
Multi-Gene Phylogeny and Taxonomy of Hydnellum (Bankeraceae, Basidiomycota) from China


Abstract
1. Introduction
2. Materials and Methods
2.1. Morphological Studies
2.2. Molecular Procedures and Phylogenetic Analyses
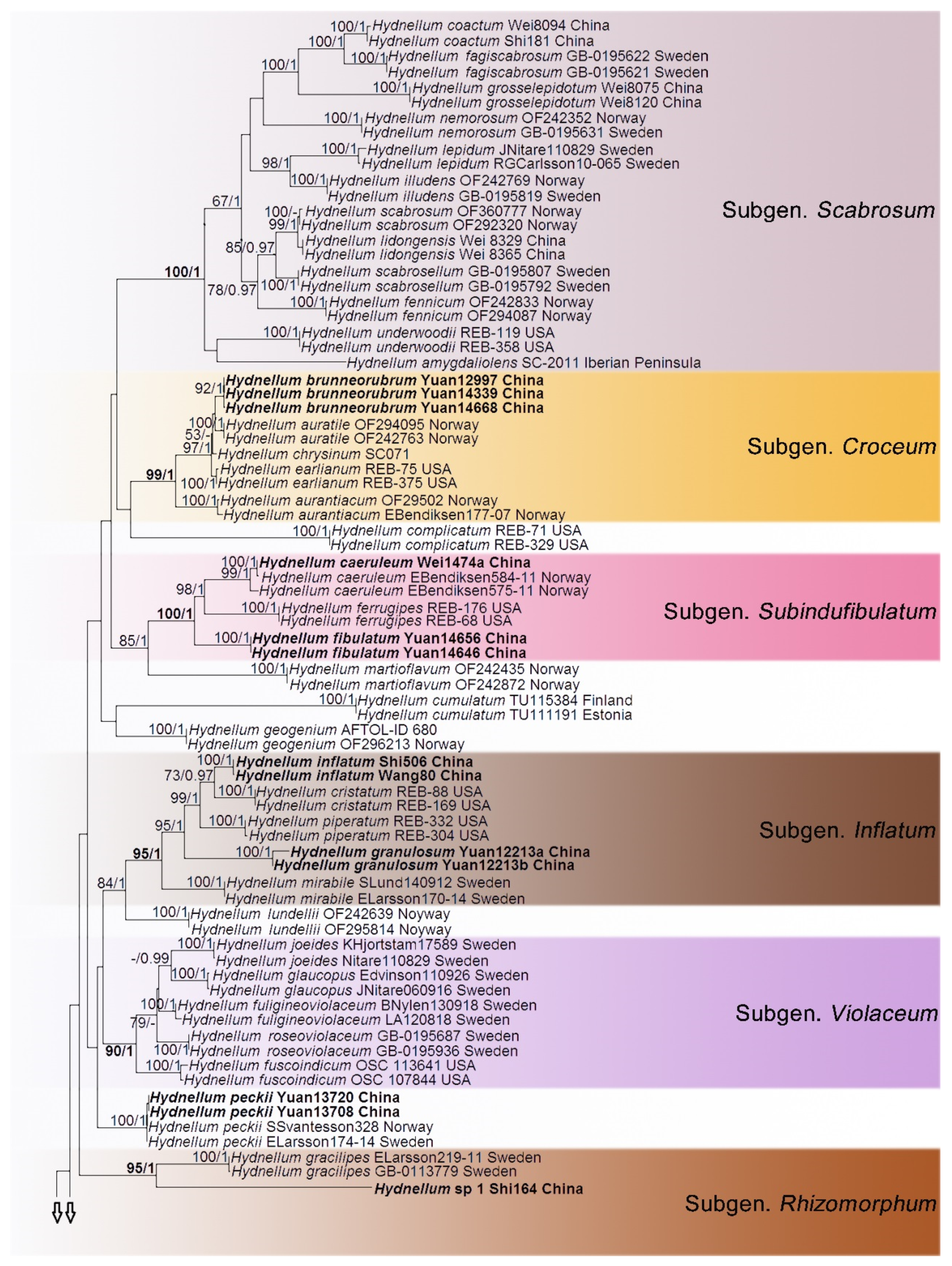
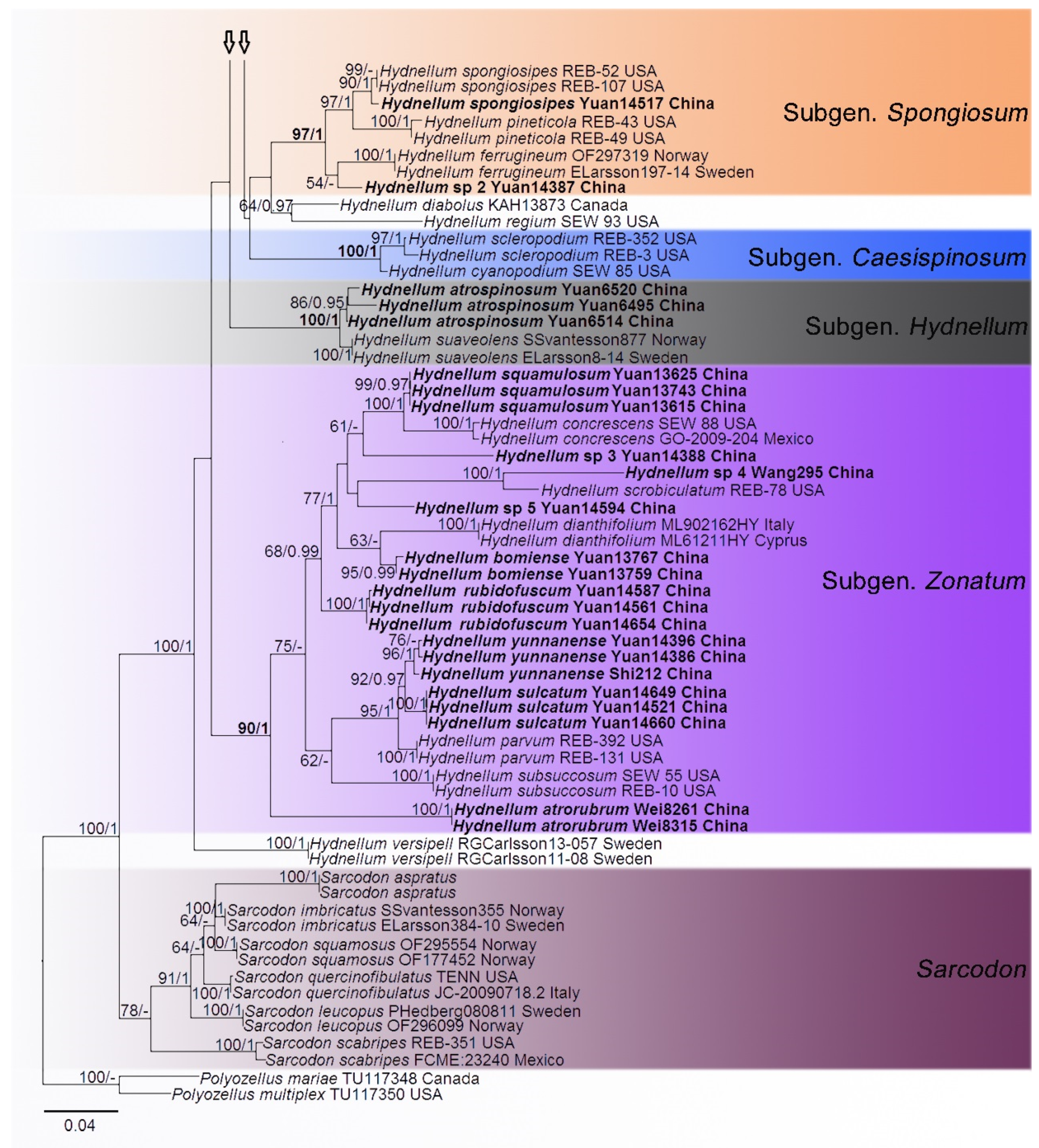
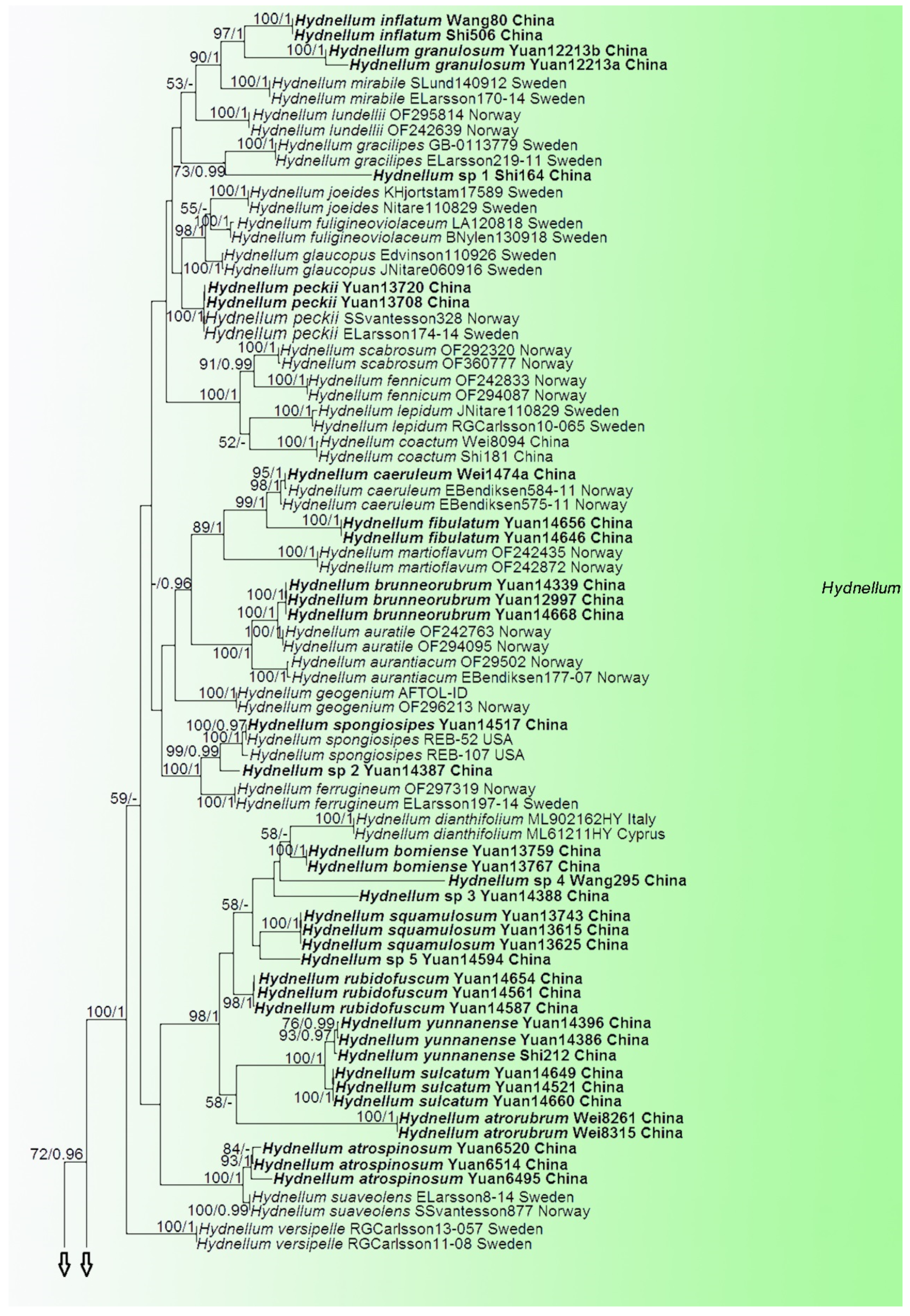
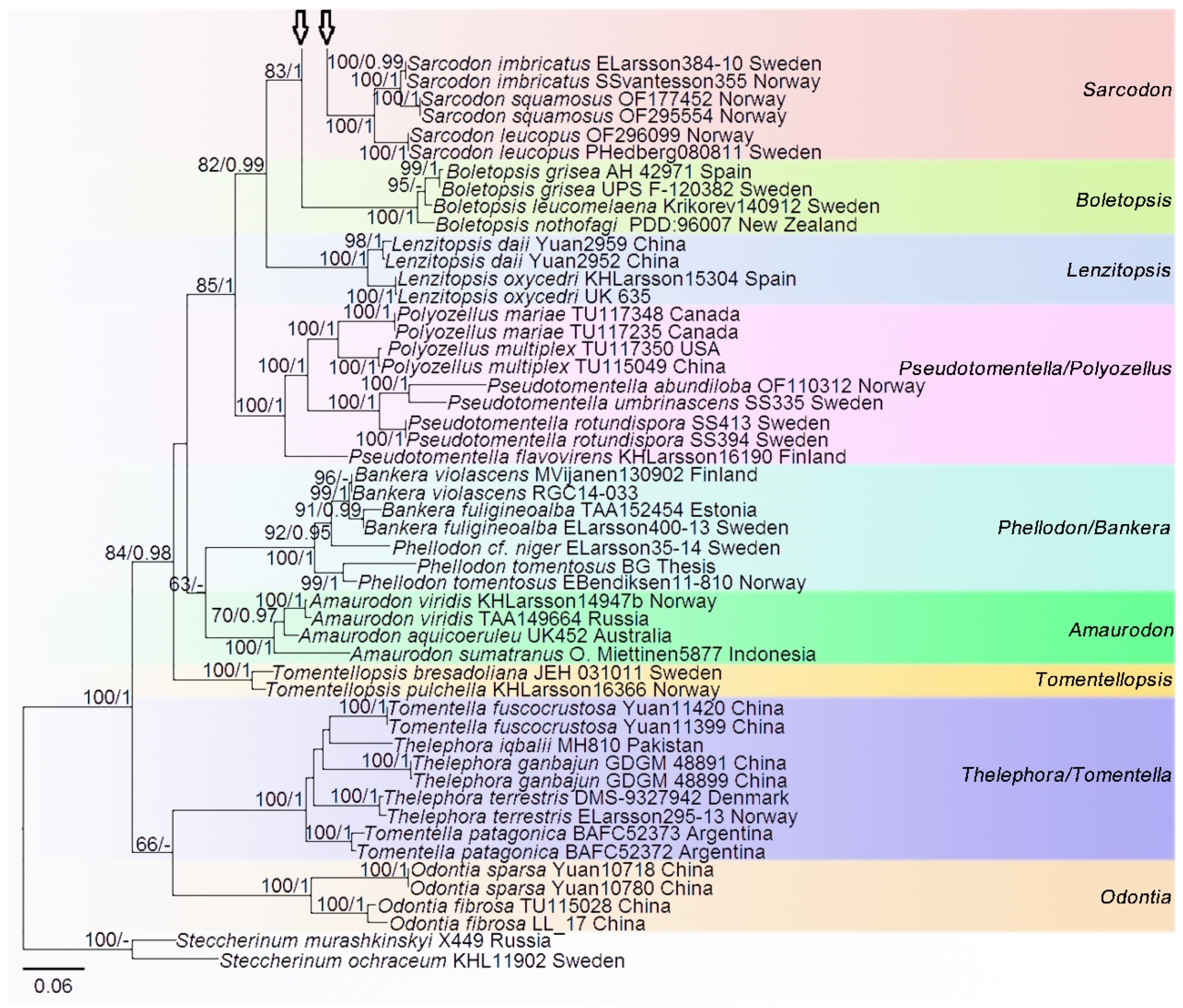
3. Results
Phylogenetic Analyses
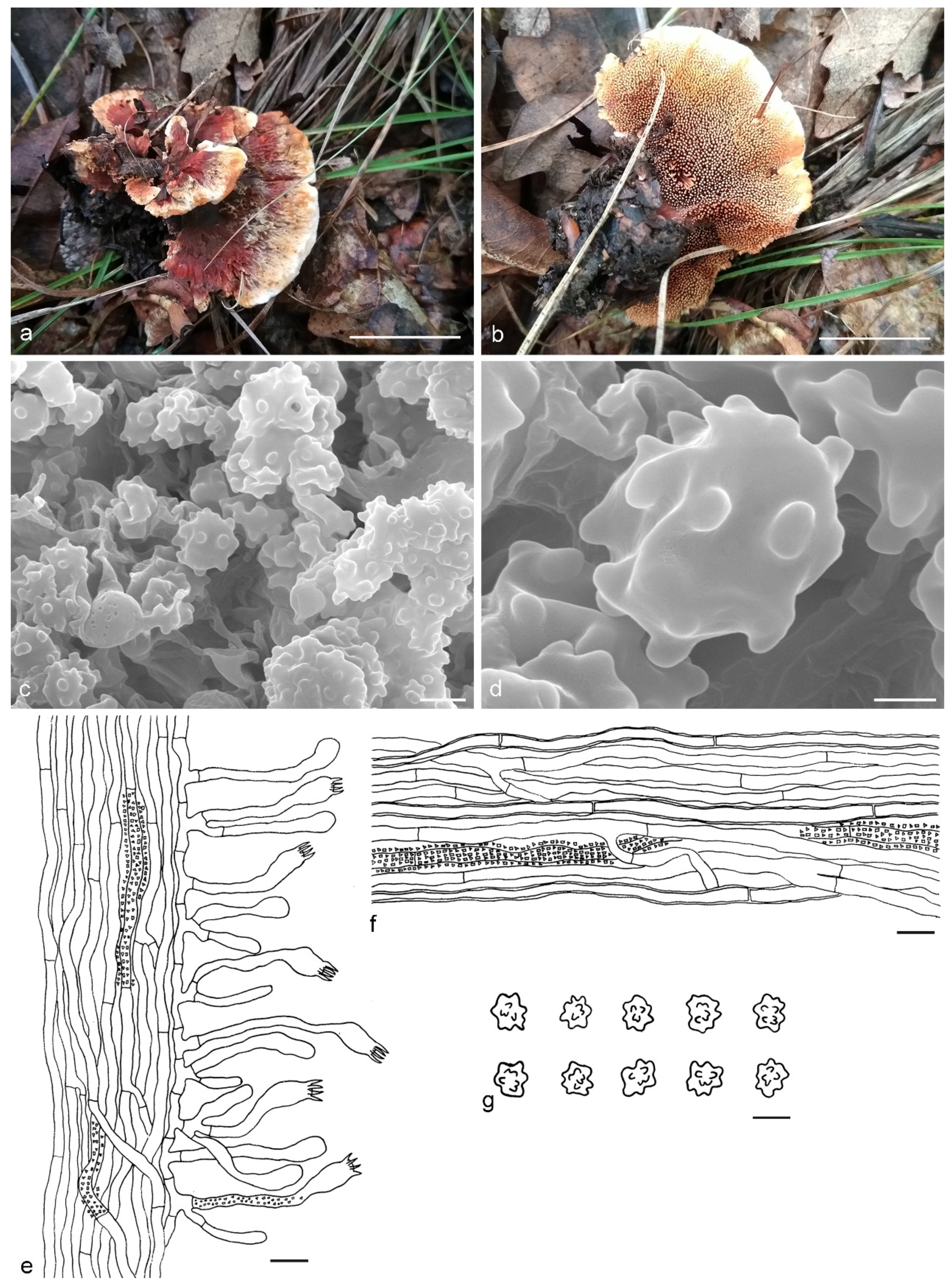
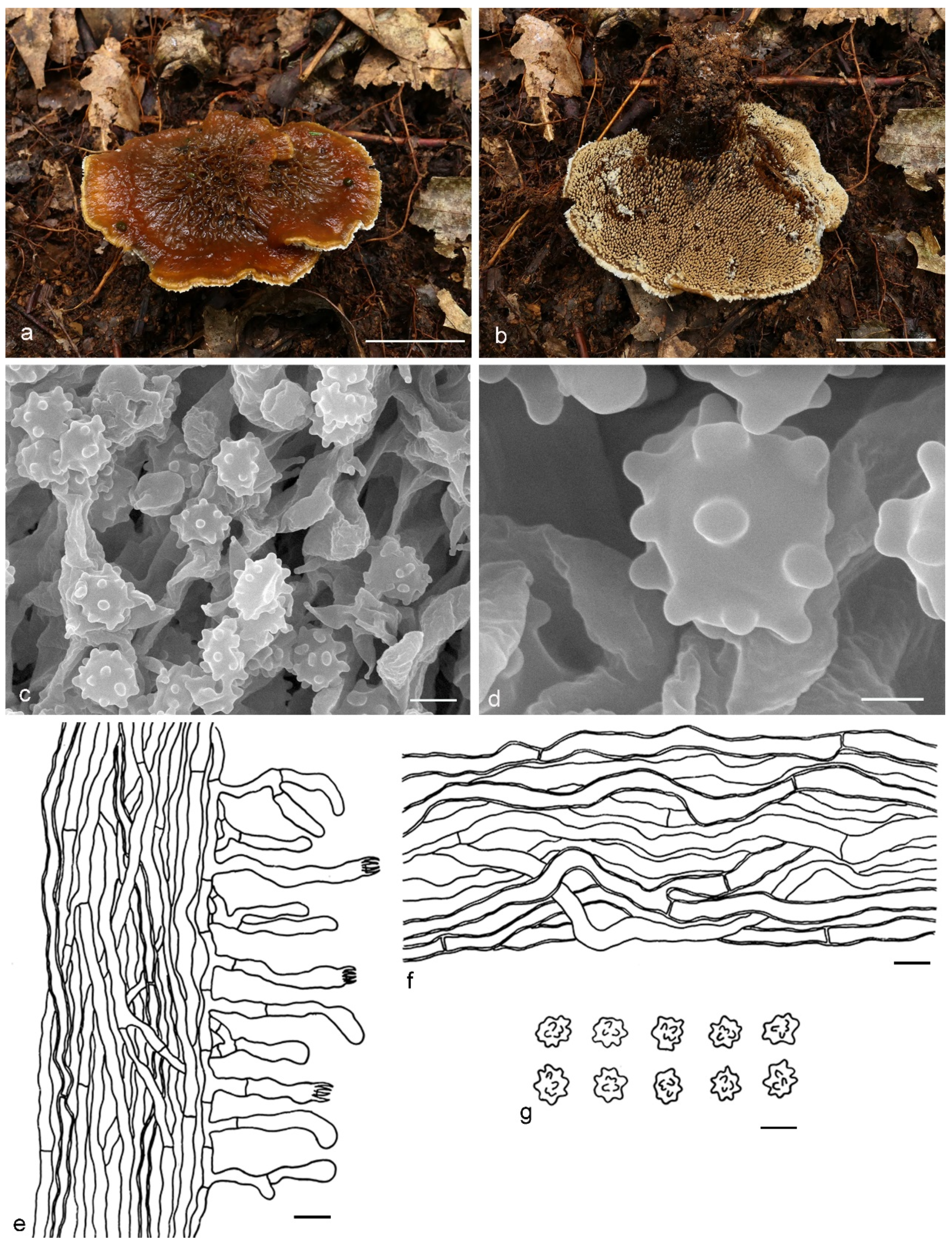
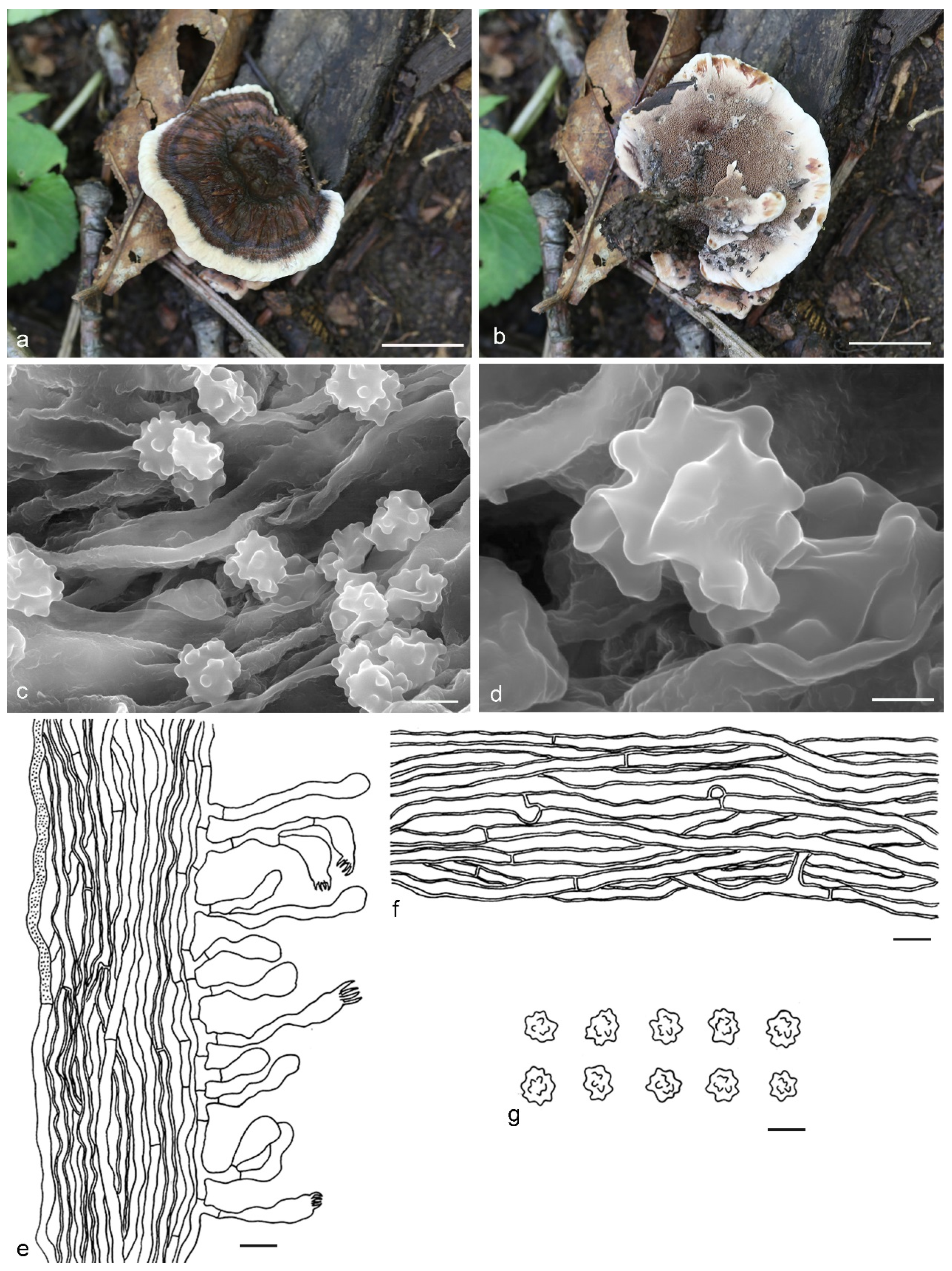
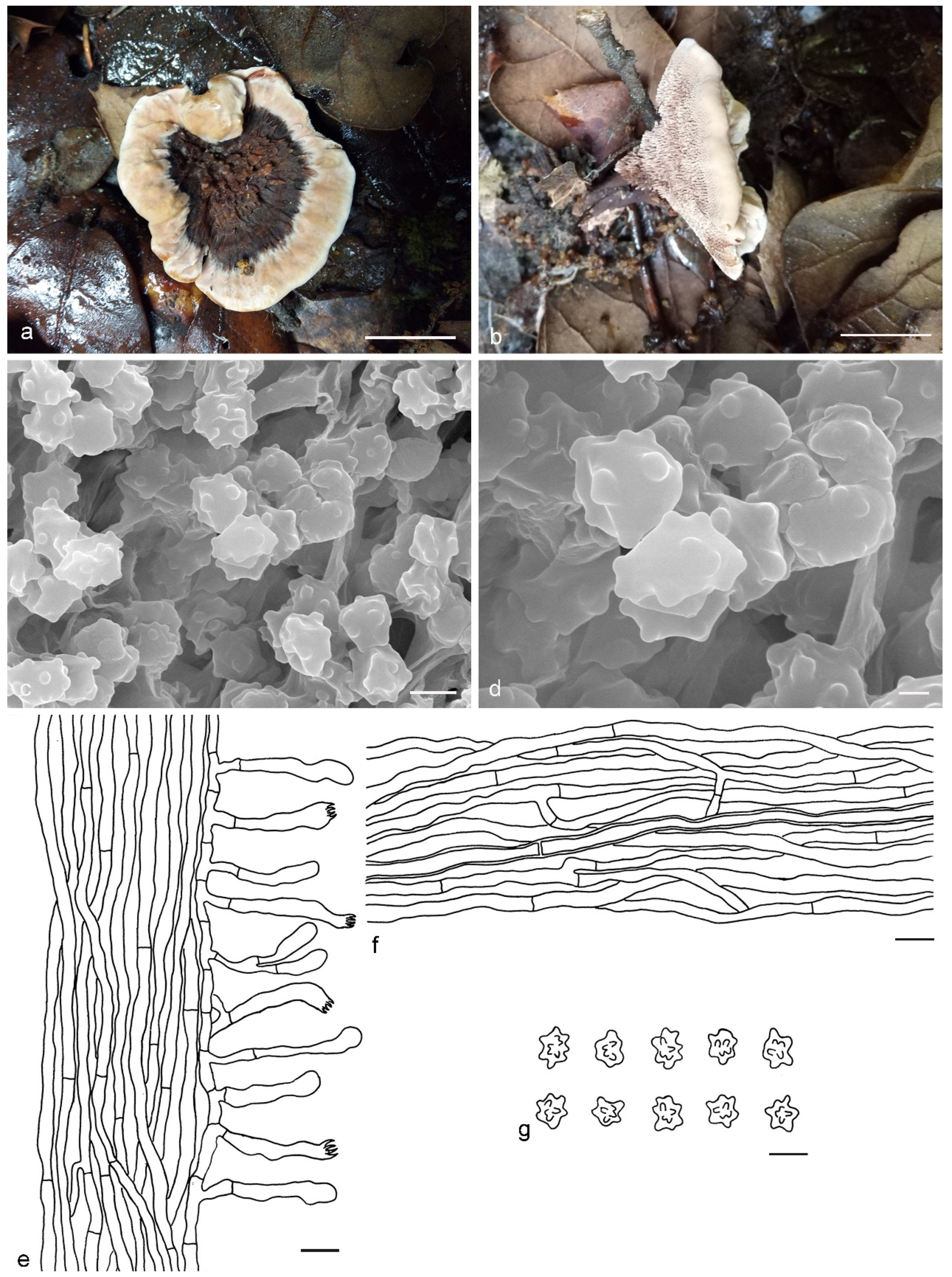
4. Taxonomy
| Key to species of Hydnellum from China | |
| 1. Basidiocarps fleshy | 2 |
| 1.→ Basidiocarps woody | 4 |
| 2. Pileal surface not scaled | H. coactum |
| 2.→ Pileal surface scaled | 3 |
| 3. Pileal surface with ascend squama | H. grosselepidotum |
| 3.→ Pileal surface with appressed squama | H. lidongensis |
| 4. Context tissue olivaceous in KOH | 5 |
| 4.→ Context tissue blue-green in KOH | H. peckii |
| 5. Hyphae with clamp-connections in the context | 6 |
| 5.→ Hyphae without clamp-connections in the context and spines | 8 |
| 6. Basidiocarps with dark violet spines underneath pileus | H. atrospinosum |
| 6.→ Basidiocarps with different colored spines underneath pileus | 7 |
| 7. Pileal surface fibrillose, rugose when dry, spines up to 1.5 mm long | H. fibulatum |
| 7.→ Pileal surface pitted, colliculose when dry, spines up to 6 mm long | H. caeruleum |
| 8. Inflated hyphae present from the context | 9 |
| 8.→ Inflated hyphae absent in the context | 10 |
| 9. Generative hyphae mostly inflated in the context, pileal surface scrobiculate when dry | H. inflatum |
| 9.→ Generative hyphae occasionally inflated in the context, pileal surface granulose when dry | H. granulosum |
| 10. Stipe thin, rhizomorphs-like | Hydnellum sp 1 |
| 10.→ Stipe cylindrical to flattened | 11 |
| 11. Pileal surface colored brownish orange to brownish red | H. brunneorubrum |
| 11.→ Pileal surface differently colored | 12 |
| 12. Pileal margin involute and wavy, sometimes lobed or rimose | 13 |
| 12.→ Pileal margin even or effused, sometimes lobed or eroded | 14 |
| 13. Pileal surface glabrescent | Hydnellum sp 4 |
| 13.→ Pileal surface not glabrescent | 15 |
| 14. Pileal surface azonate and spongy | 16 |
| 14.→ Pileal surface obsurely concentrically zonate to zonate and not spongy | 17 |
| 15. Pileal surface floccose to squamulose when fresh, context corky | H. squamulosum |
| 15.→ Pileal surface tomentose and scrupose when fresh, context woody | H. bomiense |
| 16. Basidiocarps single to gregarious and stipe single and long | H. spongiosipes |
| 16.→ Basidiocarps coalescent and stipe connate and short | Hydnellum sp 2 |
| 17. Stipe context with a dark line at centre | 18 |
| 17.→ Stipe context without a dark line at centre | 19 |
| 18. Pileus and spines grayish red | H. yunnanense |
| 18.→ Pileus brown and spines reddish brown | Hydnellum sp 3 |
| 19. Spines up to 3 or 3.5 mm long | 20 |
| 19.→ Spines up to 1 or 1.5 mm long | 21 |
| 20. Pileus light brown to dark ruby | H. atrorubrum |
| 20.→ Pileus reddish brown | H. rubidofuscum |
| 21. Basidiocarps gregarious or multiple pilei overlapping and pileal surface often grooved, scabrous to fibrous | H. sulcatum |
| 21.→ Basidiocarps solitary and pileal surface velutinous and strigose | Hydnellum sp 5 |
5. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maas Geesteranus, R.A. Die terrestrischen Stachelpilze Europas. Verh. K. Ned. Akad. van Wet. Afd. Nat. Tweede Reeks 1975, 65, 1–127. [Google Scholar]
- Baird, R.E.; Khan, S.R. The stipitate hydnums (Thelephoraceae) of Florida. Brittonia 1986, 38, 171–184. [Google Scholar] [CrossRef]
- Agerer, R. Ectomycorrhizae of Hydnellum peckii on Norway spruce and their chlamydospores. Mycologia 1993, 85, 74–83. [Google Scholar] [CrossRef]
- Stalpers, J.A. The aphyllophoraceous fungi I. Keys to the species of the Thelephorales. Stud. Mycol. 1993, 35, 1–168. [Google Scholar]
- Pegler, D.N.; Roberts, P.J.; Spooner, B.M. British Chanterelles and Tooth Fungi; Royal Botanic Gardens: Kew, UK, 1997; p. 114. [Google Scholar]
- Arnolds, E. De Stekelzwammen en Pruikzwammen van Nederland en Belgie. Coolia 2003, 46, 1–96. [Google Scholar]
- Arnolds, E. The fate of hydnoid fungi in the Netherlands and northwestern Europe. Fungal Ecol. 2010, 3, 81–88. [Google Scholar] [CrossRef]
- Arnolds, E. E. Former and present distribution of stipitate hydnaceous fungi (Basidiomycetes) in the Netherlands. Nova Hedwig. 1989, 48, 107–142. [Google Scholar]
- Bergelson, J.M.; Crawley, M.J. Mycorrhizal infection and species diversity. Nature 1998, 334, 202. [Google Scholar] [CrossRef]
- Van der Heijden, M.; Klironomos, J.; Ursic, M.; Moutoglis, P.; Streitwolf-Engel, R.; Boller, T.; Wiemken, A.; Sanders, I. Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature 1998, 396, 69–72. [Google Scholar] [CrossRef]
- Wu, F.; Zhou, L.W.; Yang, Z.L.; Bau, T.; Li, T.H.; Dai, Y.C. Resource diversity of Chinese macrofungi: Edible, medicinal and poisonous species. Fungal Divers. 2019, 98, 1–76. [Google Scholar] [CrossRef]
- Lee, D.; Boo, K.H.; Lee, J.M.; Viet, C.D.; Quyen, N.; Unno, T.; Cho, M.; Riu, K.Z.; Lee, D.S. Anti-viral activity of Hydnellum concrescens, a medicinal mushroom. Afr. J. Biotechnol. 2012, 11, 15241–15245. [Google Scholar] [CrossRef]
- Farzaneh, V.; Carvalho, I.S. A review of the health benefit potentials of herbal plant infusions and their mechanism of actions. Ind. Crop. Prod. 2015, 65, 247–258. [Google Scholar] [CrossRef]
- Lizon, P. Decline of macrofungi in Europe: An overview. Trans. Mycol. Soc. Repub. China 1993, 8, 21–48. [Google Scholar]
- Newton, A.C.; Holden, E.; Davy, L.M.; Ward, S.D.; Fleming, L.V.; Watling, R. Status and distribution of stipitate hydnoid fungi in Scottish coniferous forests. Biol. Conserv. 2002, 107, 181–192. [Google Scholar] [CrossRef]
- Van der Linde, S.; Alexander, I.J.; Anderson, I.C. A PCR-based method for detecting the mycelia of stipitate hydnoid fungi in soil. J. Microbiol. Methods 2008, 75, 40–46. [Google Scholar] [CrossRef]
- Jansen, E.J.; Van Dobben, H.F. Is decline of Cantharellus cibarius in the Netherlands due to air pollution? Ambio 1987, 16, 211–213. [Google Scholar]
- Arnolds, E. A preliminary Red Data List of macrofungi in the Netherlands. Persoonia 1989, 14, 77–125. [Google Scholar]
- Arnolds, E. Decline of ectomycorrhizal fungi in Europe. Agric. Ecosyst. Environ. 1991, 35, 209–244. [Google Scholar] [CrossRef]
- Termorshuizen, A.J.; Schaffers, A.P. The decline of sporocarps of ectomycorrhizal fungi in stands of Pinus sylvestris L. in The Netherlands: Possible causes. Nova Hedwig. 1991, 53, 267–289. [Google Scholar]
- Wallenda, T.; Kottke, I. Nitrogen deposition and ectomycorrhizas. New Phytol. 1998, 139, 169–187. [Google Scholar] [CrossRef]
- Lizon, P. Macrofungi reported as extinct or threatened with extinction in European Red Data Lists. Fungi Conserv. Newsl. 1995, 3, 3–4. [Google Scholar]
- Senn-Irlet, B.; Bieri, G.; Egli, S. Rote Liste Grosspilze. Rote Liste der Gefährdeten Arten der Schweiz; BAFU: Bern, Switzerland, 2007; Volume 18, p. 92. [Google Scholar]
- Smith, J.H.; Suz, L.M.; Ainsworth, A.M.; Smith, J.H.; Suz, L.M.; Ainsworth, A.M. Red List of Fungi for Great Britain: Bankeraceae, Cantharellaceae, Geastraceae, Hericiaceae and Selected Genera of Agaricaceae (Battarrea, Bovista, Lycoperdon & Tulostoma) and Fomitopsidaceae (Piptoporus); Jodrell Laboratory, Royal Botanic Gardens: Kew, UK, 2016; p. 90. [Google Scholar]
- Anon. UK Biodiversity Group. Tranche 2 Actions Plans—Volume III: Plants and Fungi; English Nature: Peterborough, UK, 1999; pp. 24–27. [Google Scholar]
- Maas Geesteranus, R.A. Hydnaceous fungi of the eastern old world. Vehr. K. Ned. Akad. van Wet. Afd. Nat. 1971, 60, 1–176. [Google Scholar]
- Baird, R.; Wallace, L.E.; Baker, G.; Scruggs, M. Stipitate hydnoid fungi of the temperate southeastern United States. Fungal Divers. 2013, 62, 41–114. [Google Scholar] [CrossRef]
- Binder, M.; Hibbett, D.S.; Larsson, K.H.; Larsson, E.; Langer, E.; Langer, G. The phylogenetic distribution of resupinate forms across the major clades of mushroom-forming fungi (Homobasidiomycetes). Syst. Biodivers. 2005, 3, 113–157. [Google Scholar] [CrossRef]
- Matheny, P.B.; Curtis, J.M.; Hofstetter, V.; Aime, M.C. Major clades of Agaricales: A multilocus phylogenetic overview. Mycologia 2006, 98, 982–995. [Google Scholar] [CrossRef]
- Tedersoo, L.; May, T.W.; Smith, M.E. Ectomycorrhizal lifestyle in fungi: Global diversity, distribution, and evolution of phylogenetic lineages. Mycorrhiza 2010, 20, 217–263. [Google Scholar] [CrossRef]
- Larsson, K.H.; Svantesson, S.; Miscevic, D.; Kljalg, U.; Larsson, E. Reassessment of the generic limits for Hydnellum and Sarcodon (thelephorales, basidiomycota). MycoKeys 2019, 54, 31–47. [Google Scholar] [CrossRef]
- Banker, H.J. A contribution to a revision of the North American Hydnaceae. Mem. Torrey Bot. Club 1906, 12, 99–194. [Google Scholar]
- Harrison, K.A. New or little known North American stipitate hydnums. Can. J. Bot. 1964, 42, 1205–1233. [Google Scholar] [CrossRef]
- Baird, R.E. Study of the stipitate hydnums from the southern Appalachian Mountains—Genera: Bankera, Hydnellum, Phellodon, Sarcodon. Bibl. Mycol. 1986, 104, 1–156. [Google Scholar]
- Koljalg, U.; Renvall, P. Hydnellum gracillipes—A link between stipitate and resupinate Hymenomycetes. Karstenia 2000, 40, 71–77. [Google Scholar] [CrossRef][Green Version]
- Loizides, M.; Alvarado, P.; Assyov, B.; Arnolds, E.; Moreau, P.A. Hydnellum dianthifolium sp nov. (Basidiomycota, Thelephorales), a new tooth-fungus from southern Europe with notes on H. concrescens and H. scrobiculatum. Phytotaxa 2016, 280, 23–35. [Google Scholar] [CrossRef]
- Dai, Y.C. A revised checklist of corticioid and hydnoid fungi in China for 2010. Mycoscience 2011, 52, 69–79. [Google Scholar] [CrossRef]
- Mu, Y.H.; Hu, Y.P.; Wei, Y.L.; Yuan, H.S. Hydnaceous fungi of China 8. morphological and molecular identification of three new species of Sarcodon and a new record from southwest China. MycoKeys 2020, 66, 83–103. [Google Scholar] [CrossRef]
- Rayner, R.W. A Mycological Colour Chart; Commonweath Mycological Institute and British Mycological Society: Kew, UK, 1970. [Google Scholar]
- Munsell, A.H. Munsell Soil-Color Charts with Genuine Munsell Color Chips; Munsell Color: Grand Rapids, MI, USA, 2015. [Google Scholar]
- Stöger, A.; Schaffer, J.; Ruppitsch, W. A rapid and sensitive method for direct detection of Erwinia amylovora in symptomatic and asymptomatic plant tissues by polymerase chain reaction. J. Phytopathol. 2006, 154, 469–473. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap alogarithm for the RAxML webservers. Syst. Biol. 2008, 75, 758–771. [Google Scholar] [CrossRef] [PubMed]
- Cannatella, D. Xenopus in space and time: Fossils, node calibrations, tip-dating, and paleobiogeography. Cytogenet. Genome Res. 2015, 145, 283–301. [Google Scholar] [CrossRef] [PubMed]
- Posada, D.; Crandall, K.A. MODELTEST: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [PubMed]
- Nylander, J. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- White, T.; Bruns, T.; Lee, S.; Taylor, F.; White, T.; Lee, S.H.; Taylor, L.; Shawetaylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Baird, R.E. Type studies of North American and other related taxa of stipitate hydnums: Genera Bankera, Hydnellum, Phellodon, Sarcodon. Bibl. Mycol. 1986, 103, 1–89. [Google Scholar]
- Rubio Casas, L.; Rubio Roldán, L.; Català, S. Sarcodon amygdaliolens, a new species of Sarcodon Quél. ex P. Karst. found in the Iberian Peninsula. Boletín Soc. Micológica Madr. 2011, 35, 43–56. [Google Scholar]
- Nitare, J.; Ainsworth, A.M.; Larsson, E.; Parfitt, D.; Suz, L.M.; Svantesson, S.; Larsson, K.H. Four new species of Hydnellum (Thelephorales, Basidiomycota) with a note on Sarcodon illudens. Fungal Syst. Evol. 2021, 7, 233–254. [Google Scholar] [CrossRef]
- Hahn, C.; Friebes, G.; Krisai-Greilhuber, I. Sarcodon fennicus, a boreo-montane stipitate hydnoid fungus with a remarkable smell. Austrian Mycol. Soc. 2018, 27, 43–52. [Google Scholar]
- Otto, P. Die Terrestrischen Stachelpilze der DDR—Taxonomie, Ökologie, Verbreitung und Rückgang. Doctoral Dissertation, Martin-Luther-Universität, Halle-Wittenberg, 1990. [Google Scholar]
- Hrouda, P. Hydnaceous fungi of the Czech Republic and Slovakia. Czech Mycol. 1999, 51, 99–155. [Google Scholar] [CrossRef]
- Van der Linde, S.; Alexander, I.J.; Anderson, I.C. Spatial distribution of sporocarps of stipitate hydnoid fungi and their belowground mycelium. FEMS Microbiol. Ecol. 2009, 69, 344–352. [Google Scholar] [CrossRef][Green Version]
- Thompson, G.W.; Medwe, R.J. Effects of aluminimum and manganese on the growth of ectomycorrhizal fungi. Appl. Environ. Microbiol. 1984, 48, 556–560. [Google Scholar] [CrossRef]
- Jahn, H. Der “Satanspilzhang” bei Glesse (Ottenstein), Siid-Niedersachsen. Westffilische Pilzbr. 1986, 10–11, 289–351. [Google Scholar]
- Termorshuizen, A.J.; Schaffers, A.P. Occurrence of carpophores of mycorrhizal fungi in selected stands of Pinus sylvestris in the Netherlands in relation to stand vitality and air pollution. Plant Soil 1987, 104, 209–217. [Google Scholar] [CrossRef]
- Arnolds, E. The changing macromycete flora in the Netherlands. Trans. Br. Mycol. Soc. 1988, 90, 391–406. [Google Scholar] [CrossRef]
- Hrouda, P. Bankeraceae in central Europe. 1. Czech Mycol. 2005, 57, 57–78. [Google Scholar] [CrossRef]
- Hrouda, P. Bankeraceae in central Europe. 2. Czech Mycol. 2005, 57, 279–297. [Google Scholar] [CrossRef]
- Arnolds, E.; Veerkamp, M. Basisrapport Rode Lijst Paddenstoelen; Ministry of Agriculture, Nature and Food Safety: The Hague, The Netherlands, 2008. [Google Scholar]
- Maas Geesteranus, R.A. Notes on Hydnums–VII. Persoonia 1967, 5, 1–13. [Google Scholar]
- Mleczko, P.; Zubek, S.; Kozak, M. Description of ectomycorrhiza and a new Central European locality of the rare hydnoid species Sarcodon leucopus (Pers.) Maas Geest. et Nannf. (Thelephorales, Basidiomycota). Nova Hedwig. 2011, 92, 257–272. [Google Scholar] [CrossRef]
- Pérez-De-Gregorio, M.A.; Macau, N.; Carbó, J. Sarcodon quercinofibulatum, una nueva especie del género con hifas fibulíferas. Rev. Catalana Micol. 2011, 33, 25–30. [Google Scholar]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
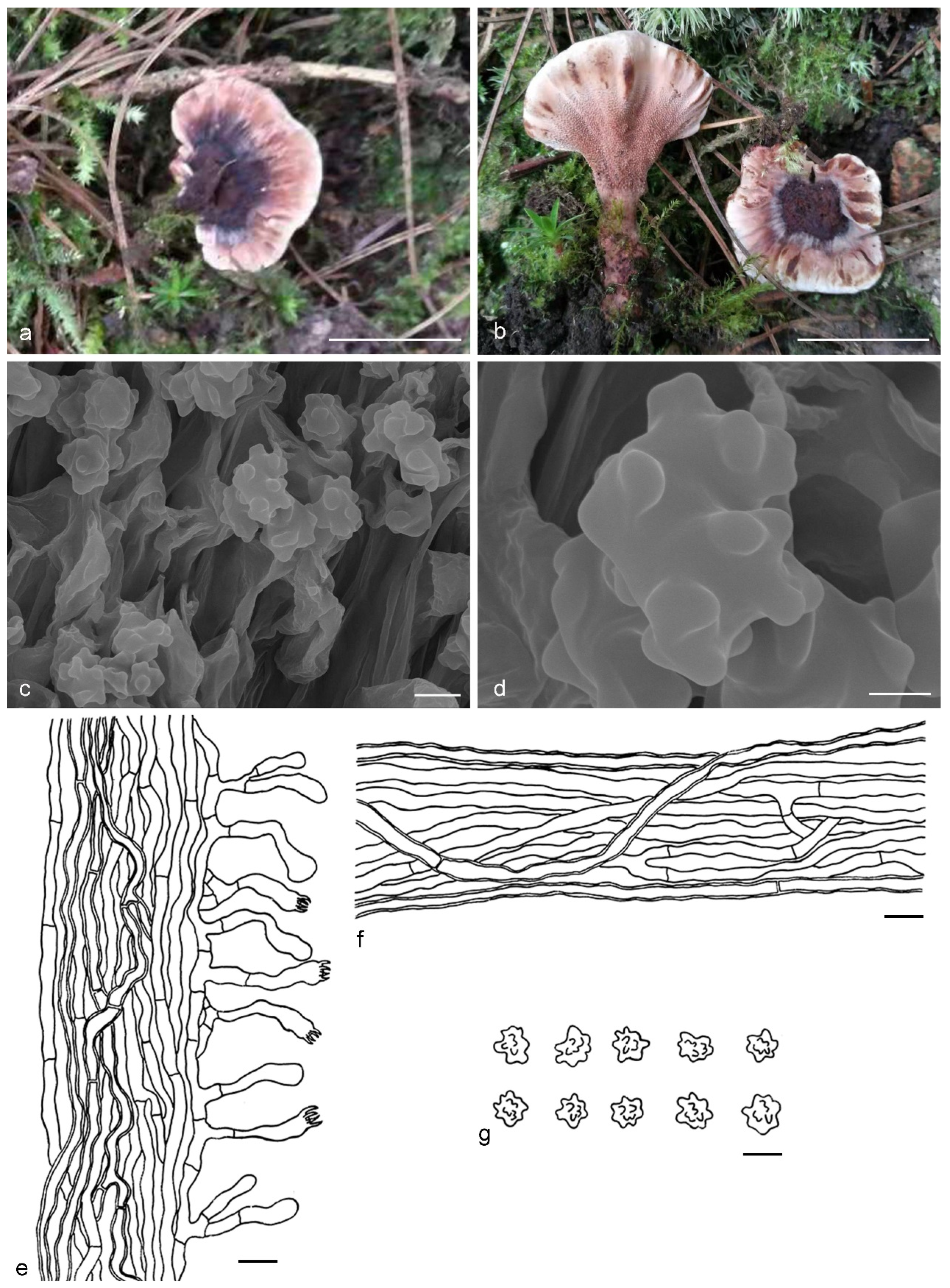{kind=link}
| Species | Geographic Origin | Voucher Number | GenBank Accessions No. | |||
|---|---|---|---|---|---|---|
| SSU | ITS | nLSU | RPB2 | |||
| Amaurodon aquicoeruleu Agerer | Australia | UK452 | – | AM490944 | AM490944 | – |
| A. sumatranus Miettinen & Kõljalg | Indonesia | O. Miettinen5877 | – | AM490943 | – | – |
| A. viridis (Alb. & Schwein.) J. Schröt. | Norway | KHLarsson14947b | – | MK602707 | MK602707 | – |
| A. viridis | Russia | TAA149664 | – | AM490942 | AY586625 | – |
| Bankera fuligineoalba (J.C. Schmidt) Coker & Beers ex Pouzar | Sweden | ELarsson400-13 | – | MK602708 | MK602708 | – |
| B. fuligineoalba | Estonia | TAA152454 | – | – | AY586635 | – |
| B. violascens (Alb. & Schwein.) Pouzar | Finland | MVijanen130902 | – | MK602709 | MK602709 | – |
| B. violascens | – | RGC14-033 | – | MH310793 | – | – |
| Boletopsis grisea (Peck) Bondartsev & Singer | Sweden | UPS F-120382 | – | MN536751 | MN535646 | – |
| B. grisea | Spain | AH 42971 | – | MN536747 | MN535642 | – |
| B. leucomelaena (Pers.) Fayod | Sweden | Krikorev140912 | – | MK602710 | MK602710 | – |
| B. nothofagi J.A. Cooper & P. Leonard | New Zealand | PDD:96007 | – | JQ417193 | – | – |
| Hydnellum amygdaliolens (Rubio Casas, Rubio Roldán & Català) E. Larss., K.H. Larss. & Kõljalg | Iberian Peninsula | SC-2011 | – | JN376763 | – | – |
| H. atrorubrum | China | Wei8315 | – | MW579937 | – | – |
| H. atrorubrum | China | Wei8261 | MW579910 | MW579936 | MW579884 | – |
| H. atrospinosum | China | Yuan6495 | MW579911 | MW579938 | MW579885 | – |
| H. atrospinosum | China | Yuan6514 | MW579913 | MW579940 | MW579886 | – |
| H. atrospinosum | China | Yuan6520 | MW579912 | MW579939 | – | – |
| H. aurantiacum (Batsch) P. Karst. | Norway | EBendiksen177-07 | – | MK602712 | MK602712 | – |
| H. aurantiacum | Norway | OF29502 | – | MK602713 | MK602713 | – |
| H. auratile (Britzelm.) Maas Geest. | Norway | OF242763 | – | MK602715 | MK602715 | – |
| H. auratile | Norway | OF294095 | – | MK602714 | MK602714 | – |
| H. bomiense | China | Yuan 13759 | MW579914 | MW579941 | MW579887 | OK254206 |
| H. bomiense | China | Yuan 13767 | MW579915 | MW579942 | – | – |
| H. bomiense | Estonia | TUF100611 | – | UDB003287 | – | – |
| H. bomiense | Costa Rica | TUF100057 | – | UDB003286 | – | – |
| H. brunneorubrum | China | Yuan12997 | MW579917 | MW579944 | MW579889 | OK254217 |
| H. brunneorubrum | China | Yuan14339 | MW579916 | MW579943 | MW579888 | OK254216 |
| H. brunneorubrum | China | Yuan14668 | MW579918 | MW579945 | MW579890 | OK254218 |
| H. caeruleum (Hornem.) P. Karst. | Norway | EBendiksen584-11 | – | MK602719 | MK602719 | – |
| H. caeruleum | Norway | EBendiksen575-11 | – | MK602718 | MK602718 | – |
| H. caeruleum | China | Wei1474a | – | MW579965 | – | – |
| H. chrysinum K.A. Harrison | – | SC071 | – | KJ534291 | – | – |
| H. coactum Y.H. Mu & H.S. Yuan | China | Wei8094 | – | MN846278 | MN846287 | – |
| H. coactum | China | Shi181 | – | MN846279 | MN846288 | – |
| H. complicatum Banker | USA | REB-71 | – | KC571711 | – | – |
| H. complicatum | USA | REB-329 | – | KC571712 | – | – |
| H. concrescens (Pers.) Banker | USA | SEW 88 | – | AY569025 | – | – |
| H. concrescens | Mexico | GO-2009-204 | – | KC152116 | – | – |
| H. cristatum (Bres.) Stalpers | USA | REB-169 | – | JN135174 | – | – |
| H. cristatum | USA | REB-88 | – | KC571718 | – | – |
| H. cumulatum K.A. Harrison | Finland | TU115384 | – | UDB011871 | UDB011871 | – |
| H. cumulatum | Estonia | TU111191 | – | UDB032402 | – | – |
| H. cyanopodium K.A. Harrison | USA | SEW 85 | – | AY569027 | – | – |
| H. diabolus Banker | Canada | KAH13873 | – | AF351863 | – | – |
| H. dianthifolium Loizides | Cyprus | ML61211HY | – | KX619419 | – | – |
| H. dianthifolium | Italy | ML902162HY | – | KX619420 | – | – |
| H. earlianum Banker | USA | REB-75 | – | KC571724 | – | – |
| H. earlianum | USA | REB-375 | – | JN135179 | – | – |
| H. fagiscabrosum A.M. Ainsw. & Nitare | Sweden | GB-0195621 | – | MW144293 | MW144293 | – |
| H. fagiscabrosum | Sweden | GB-0195622 | – | MW144296 | MW144296 | – |
| H. fennicum (P. Karst.) E. Larss, K.H. Larss. & Kõljalg | Norway | OF242833 | – | MK602738 | MK602738 | – |
| H. fennicum | Norway | OF294087 | – | MK602737 | MK602737 | – |
| H. ferrugineum (Fr.) P. Karst. | Norway | OF297319 | – | MK602720 | MK602720 | – |
| H. ferrugineum | Sweden | ELarsson197-14 | – | MK602722 | MK602722 | – |
| H. ferrugipes Coker | USA | REB-176 | – | KC571727 | – | – |
| H. ferrugipes | USA | REB-68 | – | JN135176 | – | – |
| H. fibulatum | China | Yuan14646 | MW579926 | MW579957 | – | – |
| H. fibulatum | China | Yuan14656 | MW579927 | MW579958 | – | – |
| H. fuligineoviolaceum (Kalchbr.) E. Larss., K.H. Larss. & Kõljalg | Sweden | BNylen130918 | – | MK602741 | MK602741 | – |
| H. fuligineoviolaceum | Sweden | LA120818 | – | MK602740 | MK602740 | – |
| H. fuscoindicum (K.A. Harrison) E. Larss., K.H. Larss. & Kõljalg | USA | OSC 113641 | – | EU669230 | EU669280 | – |
| H. fuscoindicum | USA | OSC 107844 | – | EU669229 | EU669279 | – |
| H. glaucopus (Maas Geest. & Nannf.) E. Larss., K.H. Larss. & Kõljalg | Sweden | JNitare060916 | – | MK602744 | MK602744 | – |
| H. glaucopus | Sweden | Edvinson110926 | – | MK602745 | MK602745 | – |
| H. geogenium (Fr.) Banker | – | AFTOL-ID 680 | AY752971 | DQ218304 | AY631900 | DQ408133 |
| H. geogenium | Norway | OF296213 | – | MK602724 | MK602724 | – |
| H. gracilipes (P. Karst.) P. Karst. | Sweden | ELarsson219-11 | – | MK602726 | MK602726 | – |
| H. gracilipes | Sweden | GB-0113779 | – | MK602727 | MK602727 | – |
| H. granulosum | China | Yuan12213a | MW579921 | MW579948 | MW579893 | OK254213 |
| H. granulosum | China | Yuan12213b | MW579920 | MW579947 | MW579892 | OK254212 |
| H. grosselepidotum Y.H. Mu & H.S. Yuan | China | Wei8120 | – | MN846274 | MN846283 | – |
| H. grosselepidotum | China | Wei8075 | – | MN846276 | MN846285 | – |
| H. illudens (Maas Geest.) Nitare | Sweden | GB-0195819 | – | MW144341 | MW144341 | – |
| H. illudens | Norway | O-F-242769 | – | MW144335 | MW144335 | – |
| H. inflatum | China | Wang80 | MW579922 | MW579949 | MW579894 | OK254210 |
| H. inflatum | China | Shi506 | MW579923 | MW579950 | MW579895 | OK254211 |
| H. joeides (Pass.) E. Larss., K.H. Larss. & Kõljalg | Sweden | KHjortstam17589 | – | MK602750 | MK602750 | – |
| H. joeides | Sweden | Nitare110829 | – | MK602751 | MK602751 | – |
| H. lepidum (Maas Geest.) E. Larss., K.H. Larss. & Kõljalg | Sweden | JNitare110829 | – | MK602754 | MK602754 | – |
| H. lepidum | Sweden | RGCarlsson10-065 | – | MK602752 | MK602752 | – |
| H. lidongensis Y.H. Mu & H.S. Yuan | China | We8365 | – | MN846280 | MN846289 | – |
| H. lidongensis | China | Wei8329 | – | MN846281 | MN846290 | – |
| H. lundellii (Maas Geest. & Nannf.) E. Larss., K.H. Larss. & Kõljalg | Norway | OF242639 | – | MK602759 | MK602759 | – |
| H. lundellii | Norway | OF295814 | – | MK602760 | MK602760 | – |
| H. martioflavum (Snell, K.A. Harrison & H.A.C. Jacks.) E. Larss., K.H. Larss. & Kõljalg | Norway | OF242435 | – | MK602762 | MK602762 | – |
| H. martioflavum | Norway | OF242872 | – | MK602761 | MK602761 | – |
| H. mirabile (Fr.) P. Karst. | Sweden | SLund140912 | – | MK602730 | MK602730 | – |
| H. mirabile | Sweden | ELarsson170-14 | – | MK602729 | MK602729 | – |
| H. nemorosum A.M. Ainsw. & E. Larss. | Norway | O-F-242352 | – | MW144372 | MW144372 | – |
| H. nemorosum | Sweden | GB-0195631 | – | MW144373 | MW144373 | – |
| H. parvum Banker | USA | REB-131 | – | JN135187 | – | – |
| H. parvum | USA | REB-392 | – | KC571717 | – | – |
| H. peckii Banker | Norway | SSvantesson328 | – | MK602731 | MK602731 | – |
| H. peckii | Sweden | ELarsson174-14 | – | MK602732 | MK602732 | – |
| H. peckii | China | Yuan13708 | MW579931 | MW579966 | MW579905 | OK254214 |
| H. peckii | China | Yuan13720 | MW579932 | MW579967 | MW579906 | OK254215 |
| H. pineticola K.A. Harrison | USA | REB-49 | – | KC571733 | – | – |
| H. pineticola | USA | REB-43 | – | JN135175 | – | – |
| H. piperatum Coker ex Maas Geest. | USA | REB-332 | – | JN135173 | – | – |
| H. piperatum | USA | REB-304 | – | KC571723 | – | – |
| H. regium K.A. Harrison | USA | SEW 93 | – | AY569031 | – | – |
| H. roseoviolaceum Nitare | Sweden | GB-0195936 | – | MW144374 | MW144374 | – |
| H. roseoviolaceum | Sweden | GB-0195687 | – | MW144375 | MW144375 | – |
| H. rubidofuscum | China | Yuan14561 | MW579924 | MW579951 | MW579896 | OK254207 |
| H. rubidofuscum | China | Yuan14587 | MW579925 | MW579952 | MW579897 | OK254208 |
| H. rubidofuscum | China | Yuan14654 | – | MW579953 | MW579898 | OK254209 |
| H. scabrosum (Fr.) E. Larss., K.H. Larss. & Kõljalg | Norway | OF360777 | – | MK602765 | MK602765 | |
| H. scabrosum | Norway | OF292320 | – | MK602766 | MK602766 | |
| H. scabrosellum Nitare | Sweden | GB-0195792 | – | MW144380 | MW144380 | – |
| H. scabrosellum | Sweden | GB-0195807 | – | MW144381 | MW144381 | – |
| H. scleropodium K.A. Harrison | USA | REB-3 | – | JN135186 | – | – |
| H. scleropodium | USA | REB-352 | – | KC571740 | – | – |
| H. scrobiculatum (Fr.) P. Karst. | USA | REB-78 | – | JN135181 | – | – |
| H. spongiosipes (Peck) Pouzar | USA | REB-107 | – | KC571743 | – | – |
| H. spongiosipes | USA | REB-52 | – | JN135184 | – | – |
| H. spongiosipes | China | Yuan14517 | MW579933 | MW579968 | MW579907 | OK254219 |
| H. squamulosum | China | Yuan13615 | – | MW579954 | – | – |
| H. squamulosum | China | Yuan13625 | – | MW579956 | MW579899 | OK254204 |
| H. squamulosum | China | Yuan13743 | – | MW579955 | – | OK254203 |
| H. suaveolens (Scop.) P. Karst. | Sweden | ELarsson8-14 | – | MK602735 | MK602735 | – |
| H. suaveolens | Norway | SSvantesson877 | – | MK602736 | MK602736 | – |
| H. subsuccosum K.A. Harrison | USA | SEW 55 | – | AY569033 | – | – |
| H. subsuccosum | USA | REB-10 | – | JN135178 | – | – |
| H. sulcatum | China | Yuan14521 | MW579930 | MW579961 | MW579902 | OK254202 |
| H. sulcatum | China | Yuan14649 | MW579929 | MW579960 | MW579901 | – |
| H. sulcatum | China | Yuan14660 | MW579928 | MW579959 | MW579900 | OK254201 |
| H. yunnanense | China | Yuan14386 | – | MW579962 | MW579903 | OK254199 |
| H. yunnanense | China | Yuan14396 | – | MW579963 | MW579904 | OK254200 |
| H. yunnanense | China | Shi212 | – | MW579964 | – | – |
| H. underwoodii (Banker) E. Larss., K.H. Larss. & Kõljalg | USA | REB-358 | – | JN135189 | – | – |
| H. underwoodii | USA | REB-119 | – | KC571782 | – | – |
| H. versipelle (Fr.) E. Larss., K.H. Larss. & Kõljalg | Sweden | RGCarlsson13-057 | – | MK602771 | MK602771 | – |
| H. versipelle | Sweden | RGCarlsson11-08 | – | MK602772 | MK602772 | – |
| Hydnellum sp 1 | China | Shi164 | – | MW579969 | – | – |
| Hydnellum sp 2 | China | Yuan14387 | MW579934 | MW579970 | MW579908 | – |
| Hydnellum sp 3 | China | Yuan14388 | – | MW579971 | – | – |
| Hydnellum sp 4 | China | Wang295 | – | MW579972 | – | – |
| Hydnellum sp 5 | China | Yuan14594 | MW579935 | MW579973 | MW579909 | OK254205 |
| Lenzitopsis daii L.W. Zhou & Kõljalg | China | Yuan 2959 | – | JN169799 | JN169795 | – |
| L. daii | China | Yuan2952 | – | JN169798 | JN169794 | – |
| L. oxycedri Malençon & Bertault | Spain | KHLarsson15304 | – | MK602774 | MK602774 | – |
| L. oxycedri | – | UK 635 | – | JN169800 | JN169796 | – |
| Odontia fibrosa (Berk. & M.A. Curtis) Kõljalg | China | TU115028 | – | MK602775 | MK602775 | |
| O. fibrosa | China | LL_17 | – | MT678878 | – | – |
| O. sparsa Yuan, Y.C. Dai & H.S. Yuan | China | Yuan10718 | – | MG719980 | – | – |
| O. sparsa | China | Yuan10780 | – | MG719979 | – | – |
| Phellodon cf. niger | Sweden | ELarsson35-14 | – | MK602782 | MK602782 | – |
| P. tomentosus (L.) Banker | Norway | EBendiksen11-810 | – | MK602781 | MK602781 | – |
| P. tomentosus | – | BG Thesis | – | – | AF518637 | – |
| Polyozellus mariae Voitk & Kõljalg | Canada | TU117348 | – | MF100831 | MF100831 | – |
| P. mariae | Canada | TU117235 | – | MF100826 | – | – |
| P. multiplex (Underw.) Murrill | USA | TU117350 | – | MF100830 | MF100830 | – |
| P. multiplex | China | TU115049 | – | MF100812 | MF100812 | – |
| Pseudotomentella abundiloba Svantesson | Norway | OF110312 | MK290731 | MK290731 | ||
| P. flavovirens (Höhn. & Litsch.) Svrček | Finland | KHLarsson16190 | – | MK602780 | MK602780 | – |
| P. rotundispora Svantesson | Sweden | SS394 | – | MK290728 | MK290728 | – |
| P. rotundispora | Sweden | SS413 | – | MK290674 | – | – |
| P. umbrinascens Svantesson | Sweden | SS335 | – | MK290697 | MK290697 | – |
| Sarcodon aspratus (Berk.) S. Ito | – | – | – | DQ448877 | – | – |
| S. aspratus | – | – | – | AF335110 | – | – |
| S. imbricatus (L.) P. Karst. | Norway | SSvantesson355 | – | MK602748 | MK602748 | – |
| S. imbricatus | Sweden | ELarsson384-10 | – | MK602747 | MK602747 | – |
| S. leucopus (Pers.) Maas Geest. & Nannf. | Norway | OF296099 | – | MK602755 | MK602755 | – |
| S. leucopus | Sweden | PHedberg080811 | – | MK602757 | MK602757 | – |
| S.quercinofibulatus Pérez-De-Greg., Macau & J. Carbó | Italy | JC-20090718.2 | – | JX271818 | MK602773 | – |
| S. quercinofibulatus | USA | TENN | – | MG663244 | – | – |
| S. scabripes (Peck) Banker | Mexico | FCME:23240 | – | EU293829 | – | – |
| S. scabripes | USA | REB-351 | – | JN135191 | – | – |
| S.squamosus (Schaeff.) P. Karst. | Norway | OF295554 | – | MK602769 | MK602769 | – |
| S. squamosus | Norway | OF177452 | – | MK602768 | MK602768 | – |
| Steccherinum murashkinskyi (Burt) Maas Geest. | Russia | X449 | – | JN710588 | JN710588 | – |
| S. ochraceum (Pers. ex J.F. Gmel.) Gray | Sweden | KHL11902 | – | JQ031130 | JQ031130 | – |
| Thelephora ganbajun M. Zang | China | GDGM 48899 | – | MF593267 | MH620355 | – |
| T. ganbajun | China | GDGM 48891 | – | MF593266 | MH620354 | – |
| T. iqbalii Nasir & Hanif | Pakistan | MH810 | – | JX241471 | – | – |
| T. terrestris Ehrh. | Denmark | DMS-9327942 | – | MT644883 | MT644883 | – |
| T. terrestris | Norway | ELarsson295-13 | – | MK602777 | MK602777 | – |
| Tomentella fuscocrustosa H.S. Yuan, X. Lu & Y.C. Dai | China | Yuan11399 | – | MK211712 | MK446366 | – |
| T. fuscocrustosa | China | Yuan11420 | – | MK211713 | MK446367 | – |
| T.patagonica Kuhar & Rajchenb. | Argentina | BAFC52372 | – | KT032090 | KT032102 | – |
| T. patagonica | Argentina | BAFC52373 | – | KT032091 | KT032103 | – |
| Tomentellopsis bresadoliana (Sacc. & Trotter) Jülich & Stalpers | Sweden | JEH 031011 | – | EU118674 | EU118674 | – |
| T. pulchella Kõljalg & Bernicchia | Norway | KHLarsson16366 | – | MK602779 | MK602779 | – |
| Genes | Primers | Primer Sequences (5’-3’) | References |
|---|---|---|---|
| nLSU | LROR | ACCCGCTGAACTTAAGC | Vilgalys & Hester 1990 [48] |
| LR7 | TACTACCACCAAGATCT | Vilgalys & Hester 1990 [48] | |
| ITS | ITS1-F | CTTGGTCATTTAGAGGAAGTAA | White et al. 1990 [49] |
| ITS4 | TCCTCCGCTTATTGATATGC | White et al. 1990 [49] | |
| nSSU | NS1 | GTAGTCATATGCTTGTCTC | White et al. 1990 [49] |
| NS4 | CTTCCGTCAATTCCTTTAAG | White et al. 1990 [49] | |
| RPB2 | bRPB2-6F | TGGGGYATGGTNTGYCCYGC | Liu et al. 1999 [50] |
| bRPB2-7.1R | CCCATRGCYTGYTTMCCCATDGC | Liu et al. 1999 [50] |
| Group | Subgenus | Species | Pileus | Spines | References |
|---|---|---|---|---|---|
| I | Croceum | Hydnellum aurantiacum | simple-septa | simple-septa | Maas Geesteranus 1975 [1] |
| H. auratile | simple-septa | simple-septa | Maas Geesteranus 1971 [26] | ||
| H. brunneorubrum | simple-septa | simple-septa | In this study | ||
| H. chrysinum | simple-septa | simple-septa | Baird 1986 [51] | ||
| H. earlianum | simple-septa | simple-septa | Baird et al. 2013 [27] | ||
| Inflatum | H. cristatum | simple-septa | simple-septa | Baird et al. 2013 [27] | |
| H. granulosum | simple-septa | simple-septa | In this study | ||
| H. inflatum | simple-septa | simple-septa | In this study | ||
| H. piperatum | simple-septa | simple-septa | Baird et al. 2013 [27] | ||
| H. mirabile | simple-septa | simple-septa | Maas Geesteranus 1975 [1] | ||
| Rhizomorphum | H. gracilipes | simple-septa | simple-septa | Koljalg & Renvall 2000 [35] | |
| Hydnellum sp 1 | simple-septa | simple-septa | In this study | ||
| Scabrosum | H. amygdaliolens | simple-septa | simple-septa | Rubio Casas et al. 2011 [52] | |
| H. coactum | simple-septa | simple-septa | Mu et al. 2020 [38] | ||
| H. fagiscabrosum | simple-septa | simple-septa | Nitare et al. 2021 [53] | ||
| H. fennicum | simple-septa | simple-septa | Hahn et al. 2018 [54] | ||
| H. grosselepidotum | simple-septa | simple-septa | Mu et al. 2020 [38] | ||
| H. illudens | simple-septa | simple-septa | Nitare et al. 2021 [53] | ||
| H. lepidum | simple-septa | simple-septa | Maas Geesteranus 1975 [1] | ||
| H. lidongensis | simple-septa | simple-septa | Mu et al. 2020 [38] | ||
| H. nemorosum | simple-septa | simple-septa | Nitare et al. 2021 [53] | ||
| H. scabrosellum | simple-septa | simple-septa | Nitare et al. 2021 [53] | ||
| H. scabrosum | simple-septa | simple-septa | Baird et al. 2013 [27] | ||
| H. underwoodii | simple-septa | simple-septa | Baird et al. 2013 [27] | ||
| Spongiosum | H. ferrugineum | simple-septa | simple-septa | Baird et al. 2013 [27] | |
| H. pineticola | simple-septa | simple-septa | Baird et al. 2013 [27] | ||
| H. spongiosipes | simple-septa | simple-septa | Baird et al. 2013 [27] | ||
| Hydnellum sp 2 | simple-septa | simple-septa | In this study | ||
| Violaceum | H. fuligineoviolaceum | simple-septa | simple-septa | Maas Geesteranus 1971 [26] | |
| H. fuscoindicum | simple-septa | simple-septa | Maas Geesteranus 1967 [65] | ||
| H. glaucopus | simple-septa | simple-septa | Maas Geesteranus 1975 [1] | ||
| H. joeides | simple-septa | simple-septa | Baird et al. 2013 [27] | ||
| H. roseoviolaceum | simple-septa | simple-septa | Nitare et al. 2021 [53] | ||
| Zonatum | H. atrorubrum | simple-septa | simple-septa | In this study | |
| H. bomiense | simple-septa | simple-septa | In this study | ||
| H. concrescens | simple-septa | simple-septa | Baird et al. 2013 [27] | ||
| H. dianthifolium | simple-septa | simple-septa | Loizides et al. 2016 [36] | ||
| H. parvum | simple-septa | simple-septa | Baird et al. 2013 [27] | ||
| H. rubidofuscum | simple-septa | simple-septa | In this study | ||
| H. scrobiculatum | simple-septa | simple-septa | Baird et al. 2013 [27] | ||
| H. squamulosum | simple-septa | simple-septa | In this study | ||
| H. subsuccosum | simple-septa | simple-septa | Baird et al. 2013 [27] | ||
| H. sulcatum | simple-septa | simple-septa | In this study | ||
| H. yunnanense | simple-septa | simple-septa | In this study | ||
| Hydnellum sp 3 | simple-septa | simple-septa | In this study | ||
| Hydnellum sp 4 | simple-septa | simple-septa | In this study | ||
| Hydnellum sp 5 | simple-septa | simple-septa | In this study | ||
| Others | H. complicatum | simple-septa | simple-septa | Baird et al. 2013 [27] | |
| H. cumulatum | simple-septa | simple-septa | Baird et al. 2013 [27] | ||
| H. lundellii | simple-septa | simple-septa | Maas Geesteranus 1975 [1] | ||
| H. martioflavum | simple-septa | simple-septa | Baird et al. 2013 [27] | ||
| II | Subindufibulatum | H. caeruleum | simple-septa, occasionally with clamp-connections | simple-septa | Baird & Khan 1986 [2]; In this study |
| H. ferrugipes | simple-septa, occasionally with clamp-connections | simple-septa | Baird et al. 2013 [27] | ||
| H. fibulatum | simple-septa, occasionally with clamp-connections | simple-septa | In this study | ||
| III | Others | H. peckii | mostly with clamp-connections, minority of simple-septa | mostly with clamp-connections, minority of simple-septa | Baird et al. 2013 [27]; In this study |
| H. versipelle | mostly with clamp-connections, minority of simple-septa | mostly with clamp-connections, minority of simple-septa | Baird et al. 2013 [27] | ||
| IV | Others | H. diabolus | clamp-connections | simple-septa | Baird et al. 2013 [27] |
| V | Hydnellum | H. atrospinosum | clamp-connections | clamp-connections | In this study |
| H. suaveolens | clamp-connections | clamp-connections | Baird et al. 2013 [27] | ||
| Caesispinosum | H. cyanopodium | clamp-connections | clamp-connections | Baird 1986 [51] | |
| H. scleropodium | clamp-connections | clamp-connections | Harrison 1964 [33] | ||
| Others | H. geogenium | clamp-connections | clamp-connections | Baird et al. 2013 [27] | |
| Sarcodon | Sarcodon aspratus | clamp-connections | clamp-connections | Maas Geesteranus 1971 [26] | |
| S. imbricatus | clamp-connections | clamp-connections | Baird et al. 2013 [27] | ||
| S. leucopus | clamp-connections | clamp-connections | Mleczko et al. 2011 [66] | ||
| S. quercinofibulatus | clamp-connections | clamp-connections | Pérez-De-Gregorio et al. 2011 [67] | ||
| S. scabripes | clamp-connections | clamp-connections | Baird et al. 2013 [27] | ||
| S. squamosus | clamp-connections | clamp-connections | Baird 1986 [34] | ||
| Others | H. regium | hyphae with few simple-septa and with a few clamp-connections | Harrison 1964 [33] | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mu, Y.-H.; Yu, J.-R.; Cao, T.; Wang, X.-H.; Yuan, H.-S. Multi-Gene Phylogeny and Taxonomy of Hydnellum (Bankeraceae, Basidiomycota) from China. J. Fungi 2021, 7, 818. https://doi.org/10.3390/jof7100818
Mu Y-H, Yu J-R, Cao T, Wang X-H, Yuan H-S. Multi-Gene Phylogeny and Taxonomy of Hydnellum (Bankeraceae, Basidiomycota) from China. Journal of Fungi. 2021; 7(10):818. https://doi.org/10.3390/jof7100818
Chicago/Turabian StyleMu, Yan-Hong, Jia-Rui Yu, Ting Cao, Xiang-Hua Wang, and Hai-Sheng Yuan. 2021. "Multi-Gene Phylogeny and Taxonomy of Hydnellum (Bankeraceae, Basidiomycota) from China" Journal of Fungi 7, no. 10: 818. https://doi.org/10.3390/jof7100818
APA StyleMu, Y.-H., Yu, J.-R., Cao, T., Wang, X.-H., & Yuan, H.-S. (2021). Multi-Gene Phylogeny and Taxonomy of Hydnellum (Bankeraceae, Basidiomycota) from China. Journal of Fungi, 7(10), 818. https://doi.org/10.3390/jof7100818