Degenerated Virulence and Irregular Development of Fusarium oxysporum f. sp. niveum Induced by Successive Subculture

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Pathogen Resources and Cultivation
2.2. Plant Materials and Growth
2.3. Generation of F. oxysporum Degenerated Variants
2.4. Phenotyping of Fungal Growth
2.5. Methods for Pathogen Inoculation
2.6. Cell Wall-Degrading Enzymes Assay
2.7. Real-Time Quantitative Reverse Transcription PCR (qRT-PCR) of Virulence Gene Expression
3. Results
3.1. Successive Subculture of Aged Mycelia Induces the Cultural Transformation
3.2. Variants of Fon Demonstrated Significant Morphology Changes
3.3. Variants of Fon Reduced Their Virulence to Susceptible Watermelon
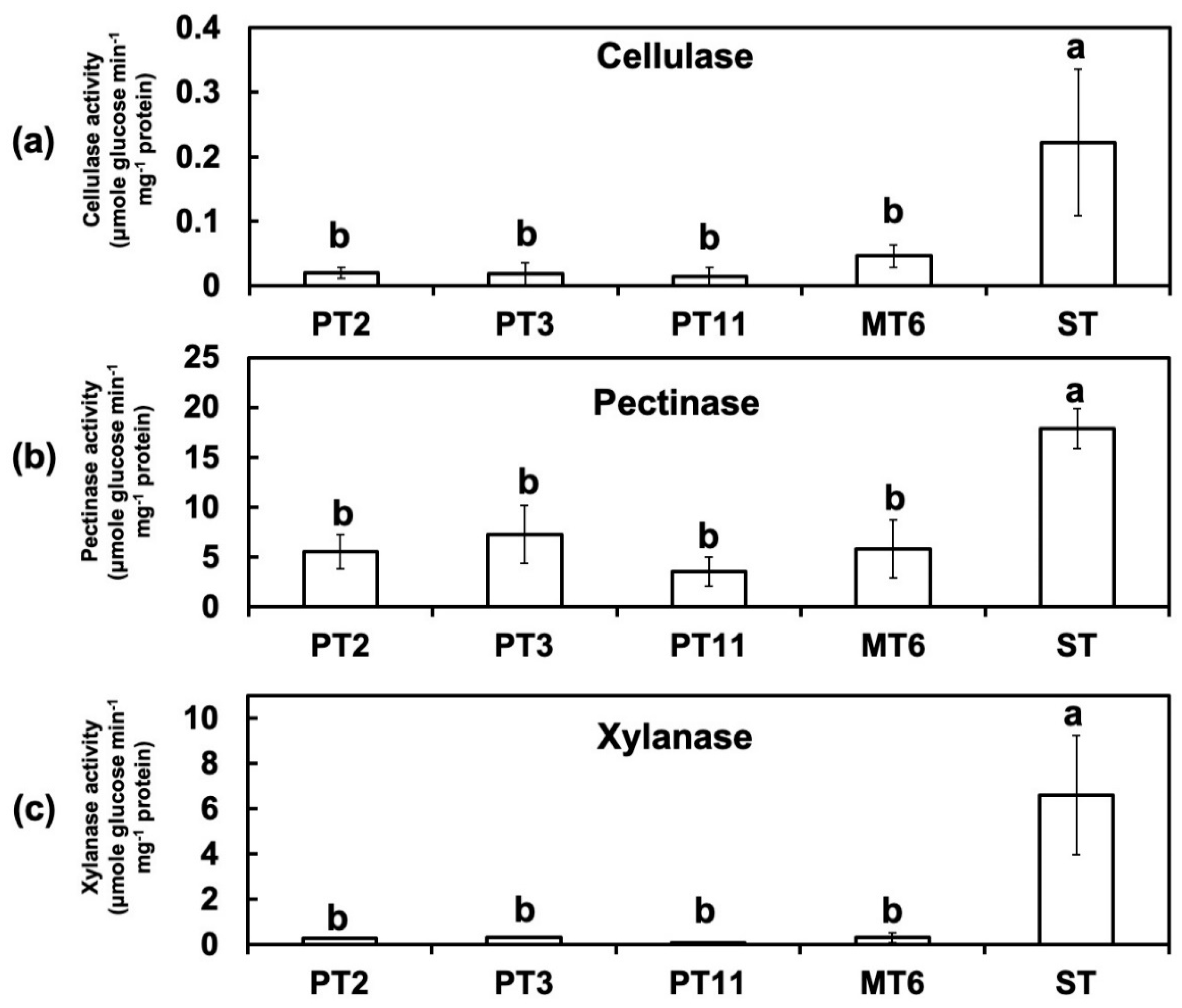
3.4. Activites of Cell Wall-Degrading Enzymes (CWDEs) Were Reduced in Degenerated Variants
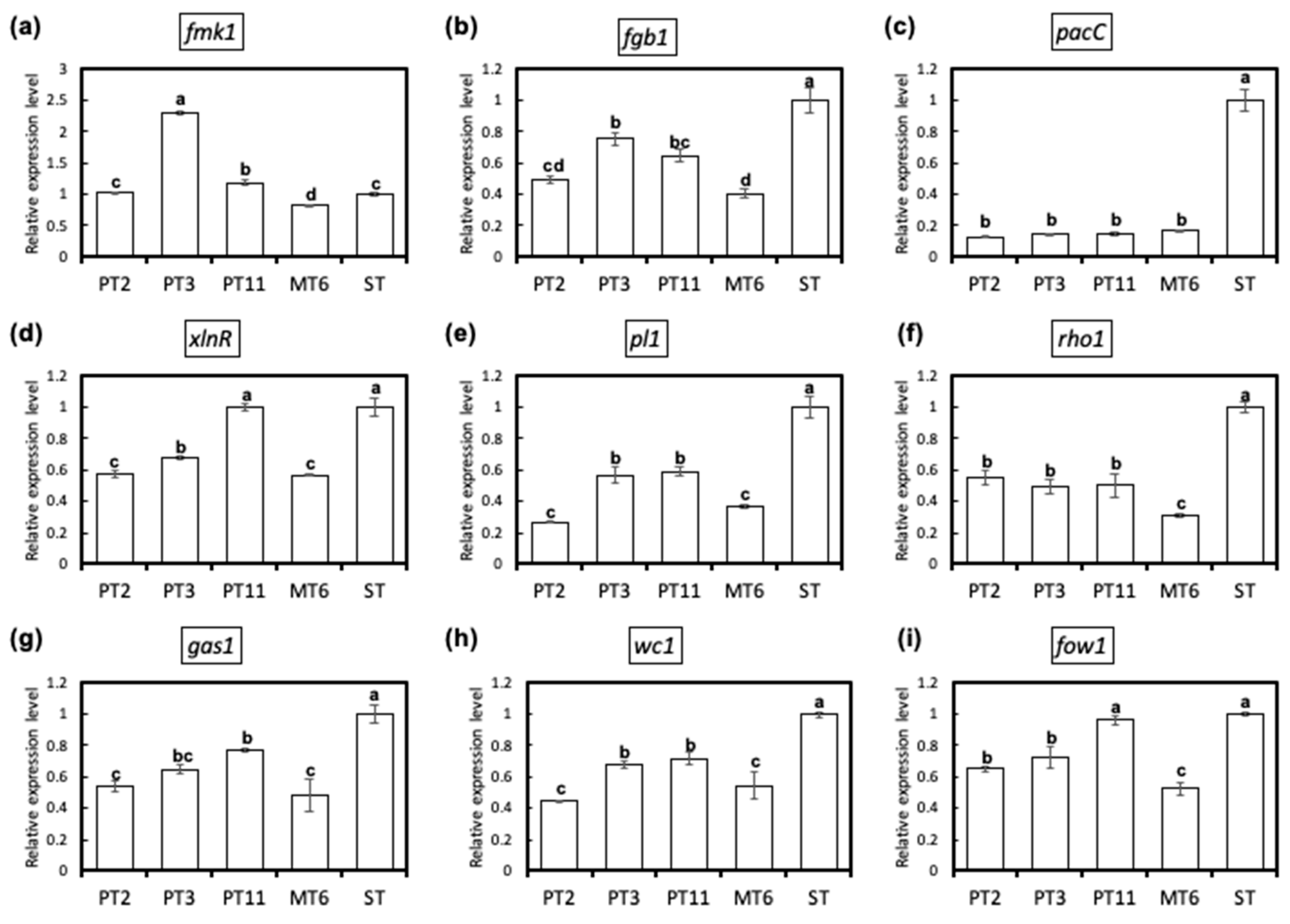
3.5. Fungus Degeneration Affects the Expression of Virulence-Related Genes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.-H.; Lin, Y.-H.; Chen, K.-S.; Huang, J.-W.; Hsiao, S.-C.; Chang, P.-F.L. Cell wall reinforcement in watermelon shoot base related to its resistance to Fusarium wilt caused by Fusarium oxysporum f. sp. niveum. J. Agric. Sci. 2015, 153, 296–305. [Google Scholar] [CrossRef]
- Michielse, C.B.; Rep, M. Pathogen profile update: Fusarium oxysporum. Mol. Plant Pathol. 2009, 10, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-H.; Chen, K.-S.; Chang, J.-Y.; Wan, Y.-L.; Hsu, C.-C.; Huang, J.-W.; Chang, P.-F.L. Development of the molecular methods for rapid detection and differentiation of Fusarium oxysporum and F. oxysporum f. sp. niveum in Taiwan. New Biotechnol. 2010, 27, 409–418. [Google Scholar] [CrossRef]
- Martyn, R.D.; Netzer, D. Resistance to races 0, 1, and 2 of Fusarium wilt of watermelon in Citrullus sp. PI-296341 -FR. HortScience 1991, 26, 429–432. [Google Scholar] [CrossRef]
- Xie, X.-G.; Huang, C.-Y.; Cai, Z.-D.; Chen, Y.; Dai, C.-C. Targeted acquisition of Fusarium oxysporum f. sp. niveum toxin-deficient mutant and its effects on watermelon Fusarium wilt. J. Agric. Food Chem. 2019, 67, 8536–8547. [Google Scholar] [CrossRef]
- Srinivas, C.; Nirmala Devi, D.; Narasimha Murthy, K.; Mohan, C.D.; Lakshmeesha, T.R.; Singh, B.; Kalagatur, N.K.; Niranjana, S.R.; Hashem, A.; Alqarawi, A.A.; et al. Fusarium oxysporum f. sp. lycopersici causal agent of vascular wilt disease of tomato: Biology to diversity—A review. Saudi J. Biol. Sci. 2019, 26, 1315–1324. [Google Scholar] [CrossRef]
- Turrà, D.; El Ghalid, M.; Rossi, F.; Di Pietro, A. Fungal pathogen uses sex pheromone receptor for chemotropic sensing of host plant signals. Nature 2015, 527, 521–524. [Google Scholar] [CrossRef]
- Gordon, T.R. Fusarium oxysporum and the Fusarium wilt syndrome. Annu. Rev. Phytopathol. 2017, 55, 23–39. [Google Scholar] [CrossRef]
- Warman, N.M.; Aitken, E.A.B. The movement of Fusarium oxysporum f. sp. cubense (sub-tropical race 4) in susceptible cultivars of banana. Front. Plant Sci. 2018, 9, 1748. [Google Scholar] [CrossRef]
- Kikot, G.E.; Hours, R.A.; Alconada, T.M. Contribution of cell wall degrading enzymes to pathogenesis of Fusarium graminearum: A review. J. Basic Microbiol. 2009, 49, 231–241. [Google Scholar] [CrossRef]
- Rauwane, M.E.; Ogugua, U.V.; Kalu, C.M.; Ledwaba, L.K.; Woldesemayat, A.A.; Ntushelo, K. Pathogenicity and virulence factors of Fusarium graminearum including factors discovered using next generation sequencing technologies and proteomics. Microorganisms 2020, 8, 305. [Google Scholar] [CrossRef] [PubMed]
- Ansari, M.A.; Butt, T.M. Effects of successive subculturing on stability, virulence, conidial yield, germination and shelf-life of entomopathogenic fungi. J. Appl. Microbiol. 2011, 110, 1460–1469. [Google Scholar] [CrossRef] [PubMed]
- Wing, N.; Burgess, L.W.; Bryden, W.L. Cultural degeneration in two Fusarium species and its effects on toxigenicity and cultural morphology. Mycol. Res. 1995, 99, 615–620. [Google Scholar] [CrossRef]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual, 1st ed.; Blackwell Pub: Ames, Iowa, 2006; p. 387. ISBN 978-0-8138-1919-8. [Google Scholar]
- Nelson, P.E.; Dignani, M.C.; Anaissie, E.J. Taxonomy, biology, and clinical aspects of Fusarium species. Clin. Microbiol. Rev. 1994, 7, 479–504. [Google Scholar] [CrossRef] [PubMed]
- Nash, S.M.; Snyder, W.C. Quantitative estimations by plate counts of propagules of the Bean root rot Fusarium in field soils. Phytopathology 1962, 52, 567–572. [Google Scholar]
- Awuah, R.T.; Lorbeer, J.W. Nature of cultural variability in Fusarium oxysporum f. sp. apii race 2. Phytopathology 1988, 78, 385–389. [Google Scholar] [CrossRef]
- King, B.C.; Donnelly, M.K.; Bergstrom, G.C.; Walker, L.P.; Gibson, D.M. An optimized microplate assay system for quantitative evaluation of plant cell wall–degrading enzyme activity of fungal culture extracts. Biotechnol. Bioeng. 2009, 102, 1033–1044. [Google Scholar] [CrossRef]
- Booth, C. The genus Fusarium; Commonwealth Mycological Institute: Kew, Surrey, UK, 1971; p. 237. ISBN SEN851980465. [Google Scholar]
- Kruger, N.J. The Bradford Method for Protein Quantitation. In The Protein Protocols Handbook; Walker, J.M., Ed.; Springer Protocols Handbooks; Humana Press: Totowa, NJ, USA, 2009; pp. 17–24. ISBN 978-1-59745-198-7. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Khani, M.; Davidson, J.A.; Sosnowski, M.R.; Scott, E.S. Atypical growth of Phoma koolunga on cultural media and on plants artificially inoculated in environmentally controlled conditions. Australas. Plant Pathol. 2017, 46, 511–514. [Google Scholar] [CrossRef]
- Sun, S.-K.; Huang, J.-W. Variation in the watermelon wilt pathogen, Fusarium oxysporum f. sp. niveum. Plant Pathol. Bull. 1979, 21, 313–332. [Google Scholar]
- Chu, Y.-M.; Jeon, J.-J.; Yea, S.-J.; Kim, Y.-H.; Yun, S.-H.; Lee, Y.-W.; Kim, K.-H. Double-stranded RNA mycovirus from Fusarium graminearum. Appl. Environ. Microbiol. 2002, 68, 2529–2534. [Google Scholar] [CrossRef]
- Hart, L.P. The effect of time of exposure to inoculum, plant age, root development, and root wounding on Fusarium yellows of celery. Phytopathology 1981, 71, 77. [Google Scholar] [CrossRef]
- Shaw, R.K.; Shaik, M.; Mir, Z.A.; Prasad, M.S.L.; Prasad, R.D.; Senthilvel, S. Establishing a high throughput screening method for large scale phenotyping of castor genotypes for resistance to Fusarium wilt disease. Phytoparasitica 2016, 44, 539–548. [Google Scholar] [CrossRef]
- Saxena, I.M. The Plant Cell Wall: Barrier and Facilitator of Environmental Perception. In Sensory Biology of Plants; Sopory, S., Ed.; Springer: Singapore, 2019; pp. 453–476. ISBN 9789811389221. [Google Scholar]
- Lampugnani, E.R.; Khan, G.A.; Somssich, M.; Persson, S. Building a plant cell wall at a glance. J. Cell Sci. 2018, 131, jcs207373. [Google Scholar] [CrossRef] [PubMed]
- Jorge, I.; Navas-Cortés, J.A.; Jiménez-Díaz, R.M.; Tena, M. Cell wall degrading enzymes in Fusarium wilt of chickpea: Correlation between pectinase and xylanase activities and disease development in plants infected with two pathogenic races of Fusarium oxysporum f. sp. ciceris. Can. J. Bot. 2006, 84, 1395–1404. [Google Scholar] [CrossRef]
- Caracuel, Z.; Martínez-Rocha, A.L.; Di Pietro, A.; Madrid, M.P.; Roncero, M.I.G. Fusarium oxysporum gas1 encodes a putative β-1, 3-glucanosyltransferase required for virulence on tomato plants. MPMI 2005, 18, 1140–1147. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Jarana, J.; Martínez-Rocha, A.L.; Roldán-Rodriguez, R.; Roncero, M.I.; Pietro, A.D. Fusarium oxysporum G-protein β subunit Fgb1 regulates hyphal growth, development, and virulence through multiple signalling pathways. Fungal Genet. Biol. 2005, 42, 61–72. [Google Scholar] [CrossRef]
- Inoue, I.; Namiki, F.; Tsuge, T. Plant colonization by the vascular wilt fungus Fusarium oxysporum requires FOW1, a gene encoding a mitochondrial protein. Plant Cell 2002, 14, 1869–1883. [Google Scholar] [CrossRef]
- Caracuel, Z.; Roncero, M.I.G.; Espeso, E.A.; González-Verdejo, C.I.; García-Maceira, F.I.; Di Pietro, A. The pH signalling transcription factor PacC controls virulence in the plant pathogen Fusarium oxysporum. Mol. Microbiol. 2003, 48, 765–779. [Google Scholar] [CrossRef]
- Daboussi, M.-J.; Capy, P. Transposable elements in filamentous fungi. Annu. Rev. Microbiol. 2003, 57, 275–299. [Google Scholar] [CrossRef] [PubMed]
- Torres-Trenas, A.; Prieto, P.; Cañizares, M.C.; García-Pedrajas, M.D.; Pérez-Artés, E. Mycovirus Fusarium oxysporum f. sp. dianthi virus 1 decreases the colonizing efficiency of its fungal host. Front. Cell. Infect. Microbiol. 2019, 9, 51. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Zhang, Z.; Li, B.; Tian, S. The pattern and function of DNA methylation in fungal plant pathogens. Microorganisms 2020, 8, 227. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amplicon | Most Homologue | Amplification Primer | |||
|---|---|---|---|---|---|
| Gene Name/bp | Protein Function | Description/Accession Number | Identity (%) | Primer Name | Sequence (5′–3′) |
| ef1α/115 | Housekeeping gene used in this study | Gibberella zeae PH-1 elongation factor 1-alpha partial mRNA/XM388987 | 98 | ef1aF | TAAGGGTTCCTTCAAGTACG |
| ef1aR | GGTGACATAGTAGCGAGGAG | ||||
| fgb1/128 | Signal transduction | F. oxysporum f. sp. lycopersici G protein beta subunit gene (fgb1), complete cds/AY219172 | 98 | fgb1F | TGGTGACATGACCTGTATGA |
| fgb1R | CACCAGAGATGAAGGTGTTT | ||||
| fow1/142 | Signal transduction | F. oxysporum plasmid pWB60Sl FOW1 gene (fow1) for putative mitochondrial carrier protein, complete cds/AB078975 | 100 | fow1F | GAGATCACCAAGCACAAGAT |
| fow1R | AGTTGGTCATCTGTCGGATA | ||||
| fmk1/147 | Mitogen-activate protein kinase | F. oxysporum f. sp. lycopersici mitogen-activate protein kinase gene (fmk1), complete cds/AF286533.1 | 99 | fmk1F | ACATTCGATCGCTCCCCTTC |
| fmk1R | ATGGGTGCTTCAGAGCTTCC | ||||
| gas1/119 | Fungal cell wall biosynthesis | F. oxysporum beta-1,3-glucanosyltransferase gene (gas1), complete cds/AY884608 | 98 | gas1F | GTGAGACCAACGACAAGACT |
| gas1R | GATAGCGTAAGTTCGGATGA | ||||
| pacC/124 | Negative regulator of pH signaling | F. oxysporum pH transcription factor gene (pacC), complete cds/AY125958 | 95 | pacCF | CTGGTCCTACTCCTAGCAAA |
| pacCR | TAGATGGTGTCTTGCATCTG | ||||
| pl1/113 | Pectin degrading enzyme | F. oxysporum f. sp. lycopersici pectate lyase gene (pl1), complete cds/AF080485 | 98 | pl1F | TTCATTCCCTACTGCTGTTC |
| pl1R | TAGCATCCTTCTCACCAGTT | ||||
| xlnR/133 | Transcription activator of xylanolytic system | F. oxysporum f. sp. lycopersici XlnR gene (xlnR), complete cds/EF057395.1 | 99 | xlnRF | TGGACAAAGGACGGAACAGG |
| xlnRR | CACCCGAAAGAGAGTGTCCC | ||||
| rho1/104 | Fungal cell wall biosynthesis | F. oxysporum small GTPase-binding protein gene (rho1), complete cds/AY884607 | 99 | rho1F | GATACGACCAGAAGACCATC |
| rho1R | GTACTTGTAGGCAGCGATCT | ||||
| wc1/129 | Photoreceptor | F. oxysporum f. sp. lycopersici white collar 1 gene (wc1), complete cds/EU327188 | 99 | wc1F | TCAACAGTGTGTCATCCATC |
| wc1R | GCTTTCGAACCAAGTGTAAC | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, T.-H.; Lin, Y.-H.; Wan, Y.-L.; Chen, K.-S.; Huang, J.-W.; Chang, P.-F.L. Degenerated Virulence and Irregular Development of Fusarium oxysporum f. sp. niveum Induced by Successive Subculture. J. Fungi 2020, 6, 382. https://doi.org/10.3390/jof6040382
Chang T-H, Lin Y-H, Wan Y-L, Chen K-S, Huang J-W, Chang P-FL. Degenerated Virulence and Irregular Development of Fusarium oxysporum f. sp. niveum Induced by Successive Subculture. Journal of Fungi. 2020; 6(4):382. https://doi.org/10.3390/jof6040382
Chicago/Turabian StyleChang, Tao-Ho, Ying-Hong Lin, Yu-Ling Wan, Kan-Shu Chen, Jenn-Wen Huang, and Pi-Fang Linda Chang. 2020. "Degenerated Virulence and Irregular Development of Fusarium oxysporum f. sp. niveum Induced by Successive Subculture" Journal of Fungi 6, no. 4: 382. https://doi.org/10.3390/jof6040382
APA StyleChang, T.-H., Lin, Y.-H., Wan, Y.-L., Chen, K.-S., Huang, J.-W., & Chang, P.-F. L. (2020). Degenerated Virulence and Irregular Development of Fusarium oxysporum f. sp. niveum Induced by Successive Subculture. Journal of Fungi, 6(4), 382. https://doi.org/10.3390/jof6040382