Understanding Ustilago maydis Infection of Multiple Maize Organs


Abstract
1. Introduction
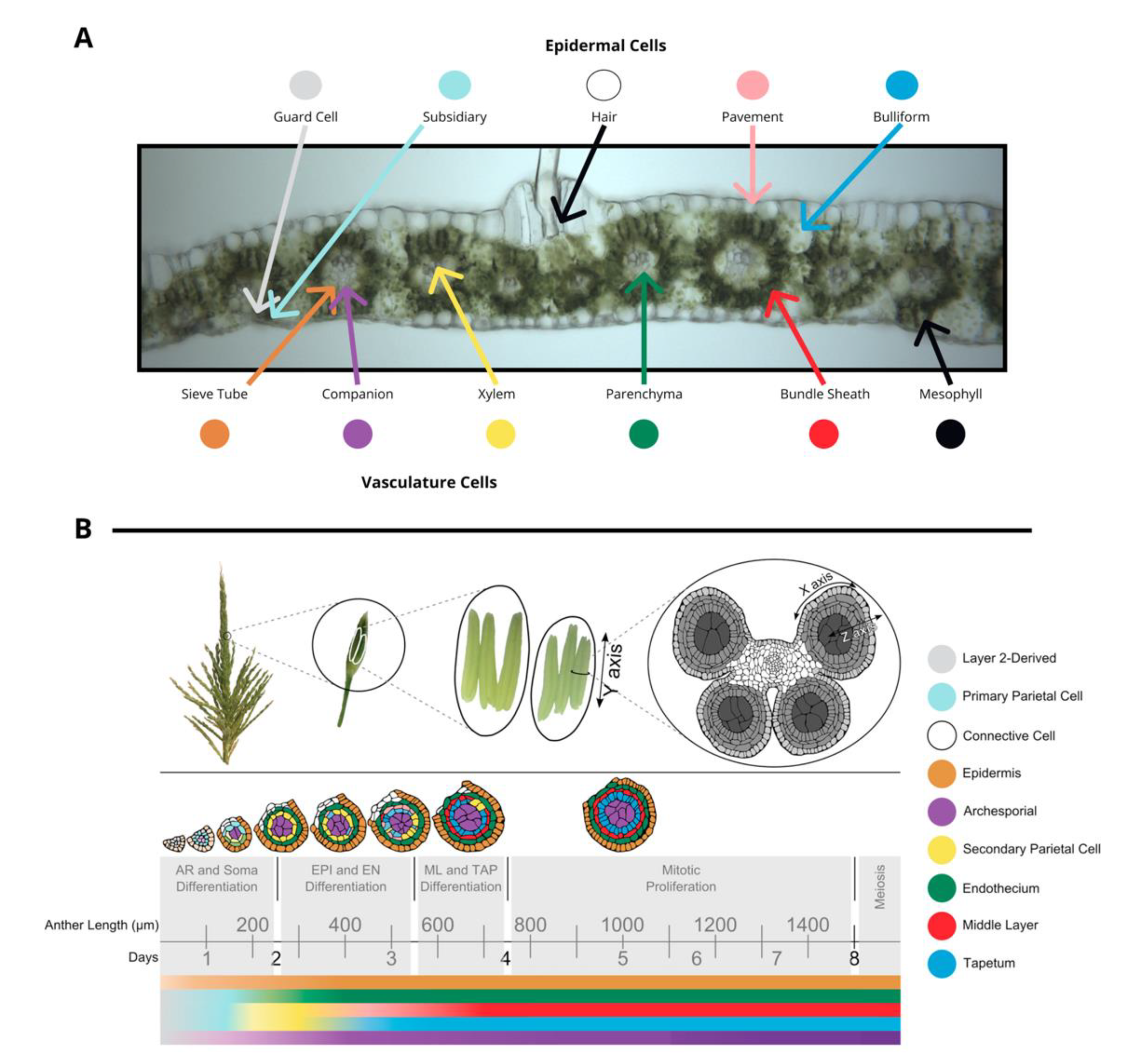
1.1. Leaf Characteristics
1.2. Reproductive Organ Characteristics
1.3. Infection and Tumor Progression
2. Exploiting U. maydis Mutants to Define Requirements for Tumor Formation
Future Directions
- Mutant characterization through transcriptomics: One way to achieve greater sensitivity for determining if a mutant impacts U. maydis virulence is to look for meaningful differences in gene expression. A more in-depth time course characterization of the normal progression of SG200 infection would establish a better baseline for what to expect. Another way to increase sensitivity is to use single cell RNA-seq (scRNA-seq) instead of bulk organ (such as an entire leaf or anther) or tissue sequencing. This would provide information from both infected and uninfected cells in the same organ as well as identifying responses in individual cell types and decreasing the amount of noise in complex organ data. Using transcriptomics as a screening approach also has the advantage of generating better information about which pathways are being impacted by a given mutation as well as at what point infection development is interrupted.
- Involvement of fungal effectors in disease progression: Thus far, characterization of the mechanism of action of most effector genes is understood at a superficial level, i.e., the specific maize process targeted. Furthermore, it is difficult to connect effector disruption to what is changing in maize cells more generally. For example, what roles do secreted fungal effectors play in host cell entry into the tumor pathway? Moreover, which specific fungal effectors are required in sequential steps of host cell redirection into the tumor pathway? Are there multiple tumor pathways, depending on cell type?
3. Exploiting Maize Mutants to Define Requirements for Tumor Formation
Future Directions
- 1.
- Exploiting maize mutants: The success thus far in exploiting maize mutants to dissect tumorigenesis should inspire additional experiments with host plants impaired in specific processes. Table 1 lists a few examples that could be fruitful. More information about each mutant is available at MaizeGDB by typing in a gene name or a process in the search box and requesting all data to retrieve phenotypic descriptions, photographs, genetic analysis, gene models, and references. Seeds are available for worldwide distribution from the Maize Coop (with links at MaizeGDB). Many of the examples involve generating a chimeric maize organ with normal and mutant tissue side-by-side. This arrangement permits scoring “normal” maize–fungal interaction near mutant host tissue–fungal interaction outcome.
- 2.
- Applying new analytical methods: New technologies also provide new opportunities to explore host–pathogen activities. Thus far, cell type information has been obtained on cell groups recovered by laser capture microdissection, but now that scRNA-seq is established for maize [27], an obvious approach is to evaluate the diverse cell types of seedling leaves, adult leaves, ears, and tassels in both infected and uninfected organs. Comparison of infected to uninfected cells between plants should identify both fungal and maize gene expression changes associated with infection progression resulting in tumor formation in every impacted cell type. Comparison of infected and uninfected cells within an infected organ from the same plant should highlight cell autonomous changes in host cells, i.e., those reflecting infection by U. maydis, and should allow discovery of non-cell-autonomous host responses as infected host cells alter the development or physiology of neighbors or cells distal from the infection. Effectors secreted extracellularly into the biotrophic interaction zone have a limited ability to diffuse to other cells; however, effectors secreted intracellularly have been shown to diffuse into and prime a response in secondary cells. In addition to the changes caused by effectors directly, maize cells can signal to each other through plasmodesmata or by utilizing secreted molecules (ions, hormones, peptides) or altered plasma membrane properties (lipid composition or modifications of existing lipids) that influence tissue or organ physiology or development. Alterations in host cell physiology during infection could thus be expected to cause non-cell-autonomous events, and it will be very interesting to determine the spatiotemporal distribution of this class of responses.
- 3.
- Proteomics: Quantitative proteomics could contribute substantially to our understanding of host–pathogen interaction. mRNA analysis through RNA-seq is only a proxy for the “real actors” in host cell responses, the protein complement. In addition to improvements in mass spectrometry—both in the range of protein types that can be analyzed and miniaturization of sample size—that will be discovery tools, proximity labeling such as Turbo-ID can establish specific fungal effector–maize protein interactions, identify membrane proteins involved in binding transported effectors, etc. [30].
- 4.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brefort, T.; Doehlemann, G.; Mendoza-Mendoza, A.; Reissmann, S.; Djamei, A.; Kahmann, R. Ustilago maydis as a Pathogen. Annu. Rev. Phytopathol. 2009, 47, 423–445. [Google Scholar] [CrossRef] [PubMed]
- Kämper, J.; Kahmann, R.; Bölker, M.; Ma, L.-J.; Brefort, T.; Saville, B.J.; Banuett, F.; Kronstad, J.W.; Gold, S.E.; Müller, O.; et al. Insights from the Genome of the Biotrophic Fungal Plant Pathogen Ustilago maydis. Nature 2006, 444, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Kelliher, T.; Nguyen, L.; Walbot, V. Ustilago maydis Reprograms Cell Proliferation in Maize Anthers. Plant J. 2013, 75, 903–914. [Google Scholar] [CrossRef]
- Matei, A.; Ernst, C.; Günl, M.; Thiele, B.; Altmüller, J.; Walbot, V.; Usadel, B.; Doehlemann, G. How to Make a Tumour: Cell Type Specific Dissection of Ustilago maydis- Induced Tumour Development in Maize Leaves. New Phytol. 2018, 217, 1681–1695. [Google Scholar] [CrossRef]
- Egger, R.L.; Walbot, V. A Framework for Evaluating Developmental Defects at the Cellular Level: An Example from Ten Maize Anther Mutants Using Morphological and Molecular Data. Dev. Biol. 2016, 419, 26–40. [Google Scholar] [CrossRef]
- Kelliher, T.; Walbot, V. Emergence and Patterning of the Five Cell Types of the Zea Mays Anther Locule. Dev. Biol. 2011, 350, 32–49. [Google Scholar] [CrossRef]
- Doehlemann, G.; Wahl, R.; Vranes, M.; de Vries, R.P.; Kämper, J.; Kahmann, R. Establishment of Compatibility in the Ustilago maydis/Maize Pathosystem. J. Plant Physiol. 2008, 165, 29–40. [Google Scholar] [CrossRef]
- Doehlemann, G.; Wahl, R.; Horst, R.J.; Voll, L.M.; Usadel, B.; Poree, F.; Stitt, M.; Pons-Kühnemann, J.; Sonnewald, U.; Kahmann, R.; et al. Reprogramming a Maize Plant: Transcriptional and Metabolic Changes Induced by the Fungal Biotroph Ustilago maydis: Plant Response to U. maydis Infection. Plant J. 2008, 56, 181–195. [Google Scholar] [CrossRef]
- Walbot, V.; Skibbe, D.S. Maize Host Requirements for Ustilago maydis Tumor Induction. Sex. Plant Reprod. 2010, 23, 1–13. [Google Scholar] [CrossRef][Green Version]
- Schirawski, J.; Mannhaupt, G.; Münch, K.; Brefort, T.; Schipper, K.; Doehlemann, G.; Di Stasio, M.; Rössel, N.; Mendoza-Mendoza, A.; Pester, D.; et al. Pathogenicity Determinants in Smut Fungi Revealed by Genome Comparison. Science 2010, 330, 1546–1548. [Google Scholar] [CrossRef]
- Ökmen, B.; Kemmerich, B.; Hilbig, D.; Wemhöner, R.; Aschenbroich, J.; Perrar, A.; Huesgen, P.F.; Schipper, K.; Doehlemann, G. Dual Function of a Secreted Fungalysin Metalloprotease in Ustilago maydis. New Phytol. 2018, 220, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Doehlemann, G.; Reissmann, S.; Aßmann, D.; Fleckenstein, M.; Kahmann, R. Two Linked Genes Encoding a Secreted Effector and a Membrane Protein Are Essential for Ustilago maydis-Induced Tumour Formation: A Novel Gene Cluster of Ustilago maydis. Mol. Microbiol. 2011, 81, 751–766. [Google Scholar] [CrossRef] [PubMed]
- Schilling, L.; Matei, A.; Redkar, A.; Walbot, V.; Doehlemann, G. Virulence of the Maize Smut Ustilago maydis Is Shaped by Organ-Specific Effectors: Organ-Specific Effectors of Ustilago maydis. Mol. Plant Pathol. 2014, 15, 780–789. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Gollin, I.; Rössel, N.; Kahmann, R. The Functionally Conserved Effector Sta1 Is a Fungal Cell Wall Protein Required for Virulence in Ustilago maydis. New Phytol. 2020, 227, 185–199. [Google Scholar] [CrossRef]
- Brefort, T.; Tanaka, S.; Neidig, N.; Doehlemann, G.; Vincon, V.; Kahmann, R. Characterization of the Largest Effector Gene Cluster of Ustilago maydis. PLoS Pathog. 2014, 10, e1003866. [Google Scholar] [CrossRef]
- Tanaka, S.; Brefort, T.; Neidig, N.; Djamei, A.; Kahnt, J.; Vermerris, W.; Koenig, S.; Feussner, K.; Feussner, I.; Kahmann, R. A Secreted Ustilago maydis Effector Promotes Virulence by Targeting Anthocyanin Biosynthesis in Maize. eLife 2014, 3, e01355. [Google Scholar] [CrossRef]
- Tanaka, S.; Schweizer, G.; Rössel, N.; Fukada, F.; Thines, M.; Kahmann, R. Neofunctionalization of the Secreted Tin2 Effector in the Fungal Pathogen Ustilago maydis. Nat. Microbiol. 2019, 4, 251–257. [Google Scholar] [CrossRef]
- Redkar, A.; Hoser, R.; Schilling, L.; Zechmann, B.; Krzymowska, M.; Walbot, V.; Doehlemann, G. A Secreted Effector Protein of Ustilago maydis Guides Maize Leaf Cells to Form Tumors. Plant Cell 2015, 27, 1332–1351. [Google Scholar] [CrossRef]
- Villajuana-Bonequi, M.; Matei, A.; Ernst, C.; Hallab, A.; Usadel, B.; Doehlemann, G. Cell Type Specific Transcriptional Reprogramming of Maize Leaves during Ustilago maydis Induced Tumor Formation. Sci. Rep. 2019, 9, 10227. [Google Scholar] [CrossRef]
- Baumgarten, A.M.; Suresh, J.; May, G.; Phillips, R.L. Mapping QTLs Contributing to Ustilago maydis Resistance in Specific Plant Tissues of Maize. Theor. Appl. Genet. 2007, 114, 1229–1238. [Google Scholar] [CrossRef]
- Jackson-Ziems, T.A. Smut Diseases of Corn. Neb. Ext. Publ. 2014, G2223. [Google Scholar]
- Callow, J.A. Endopolyploidy in Maize Smut Neoplasms Induced by the Maize Smut Fungus, Ustilago maydis. New Phytol. 1975, 75, 253–257. [Google Scholar] [CrossRef]
- Callow, J.A.; Ling, I.T. Histology of Neoplasms and Chlorotic Lesions in Maize Seedlings Following the Injection of Sporidia of Ustilago maydis (DC) Corda. Physiol. Plant Pathol. 1973, 3, 489–494. [Google Scholar] [CrossRef]
- Skibbe, D.S.; Doehlemann, G.; Fernandes, J.; Walbot, V. Maize Tumors Caused by Ustilago maydis Require Organ-Specific Genes in Host and Pathogen. Science 2010, 328, 89–92. [Google Scholar] [CrossRef]
- Fernandes, J.; Brendel, V.; Gai, X.; Lal, S.; Chandler, V.L.; Elumalai, R.P.; Galbraith, D.W.; Pierson, E.A.; Walbot, V. Comparison of RNA Expression Profiles Based on Maize Expressed Sequence Tag Frequency Analysis and Micro-Array Hybridization. Plant Physiol. 2002, 128, 896–910. [Google Scholar] [CrossRef]
- Sosso, D.; van der Linde, K.; Bezrutczyk, M.; Schuler, D.; Schneider, K.; Kämper, J.; Walbot, V. Sugar Partitioning between Ustilago maydis and Its Host Zea Mays L during Infection. Plant Physiol. 2019, 179, 1373–1385. [Google Scholar] [CrossRef]
- Nelms, B.; Walbot, V. Defining the Developmental Program Leading to Meiosis in Maize. Science 2019, 364, 52–56. [Google Scholar] [CrossRef]
- van der Linde, K.; Timofejeva, L.; Egger, R.L.; Ilau, B.; Hammond, R.; Teng, C.; Meyers, B.C.; Doehlemann, G.; Walbot, V. Pathogen Trojan Horse Delivers Bioactive Host Protein to Alter Maize Anther Cell Behavior in Situ. Plant Cell 2018, 30, 528–542. [Google Scholar] [CrossRef]
- Fiedler, I.-C.; Weiberg, A.; van der Linde, K. Using Ustilago maydis as a Trojan Horse for in Situ Delivery of Maize Proteins. J. Vis. Exp. 2019, 58746. [Google Scholar] [CrossRef]
- Cho, K.F.; Branon, T.C.; Rajeev, S.; Svinkina, T.; Udeshi, N.D.; Thoudam, T.; Kwak, C.; Rhee, H.-W.; Lee, I.-K.; Carr, S.A.; et al. Split-TurboID Enables Contact-Dependent Proximity Labeling in Cells. Proc. Natl. Acad. Sci. USA 2020, 117, 12143–12154. [Google Scholar] [CrossRef]
- Simon, M.L.A.; Platre, M.P.; Assil, S.; van Wijk, R.; Chen, W.Y.; Chory, J.; Dreux, M.; Munnik, T.; Jaillais, Y. A Multi-Colour/Multi-Affinity Marker Set to Visualize Phosphoinositide Dynamics in Arabidopsis. Plant J. 2014, 77, 322–337. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Bouza, M.; Wu, C.; Guo, H.; Huang, D.; Doron, G.; Temenoff, J.S.; Stecenko, A.A.; Wang, Z.L.; Fernández, F.M. Sub-Nanoliter Metabolomics via Mass Spectrometry to Characterize Volume-Limited Samples. Nat. Commun. 2020, 11, 5625. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Disrupted Process | Gene Name | Notes |
|---|---|---|
| Photosynthesis | iojap | Random loss of chloroplast ribosomes results in stripes of white, yellow, pale green, and normal green on leaves. In this chimera, you can test mutant and impaired tissue simultaneously. |
| Chloroplast greening | zb4 | Transverse leaf sectors (physiological chimera likely caused by day or night temperature) allow study of fungal growth in affected and normal areas simultaneously. |
| Leaf sugar metabolism | tie dyed 1, 2 | Differentially impacted areas on the same leaf. |
| Anther developmental timing | ms8 | Heterochronic anther mutant in which specific steps are delayed; other examples include csmd1. |
| Conditional anther growth | dcl5, ocl4 | Temperature-dependent phenotypes permit generating a chimeric tassel of normal and arrested anthers by temperature treatment for 1 or a few days. |
| Response to pathogens | Diverse les | Mutants spontaneously express symptoms of specific pathogen infections resulting in zones with or without host leaf responses. |
| Leaf and stem development | dwarf, nana | Multiple loci disrupted in gibberellin (dwarf) or brassinosteriod (nana1, 2) biogenesis or response. |
| Any target | Utilize CRISPR/Cas9 to disrupt genes of interest. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferris, A.C.; Walbot, V. Understanding Ustilago maydis Infection of Multiple Maize Organs. J. Fungi 2021, 7, 8. https://doi.org/10.3390/jof7010008
Ferris AC, Walbot V. Understanding Ustilago maydis Infection of Multiple Maize Organs. Journal of Fungi. 2021; 7(1):8. https://doi.org/10.3390/jof7010008
Chicago/Turabian StyleFerris, Alex C, and Virginia Walbot. 2021. "Understanding Ustilago maydis Infection of Multiple Maize Organs" Journal of Fungi 7, no. 1: 8. https://doi.org/10.3390/jof7010008
APA StyleFerris, A. C., & Walbot, V. (2021). Understanding Ustilago maydis Infection of Multiple Maize Organs. Journal of Fungi, 7(1), 8. https://doi.org/10.3390/jof7010008