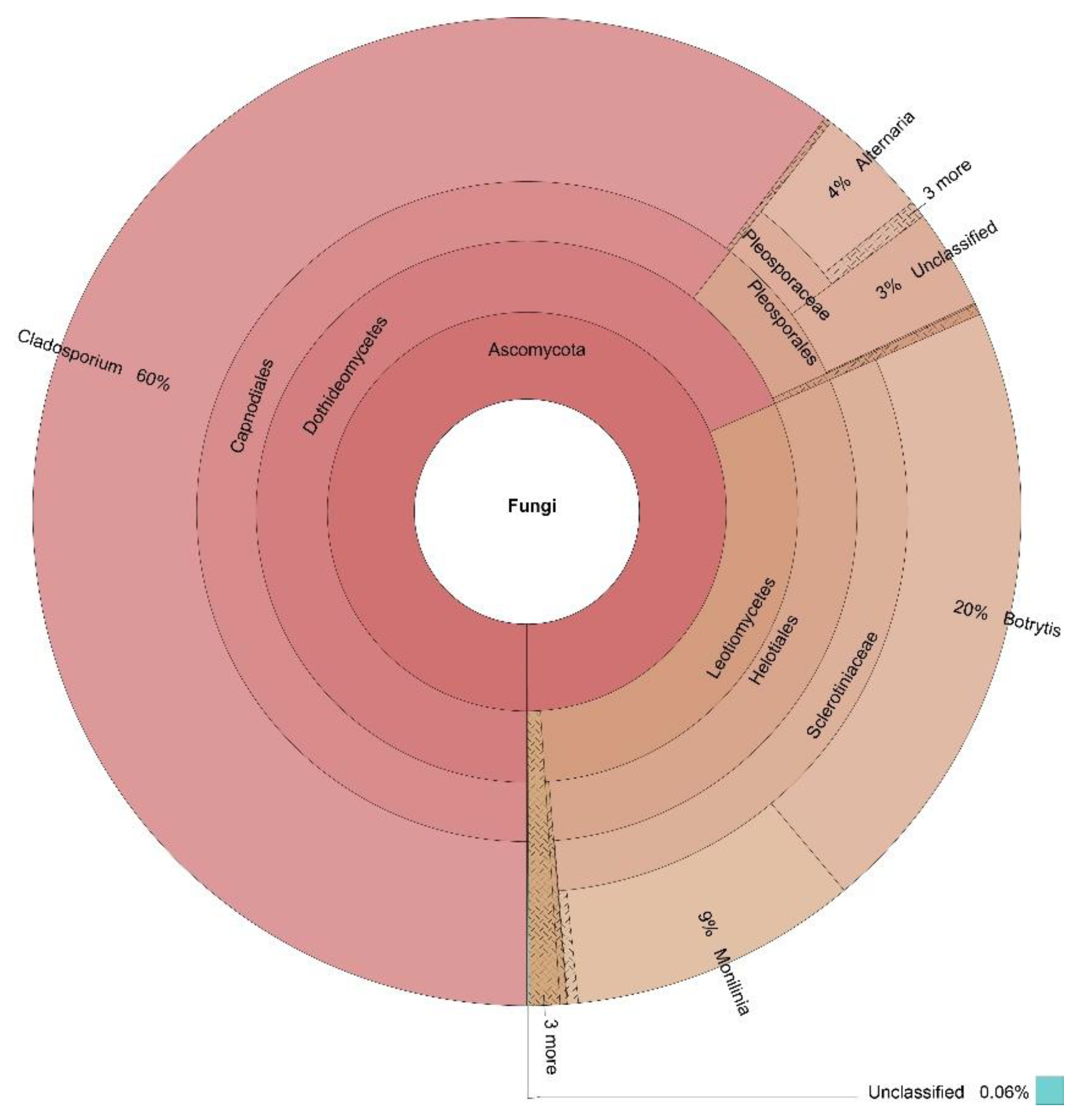
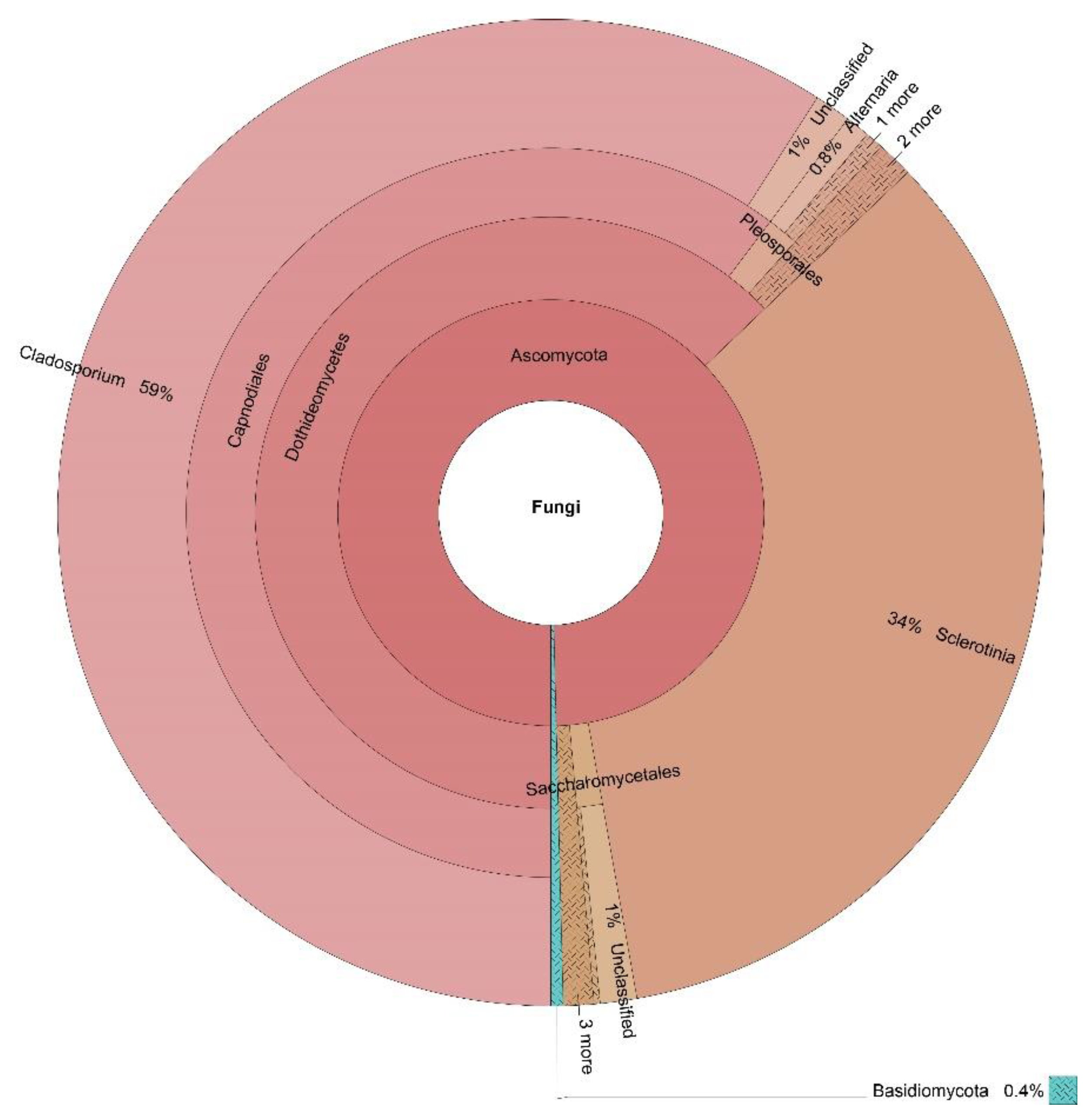
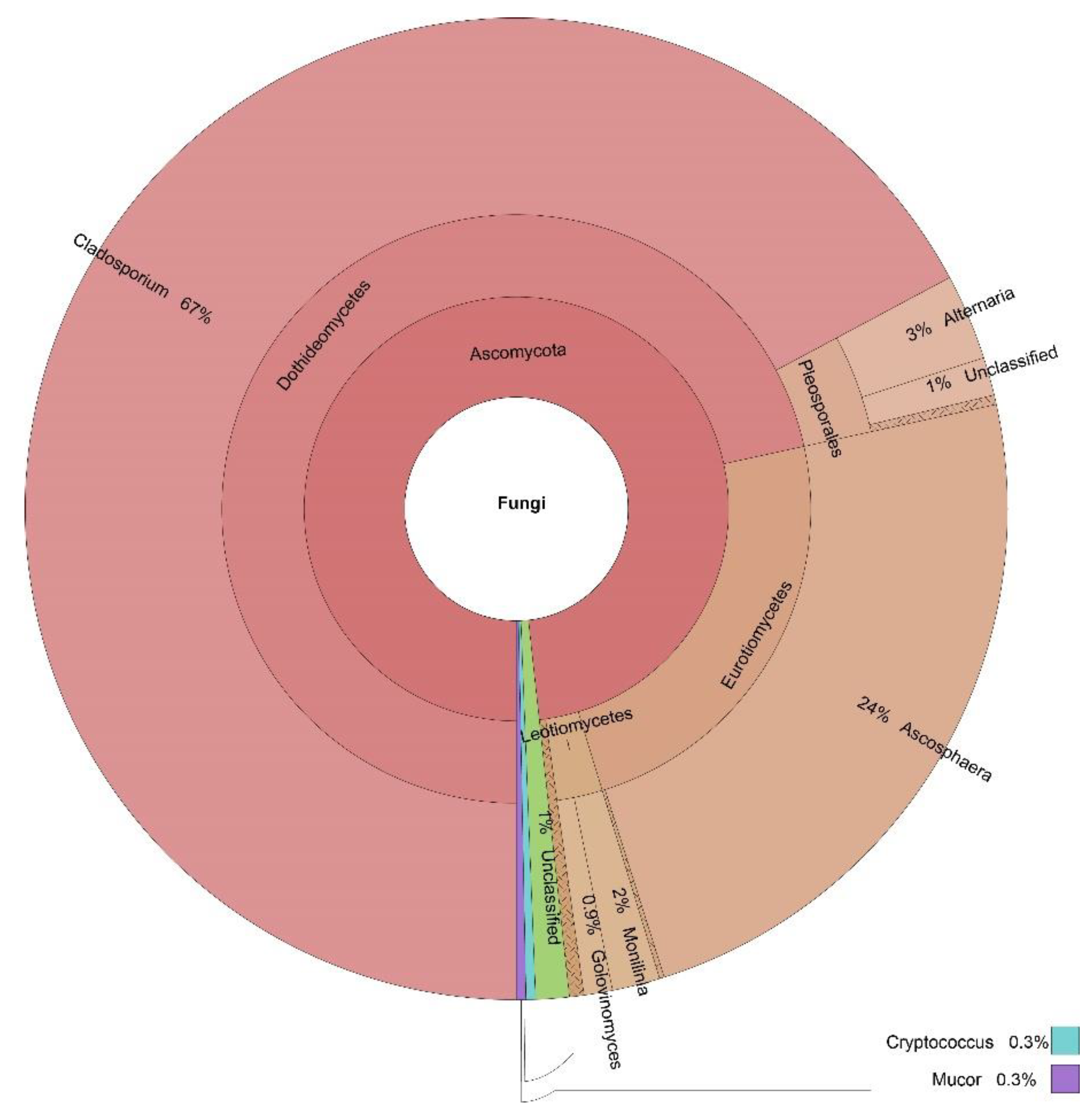
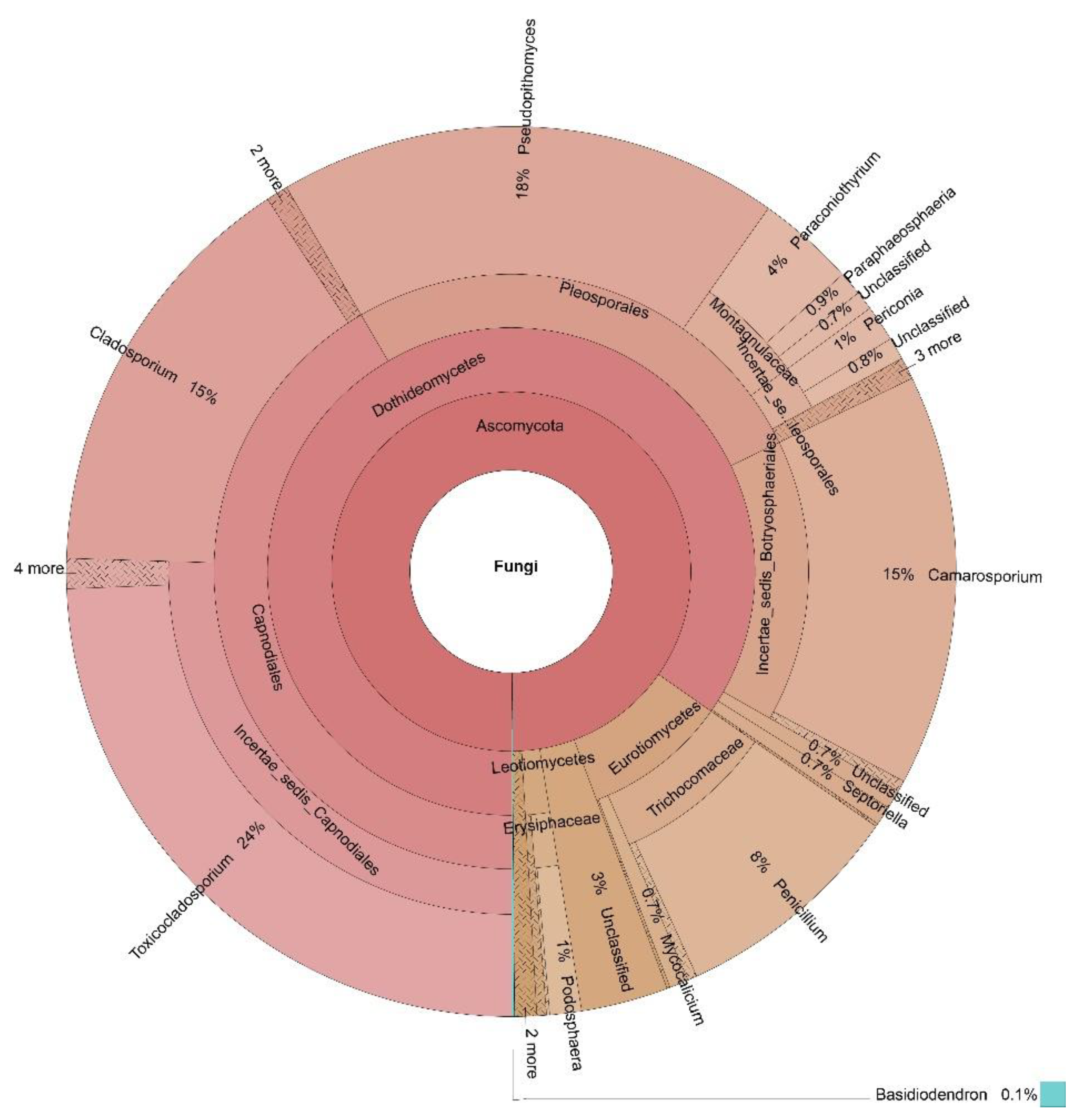
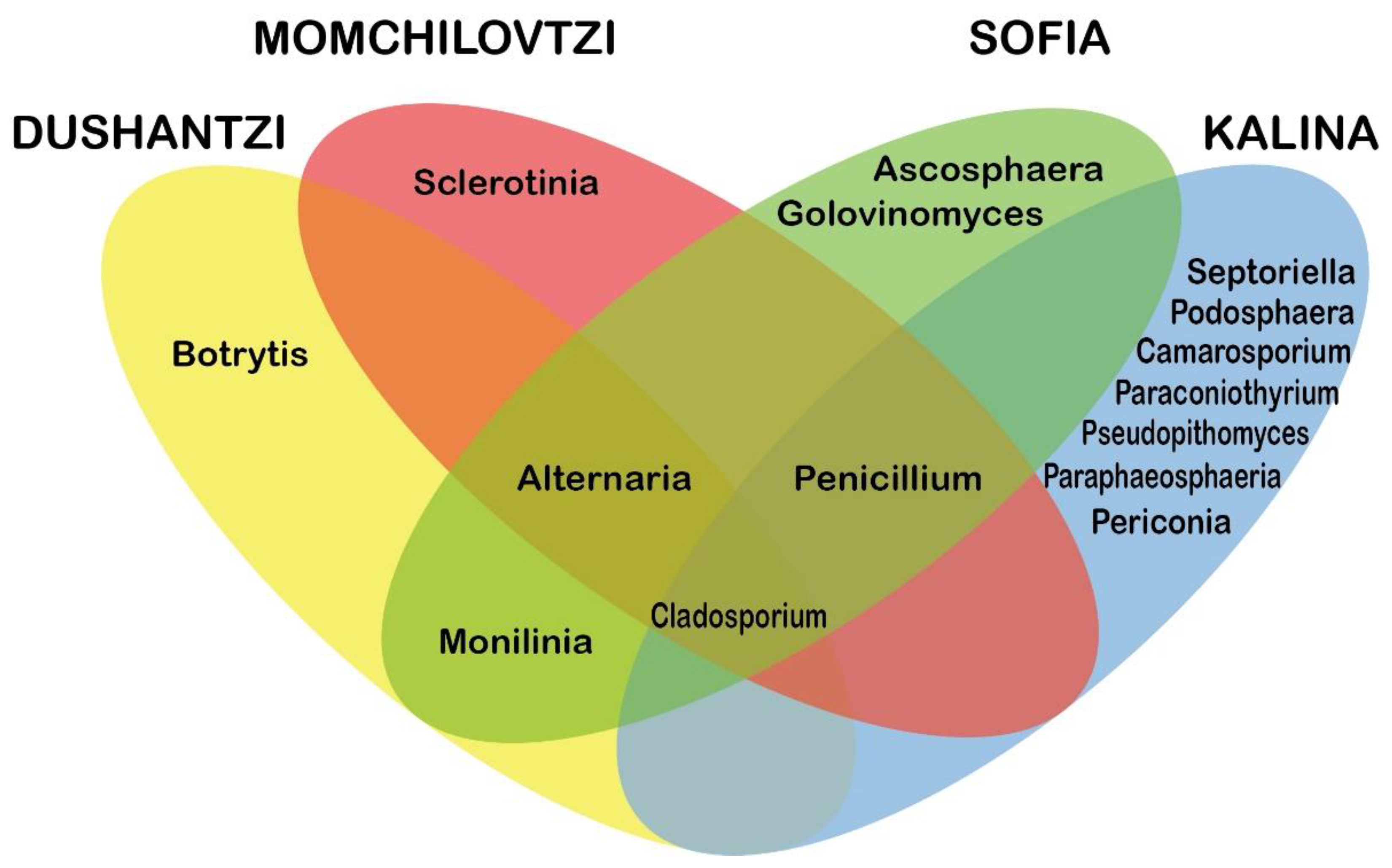
3.3. OTUs Taxonomic Annotation
3.4. Alpha Diversity Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bakour, M.; Fernandes, Â.; Barros, L.; Sokovic, M.; Ferreira, I.C.F.R.; Lyoussi, B. Bee bread as a functional product: Chemical composition and bioactive properties. LWT 2019, 109, 276–282. [Google Scholar] [CrossRef]
- Mărgăoan, R.; Stranț, M.; Varadi, A.; Topal, E.; Yücel, B.; Cornea-Cipcigan, M.; Campos, M.G.; Vodnar, D.C. Bee Collected Pollen and Bee Bread: Bioactive Constituents and Health Benefits. Antioxidants 2019, 8, 568. [Google Scholar] [CrossRef]
- Yucel, B.; Topal, E.; Kosoglu, M. Chapter 2: Bee products as functional food. In Superfood and Functional Food—An Overview of Their Processing and Utilization; InTech: London, UK, 2017; pp. 15–33. [Google Scholar]
- Ispirli, H.; Dertli, E. Detection of fructophilic lactic acid bacteria (FLAB) in bee bread and bee pollen samples and determination of their functional roles. J. Food Process. Preserv. 2021, 45, e15414. [Google Scholar] [CrossRef]
- Pełka, K.; Worobo, R.; Walkusz, J.; Szweda, P. Bee Pollen and Bee Bread as a Source of Bacteria Producing Antimicrobials. Antibiotics 2021, 10, 713. [Google Scholar] [CrossRef]
- Vásquez, A.; Olofsson, T.C. The lactic acid bacteria involved in the production of bee pollen and bee bread. J. Apic. Res. 2009, 48, 189–195. [Google Scholar] [CrossRef]
- Gilliam, M.; Prest, D.B.; Lorenz, B.J. Microbiology of pollen and bee bread: Taxonomy and enzymology of molds. Apidologie 1989, 20, 53–68. [Google Scholar] [CrossRef]
- Tauber, J.P.; McMahon, D.; Ryabov, E.V.; Kunat, M.; Ptaszyńska, A.A.; Evans, J.D. Honeybee intestines retain low yeast titers, but no bacterial mutualists, at emergence. Yeast 2021. [Google Scholar] [CrossRef]
- Sinpoo, C.; Williams, G.R.; Chantawannakul, P. Dynamics of fungal communities in corbicular pollen and bee bread. Chiang Mai J. Sci. 2017, 44, 1244–1256. [Google Scholar]
- Disayathanoowat, T.; Li, H.; Supapimon, N.; Suwannarach, N.; Lumyong, S.; Chantawannakul, P.; Guo, J. Different Dynamics of Bacterial and Fungal Communities in Hive-Stored Bee Bread and Their Possible Roles: A Case Study from Two Commercial Honey Bees in China. Microorganisms 2020, 8, 264. [Google Scholar] [CrossRef]
- Sinkevicine, J.; Amsiejus, A. Prevalence of microscopic fungi in bee pollen. Biologija 2019, 65, 41–47. [Google Scholar] [CrossRef]
- Yoder, J.A.; Jajack, A.J.; Rosselot, A.E.; Smith, T.J.; Yerke, M.C.; Sammataro, D. Fungicide Contamination Reduces Beneficial Fungi in Bee Bread Based on an Area-Wide Field Study in Honey Bee, Apis mellifera, Colonies. J. Toxicol. Environ. Health Part A 2013, 76, 587–600. [Google Scholar] [CrossRef]
- Hsu, C.-K.; Wang, D.-Y.; Wu, M.-C. A Potential Fungal Probiotic Aureobasidium melanogenum CK-CsC for the Western Honey Bee, Apis mellifera. J. Fungi 2021, 7, 508. [Google Scholar] [CrossRef]
- Dharampal, P.S.; Carlson, C.; Currie, C.R.; Steffan, S.A. Pollen-borne microbes shape bee fitness. Proc. R. Soc. B Boil. Sci. 2019, 286, 20182894. [Google Scholar] [CrossRef]
- González, G.; Hinojo, M.; Mateo-Castro, R.; Medina-Vaya, A.; Jiménez, M. Occurrence of mycotoxin producing fungi in bee pollen. Int. J. Food Microbiol. 2005, 105, 1–9. [Google Scholar] [CrossRef]
- Kostić, A.Ž.; Milinčić, D.D.; Petrović, T.S.; Krnjaja, V.S.; Stanojević, S.P.; Barać, M.B.; Tešić, Ž.L.; Pešić, M.B. Mycotoxins and mycotoxin producing fungi in pollen: Review. Toxins 2019, 11, 64. [Google Scholar] [CrossRef]
- Isayeva, A.F.; Seyidova, G.M.; Abdullayeva, S.A.; Novruzova, M.S.; Muradov, P.Z. Characteristic of Species Composition of Fungi Involved in the Formation of Mycobiota of Honey Bees in Azerbaijan. Adv. Biosci. Biotechnol. 2019, 10, 282–287. [Google Scholar] [CrossRef]
- Ye, M.-H.; Fan, S.-H.; Li, X.-Y.; Tarequl, I.M.; Yan, C.-X.; Wei, W.-H.; Yang, S.-M.; Zhou, B. Microbiota dysbiosis in honeybee (Apis mellifera L.) larvae infected with brood diseases and foraging bees exposed to agrochemicals. R. Soc. Open Sci. 2021, 8, 201805. [Google Scholar] [CrossRef]
- Ptaszyńska, A.; Latoch, P.; Hurd, P.; Polaszek, A.; Michalska-Madej, J.; Grochowalski, Ł.; Strapagiel, D.; Gnat, S.; Załuski, D.; Gancarz, M.; et al. Amplicon Sequencing of Variable 16S rRNA from Bacteria and ITS2 Regions from Fungi and Plants, Reveals Honeybee Susceptibility to Diseases Results from Their Forage Availability under Anthropogenic Landscapes. Pathogens 2021, 10, 381. [Google Scholar] [CrossRef]
- Ruttner, F. Chapter 14: Honeybees of the central Mediterranean and southeastern Europe. In Biogeography and Taxonomy of Honeybees; Gabler: Oslo, Norway, 1988; pp. 240–257. [Google Scholar]
- Magoč, T.; Salzberg, S. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef]
- Haas, B.J.; Gevers, D.; Earl, A.M.; Feldgarden, M.; Ward, D.V.; Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Highlander, S.K.; Sodergren, E.; et al. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011, 21, 494–504. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kõljalg, U.; Nilsson, H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef]
- Ondov, B.D.; Bergman, N.H.; Phillippy, A.M. Interactive metagenomic visualization in a Web browser. BMC Bioinform. 2011, 12, 385. [Google Scholar] [CrossRef]
- Chao, A. Non-parametric estimation of the classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Chao, A.; Lee, S.M. Estimating the number of classes via sample coverage. J. Am. Stat. Assoc. 1992, 87, 210–217. [Google Scholar] [CrossRef]
- Chao, A.; Yang, M.C.K. Stopping Rules and Estimation for Recapture Debugging with Unequal Failure Rates. Biometrika 1993, 80, 193. [Google Scholar] [CrossRef]
- Lemos, L.N.; Fulthorpe, R.R.; Triplett, E.W.; Roesch, L.F. Rethinking microbial diversity analysis in the high throughput sequencing era. J. Microbiol. Methods 2011, 86, 42–51. [Google Scholar] [CrossRef]
- McCarthy, B.C.; Magurran, A.E. Measuring Biological Diversity. J. Torrey Bot. Soc. 2004, 131, 277. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Chao, A.; Lee, S.-M.; Chen, T.-C. A generalized good’s nonparametric coverage estimator. Chin. J. Math. 1988, 16, 189–199. [Google Scholar]
- Faith, D.P. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 1992, 61, 1–10. [Google Scholar] [CrossRef]
- Buée, M.; Reich, M.; Murat, C.; Morin, E.; Nilsson, R.H.; Uroz, S.; Martin, F. 454 Pyrosequencing analyses of forest soils reveal an unexpectedly high fungal diversity. New Phytol. 2009, 184, 449–456. [Google Scholar] [CrossRef]
- Lepère, C.; Domaizon, I.; Humbert, J.-F.; Jardillier, L.; Hugoni, M.; Debroas, D. Diversity, spatial distribution and activity of fungi in freshwater ecosystems. PeerJ 2019, 7, e6247. [Google Scholar] [CrossRef]
- Wang, Y.; Sen, B.; He, Y.; Xie, N.; Wang, G. Spatiotemporal Distribution and Assemblages of Planktonic Fungi in the Coastal Waters of the Bohai Sea. Front. Microbiol. 2018, 9, 584. [Google Scholar] [CrossRef]
- Rich, J.O.; Anderson, A.M.; Leathers, T.D.; Bischoff, K.M.; Liu, S.; Skory, C.D. Microbial contamination of commercial corn-based fuel ethanol fermentations. Bioresour. Technol. Rep. 2020, 11, 100433. [Google Scholar] [CrossRef]
- Aregbe, A.Y.; Mu, T.; Sun, H. Effect of different pretreatment on the microbial diversity of fermented potato revealed by high-throughput sequencing. Food Chem. 2019, 290, 125–134. [Google Scholar] [CrossRef]
- Kiš, M.; Furmeg, S.; Tkalec, V.J.; Zadravec, M.; Lugomer, M.D.; Končurat, A.; Benić, M.; Pavliček, D. Characterisation of Croatian honey by physicochemical and microbiological parameters with mold identification. J. Food Saf. 2018, 38, e12492. [Google Scholar] [CrossRef]
- Kiš, M.; Furmeg, S.; Tkalec, V.; Zadravec, M.; Benić, M.; Sokolović, J.; Majnarić, D. Identification of moulds from Croatian honey. Qual. Assur. Saf. Crop. Foods 2019, 11, 571–576. [Google Scholar] [CrossRef]
- Barbosa, R.; Bezerra, J.D.P.; Souza-Motta, C.M.; Frisvad, J.; Samson, R.A.; Oliveira, N.T.; Houbraken, J. New Penicillium and Talaromyces species from honey, pollen and nests of stingless bees. Antonie van Leeuwenhoek 2018, 111, 1883–1912. [Google Scholar] [CrossRef]
- Anderson, K.E.; Sheehan, T.H.; Eckholm, B.; Mott, B.M.; Degrandihoffman, G. An emerging paradigm of colony health: Microbial balance of the honey bee and hive (Apis mellifera). Insectes Sociaux 2011, 58, 431–444. [Google Scholar] [CrossRef]
- Marin-Felix, Y.; Restrepo, M.H.; Wingfield, M.; Akulov, O.; Carnegie, A.; Cheewangkoon, R.; Gramaje, D.; Groenewald, J.; Guarnaccia, V.; Halleen, F.; et al. Genera of phytopathogenic fungi: GOPHY 2. Stud. Mycol. 2019, 92, 47–133. [Google Scholar] [CrossRef] [PubMed]
- Batra, L.R.; Batra, S.W.T. Floral mimicry induced by mummy-berry fungus exploits host’s pollinators as vectors. Science 1985, 228, 1011. [Google Scholar] [CrossRef]
- Parish, J.B.; Scott, E.S.; Hogendoorn, K. Nutritional benefit of fungal spores for honey bee workers. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Boland, G.; Hall, R. Index of plant hosts of Sclerotinia sclerotiorum. Can. J. Plant Pathol. 1994, 16, 93–108. [Google Scholar] [CrossRef]
- Ngalimat, M.S.; Rahman, R.N.Z.R.A.; Yusof, M.T.; Hamzah, A.S.A.; Zawawi, N.; Sabri, S. A Review on the Association of Bacteria with Stingless Bees. Sains Malays. 2020, 49, 1853–1863. [Google Scholar] [CrossRef]
- Janashia, I.; Choiset, Y.; Józefiak, D.; Déniel, F.; Coton, E.; Moosavi-Movahedi, A.A.; Chanishvili, N.; Haertlé, T. Beneficial Protective Role of Endogenous Lactic Acid Bacteria Against Mycotic Contamination of Honeybee Beebread. Probiotics Antimicrob. Proteins 2018, 10, 638–646. [Google Scholar] [CrossRef] [PubMed]
- Takamatsu, S.; Matsuda, S.; Grigaliunaite, B. Comprehensive phylogenetic analysis of the genus Golovinomyces (Ascomycota: Erysiphales) reveals close evolutionary relationships with its host plants. Mycologia 2013, 105, 1135–1152. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, M.; Kim Hyun, S.; Gurung Sun, K.; Bazie, S.; Lee Hyun, G.; Lee Hyang, B.; Lee Youn, S. Two new records of ascomycetes from crop field soils in korea. Korean J. Mycol. 2017, 45, 167–174. [Google Scholar] [CrossRef][Green Version]
- Fotedar, R.; Sandoval-Denis, M.; Kolecka, A.; Zeyara, A.; Al Malki, A.; Al Shammari, H.; Al Marri, M.; Kaul, R.; Boekhout, T. Toxicocladosporium aquimarinum sp. nov. and Toxicocladosporium qatarense sp. nov., isolated from marine waters of the Arabian Gulf surrounding Qatar. Int. J. Syst. Evol. Microbiol. 2019, 69, 2992–3000. [Google Scholar] [CrossRef]
- Hashimoto, K.; Oda, H.; Saito, Y.; Akimoto, M.; Nojiri, T.; Kawakami, Y. Isolation of simplicillium sympodiophorum and toxicocladosporium irritans from the blowout air of household air conditioners. Biocontrol Sci. 2021, 26, 105–111. [Google Scholar] [CrossRef]
- Crous, P.; Wingfield, M.; Richardson, D.; Le Roux, J.; Strasberg, D.; Edwards, J.; Roets, F.; Hubka, V.; Taylor, P.; Heykoop, M.; et al. Fungal Planet description sheets: 400–468. Pers. Mol. Phylogeny Evol. Fungi 2016, 36, 316–458. [Google Scholar] [CrossRef]
- Bezerra, J.D.P.; Sandoval-Denis, M.; Paiva, L.M.; Silva, G.; Groenewald, J.Z.; Souza-Motta, C.M.; Crous, P.W. New endophytic Toxicocladosporium species from cacti in Brazil, and description of Neocladosporium gen. nov. IMA Fungus 2017, 8, 77–97. [Google Scholar] [CrossRef]
- Moubasher, A.; Abdel-Sater, M.; Soliman, Z.S. Diversity of floricolous yeasts and filamentous fungi of some ornamental and edible fruit plants in assiut area, egypt. Current Research in Environmental & Applied Mycology. J. Fungal Biol. 2018, 8, 135–161. [Google Scholar] [CrossRef]
- Brentassi, M.E.; Medina, R.; de la Fuente, D.; Franco, M.E.; Toledo, A.V.; Saparrat, M.C.; Balatti, P.A. Endomycobiome associated with females of the planthopper Delphacodes kuscheli (Hemiptera: Delphacidae): A metabarcoding approach. Heliyon 2020, 6, e04634. [Google Scholar] [CrossRef]
- Wu, H.; Rao, Z.-C.; Cao, L.; De Clercq, P.; Han, R.-C. Infection of Ophiocordyceps sinensis Fungus Causes Dramatic Changes in the Microbiota of Its Thitarodes Host. Front. Microbiol. 2020, 11, 577268. [Google Scholar] [CrossRef] [PubMed]
- Goh, J.; Jeon, Y.-J.; Mun Hye, Y.; Chung, N.; Park, Y.-H.; Park, S.; Hwang, H.; Cheon, W. Isolation and characterization of eleven unrecorded pezizomycotina species from freshwater ecosystems in korea. Korean J. Mycol. 2020, 48, 423–443. [Google Scholar] [CrossRef]
- Ningsih, B.N.S.; Rukachaisirikul, V.; Pansrinun, S.; Phongpaichit, S.; Preedanon, S.; Sakayaroj, J. New aromatic polyketides from the marine-derived fungus Pseudopithomyces maydicus PSU-AMF350 and their antimicrobial activity. Nat. Prod. Res. 2021, 1–8. [Google Scholar] [CrossRef]
- Chin, J.M.W.; Puchooa, D.; Bahorun, T.; Jeewon, R. Molecular characterization of marine fungi associated with Haliclona sp. (sponge) and Turbinaria conoides and Sargassum portierianum (brown algae). In Proceedings of the National Academy of Sciences, India Section B: Biological Sciences; Gabler: Oslo, Norway, 2021; pp. 1–14. [Google Scholar]
- Yuan, X.-L.; Cao, M.; Liu, X.-M.; Du, Y.-M.; Shen, G.-M.; Zhang, Z.-F.; Li, J.-H.; Zhang, P. Composition and Genetic Diversity of the Nicotiana tabacum Microbiome in Different Topographic Areas and Growth Periods. Int. J. Mol. Sci. 2018, 19, 3421. [Google Scholar] [CrossRef] [PubMed]
- Perelló, A.; Aulicino, M.; Stenglein, S.A.; Labuda, R.; Moreno, M.V. Pseudopithomyces chartarum associated with wheat seeds in Argentina, pathogenicity and evaluation of toxigenic ability. Eur. J. Plant Pathol. 2017, 148, 491–496. [Google Scholar] [CrossRef]
- You, N.; Xu, J.; Wang, L.; Zhuo, L.; Zhou, J.; Song, Y.; Ali, A.; Luo, Y.; Yang, J.; Yang, W.; et al. Fecal Fungi Dysbiosis in Nonalcoholic Fatty Liver Disease. Obesity 2021, 29, 350–358. [Google Scholar] [CrossRef]
- Majumder, R.; Sutcliffe, B.; Taylor, P.W.; Chapman, T.A. Microbiome of the Queensland Fruit Fly through Metamorphosis. Microorganisms 2020, 8, 795. [Google Scholar] [CrossRef]
- Damm, U.; Verkley, G.; Crous, P.; Fourie, P.; Haegi, A.; Riccioni, L. Novel Paraconiothyrium species on stone fruit trees and other woody hosts. Pers. Mol. Phylogeny Evol. Fungi 2008, 20, 9–17. [Google Scholar] [CrossRef]
- Perazzolli, M.; Nesler, A.; Giovannini, O.; Antonielli, L.; Puopolo, G.; Pertot, I. Ecological impact of a rare sugar on grapevine phyllosphere microbial communities. Microbiol. Res. 2020, 232, 126387. [Google Scholar] [CrossRef]
- Kevan, P.G.; Shipp, L.; Smagghe, G. Ecological intensification: Managing biocomplexity and biodiversity in agriculture through pollinators, pollination and deploying biocontrol agents against crop and pollinator diseases, pests and parasites. In Entomovectoring for Precision Biocontrol and Enhanced Pollination of Crops; Gabler: Oslo, Norway, 2020; pp. 19–51. [Google Scholar]
- Dantas, S.B.S.; Alves, F.A.M.; Chapla, V.M. Chemical, diversity and biotechnological potential of endophytic fungi isolated from Brazilian Cerrado plants. Biota Neotrop. 2021, 21, 20201069. [Google Scholar] [CrossRef]
- Pérez-Atanes, S.; Seijo-Coello, M.D.C.; Méndez-Alvarez, J. Contribution to the study of fungal spores in honeys of Galicia (NW Spain). Grana 2001, 40, 217–222. [Google Scholar] [CrossRef]
- Shao, M.-W.; Lu, Y.-H.; Miao, S.; Zhang, Y.; Chen, T.-T.; Zhang, Y.-L. Diversity, Bacterial Symbionts and Antibacterial Potential of Gut-Associated Fungi Isolated from the Pantala flavescens Larvae in China. PLoS ONE 2015, 10, e0134542. [Google Scholar] [CrossRef] [PubMed]
- Huseyin, E.; Selcuk, F.; Akgul, H. Species of camarosporium on forest trees and shrubs new reports from anatolian peninsula. Int. J. For. Hortic. 2017, 3, 1–8. [Google Scholar]
- Magyar, D.; Mura-Mészáros, A.; Grillenzoni, F.; Information, R. Fungal diversity in floral and honeydew honeys. Acta Bot. Hung. 2016, 58, 145–166. [Google Scholar] [CrossRef]
- Azhari, A.; Supratman, U. The Chemistry and Pharmacology of Fungal Genus Periconia: A Review. Sci. Pharm. 2021, 89, 34. [Google Scholar] [CrossRef]
- Marin-Felix, Y.; Restrepo, M.H.; Iturrieta-González, I.; García, D.; Gené, J.; Groenewald, J.; Cai, L.; Chen, Q.; Quaedvlieg, W.; Schumacher, R.; et al. Genera of phytopathogenic fungi: GOPHY 3. Stud. Mycol. 2019, 94, 1–124. [Google Scholar] [CrossRef] [PubMed]
- Gunasekaran, R.; Janakiraman, D.; Rajapandian, S.G.K.; Appavu, S.P.; Venkatesh, P.N.; Prajna, L. Periconia species-An unusual fungal pathogen causing mycotic keratitis. Indian J. Med. Microbiol. 2021, 39, 36–40. [Google Scholar] [CrossRef]
- Egerer, M.; Kowarik, I. Confronting the Modern Gordian Knot of Urban Beekeeping. Trends Ecol. Evol. 2020, 35, 956–959. [Google Scholar] [CrossRef]
- Sponsler, D.B.; Bratman, E.Z. Beekeeping in, of or for the city? A socioecological perspective on urban apiculture. People Nat. 2021, 3, 550–559. [Google Scholar] [CrossRef]
- Giner, C.R.; Forn, I.; Romac, S.; Logares, R.; de Vargas, C.; Massana, R. Environmental Sequencing Provides Reasonable Estimates of the Relative Abundance of Specific Picoeukaryotes. Appl. Environ. Microbiol. 2016, 82, 4757–4766. [Google Scholar] [CrossRef]
- Zheng, X.; Liu, F.; Shi, X.; Wang, B.; Li, K.; Li, B.; Zhuge, B. Dynamic correlations between microbiota succession and flavor development involved in the ripening of Kazak artisanal cheese. Food Res. Int. 2018, 105, 733–742. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Number of the Raw Paired-End Reads | Number of the Raw Tags | Number of the Clean Tags | Number of the Effective Tags | Number of Bases of the Effective Tags (Nt) | Average Length of the Effective Tags (Nt) | Q20 Value | Q30 Value | GC Content Percentage in Effective Tags | Percentage of Effective Tags in Raw Paired End |
|---|---|---|---|---|---|---|---|---|---|---|
| Dushantzi | 139,921 | 138,997 | 138,469 | 137,837 | 50,312,892 | 365 | 99.39 | 97.57 | 54.30 | 98.51 |
| Momchilovtzi | 116,925 | 116,125 | 115,783 | 115,635 | 41,254,729 | 357 | 99.49 | 97.95 | 58.24 | 98.90 |
| Sofia | 86,102 | 84,754 | 83,876 | 83,679 | 30,482,594 | 364 | 98.24 | 94.94 | 59.32 | 97.19 |
| Kalina | 112,607 | 108,655 | 102,645 | 102,273 | 37,731,509 | 369 | 97.10 | 92.14 | 62.91 | 90.82 |
| Location | Total Tags | Unclassified Tags | Taxon Tags | % of the Unclassified Tags | Unique Tags | % of the Unique Tags | OTUs |
|---|---|---|---|---|---|---|---|
| Dushantzi | 275,674 | 267,432 | 3556 | 97.01% | 4686 | 1.70% | 169 |
| Momchilovtzi | 231,270 | 224,174 | 504 | 96.93% | 6592 | 2.85% | 156 |
| Sofia | 167,358 | 157,204 | 638 | 93.93% | 9516 | 5.69% | 185 |
| Kalina | 204,546 | 201,080 | 1742 | 98.31% | 1724 | 0.84% | 281 |
| Dushantzi | Momchilovtzi | Sofia | Kalina | |
|---|---|---|---|---|
| Ascomycota | 99.94% | 99.96% | 99.40% | 99.90% |
| Basidiomycota | - | 0.04% | 0.30% | 0.10% |
| Mucoromycota | - | - | 0.30% | - |
| Unclassified | 0.06% | - | - | - |
| Dushantzi | Momchilovtzi | Sofia | Kalina | Average | |
|---|---|---|---|---|---|
| Cladosporium | 60% | 59% | 67% | 15% | 54% |
| Penicillium | 0% | 1% | 1% | 8% | 3% |
| Alternaria | 4% | 1% | 3% | 0% | 2% |
| Monilinia | 9% | 0% | 2% | 0% | 3% |
| Sclerotinia | 0% | 34% | 0% | 0% | 9% |
| Ascosphaera | 0% | 0% | 24% | 0% | 6% |
| Toxicocladosporium | 0% | 0% | 0% | 24% | 6% |
| Botrytis | 20% | 0% | 0% | 0% | 5% |
| Pseudopithomyces | 0% | 0% | 0% | 18% | 5% |
| Camarosporium | 0% | 0% | 0% | 15% | 4% |
| Paraconiothyrium | 0% | 0% | 0% | 4% | 1% |
| Podosphaera | 0% | 0% | 0% | 1% | 0% |
| Golovinomyces | 0% | 0% | 1% | 0% | 0% |
| Paraphaeosphaeria | 0% | 0% | 0% | 1% | 0% |
| Periconia | 0% | 0% | 0% | 1% | 0% |
| Septoriella | 0% | 0% | 0% | 1% | 0% |
| Location | Observed Species | Shannon | Simpson | Chao1 | ACE | Goods Coverage | PD Whole Tree |
|---|---|---|---|---|---|---|---|
| Dushantzi | 169 | 2.140 | 0.635 | 180.538 | 183.600 | 1.000 | 14.665 |
| Momchilovtzi | 156 | 2.963 | 0.727 | 171.120 | 177.229 | 1.000 | 13.575 |
| Sofia | 185 | 2.924 | 0.795 | 200.120 | 204.572 | 1.000 | 12.889 |
| Kalina | 281 | 2.178 | 0.506 | 281.000 | 281.000 | 1.000 | 17.992 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dimov, S.G.; Zagorchev, L.; Iliev, M.; Dekova, T.; Ilieva, R.; Kitanova, M.; Georgieva-Miteva, D.; Dimitrov, M.; Peykov, S. A Snapshot Picture of the Fungal Composition of Bee Bread in Four Locations in Bulgaria, Differing in Anthropogenic Influence. J. Fungi 2021, 7, 845. https://doi.org/10.3390/jof7100845
Dimov SG, Zagorchev L, Iliev M, Dekova T, Ilieva R, Kitanova M, Georgieva-Miteva D, Dimitrov M, Peykov S. A Snapshot Picture of the Fungal Composition of Bee Bread in Four Locations in Bulgaria, Differing in Anthropogenic Influence. Journal of Fungi. 2021; 7(10):845. https://doi.org/10.3390/jof7100845
Chicago/Turabian StyleDimov, Svetoslav G., Lyuben Zagorchev, Mihail Iliev, Tereza Dekova, Ralitza Ilieva, Meglena Kitanova, Dimitrina Georgieva-Miteva, Martin Dimitrov, and Slavil Peykov. 2021. "A Snapshot Picture of the Fungal Composition of Bee Bread in Four Locations in Bulgaria, Differing in Anthropogenic Influence" Journal of Fungi 7, no. 10: 845. https://doi.org/10.3390/jof7100845
APA StyleDimov, S. G., Zagorchev, L., Iliev, M., Dekova, T., Ilieva, R., Kitanova, M., Georgieva-Miteva, D., Dimitrov, M., & Peykov, S. (2021). A Snapshot Picture of the Fungal Composition of Bee Bread in Four Locations in Bulgaria, Differing in Anthropogenic Influence. Journal of Fungi, 7(10), 845. https://doi.org/10.3390/jof7100845