Haloadaptative Responses of Aspergillus sydowii to Extreme Water Deprivation: Morphology, Compatible Solutes, and Oxidative Stress at NaCl Saturation


 ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Strain, Preservation, and Culture Conditions
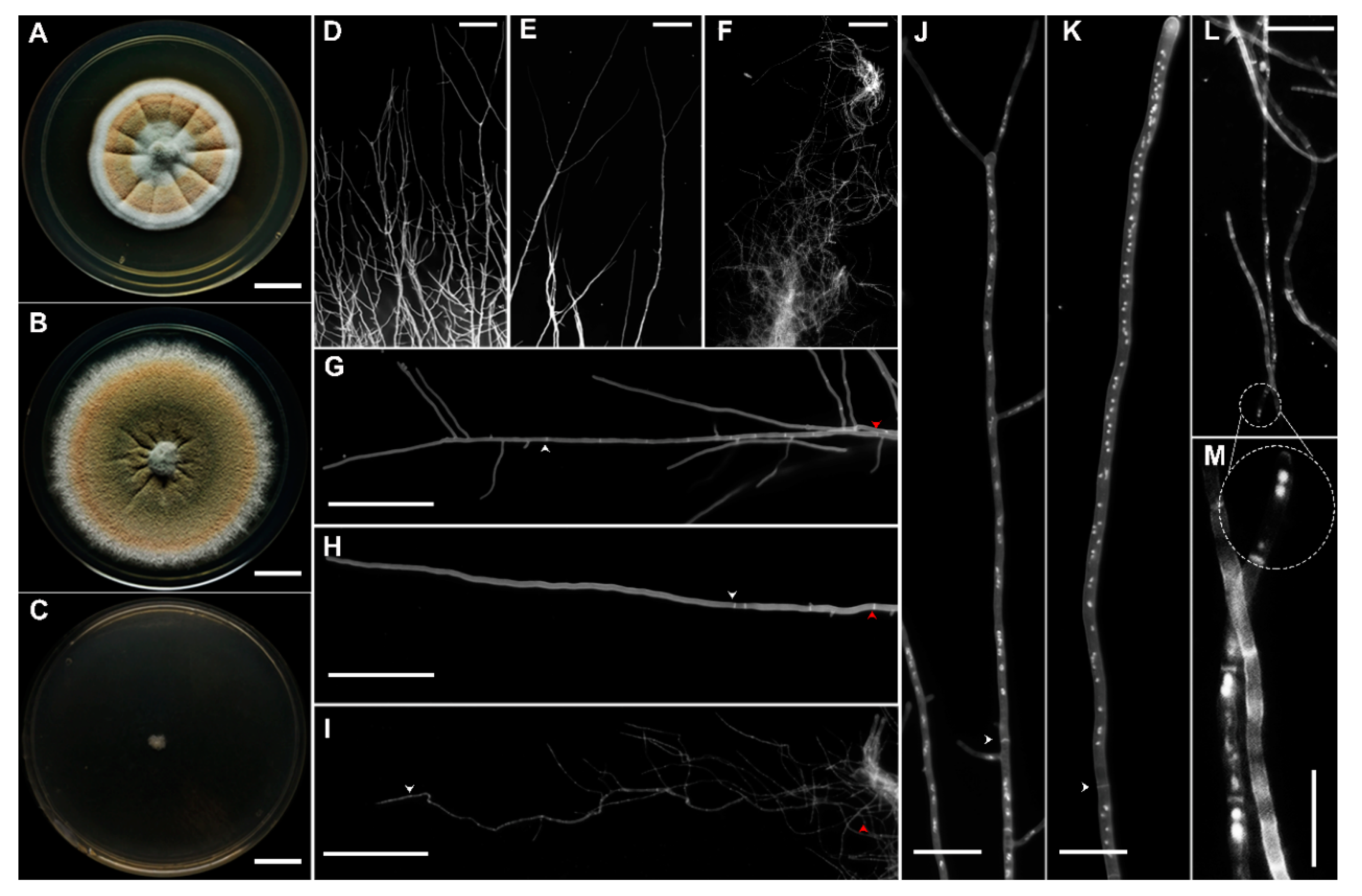
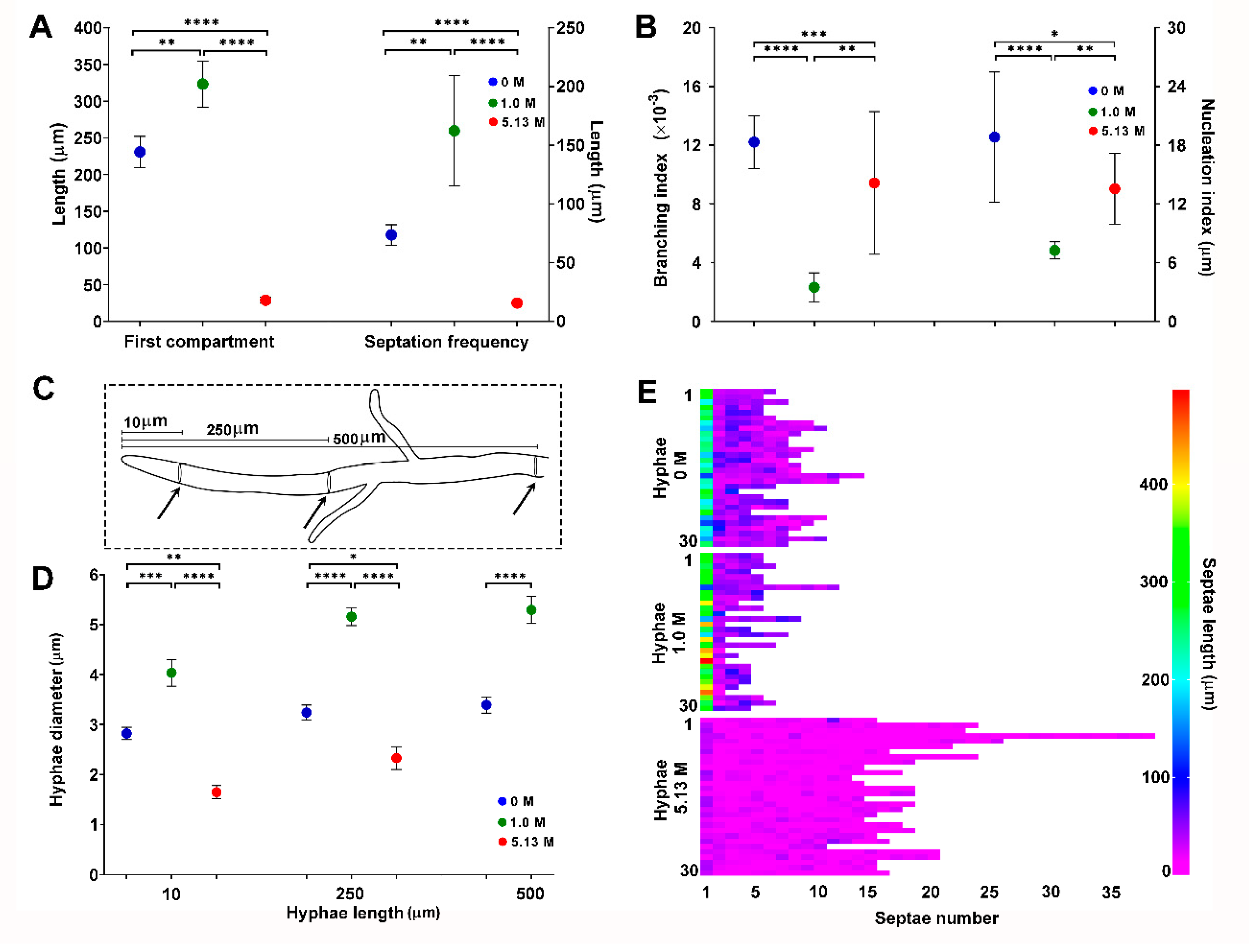
2.2. Morphological Analysis of Aspergillus sydowii EXF-12860 at Saturated NaCl Solution
2.3. Osmolytes in Aspergillus sydowii EXF-12860 Exposed to Saturated NaCl Concentration
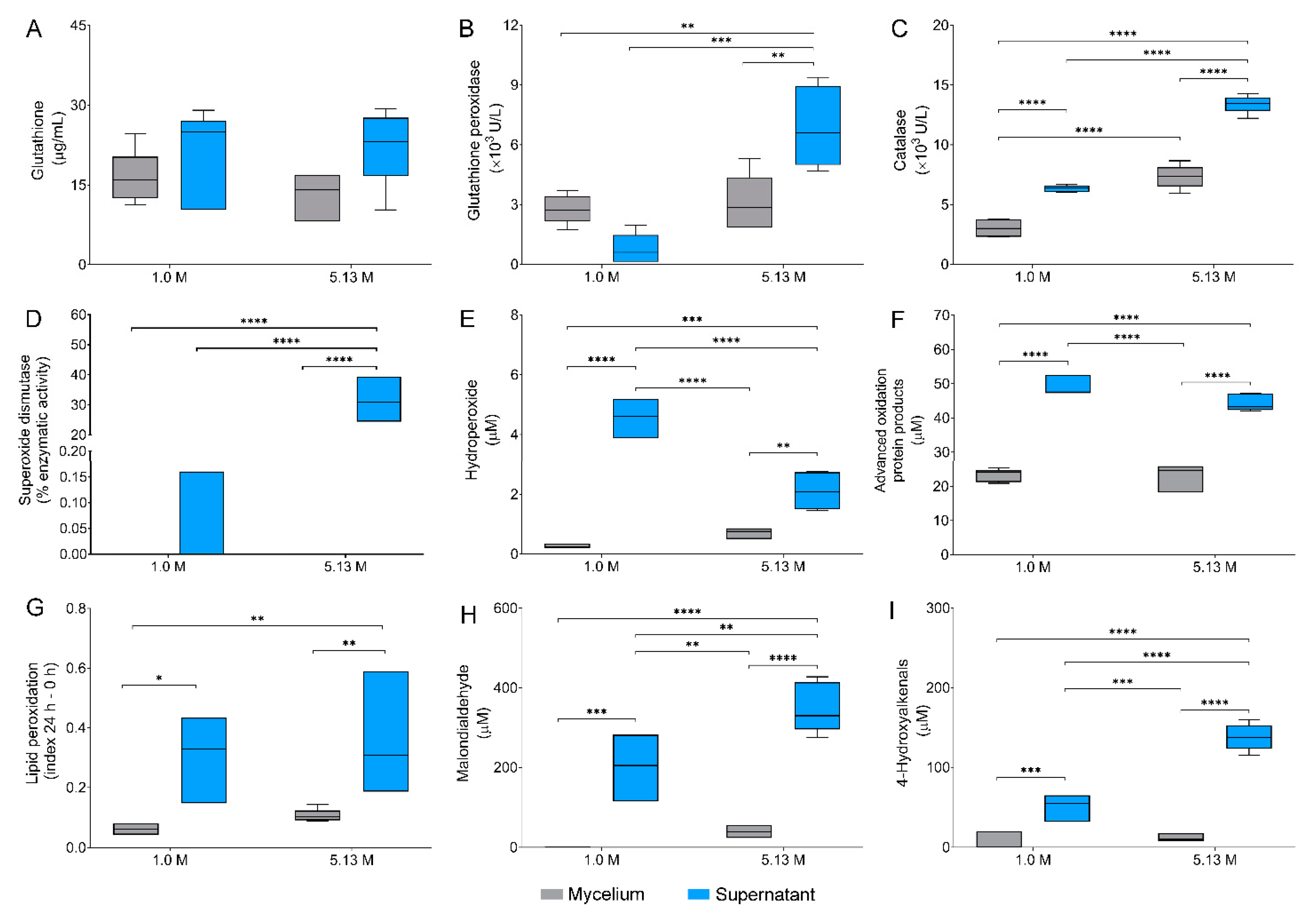
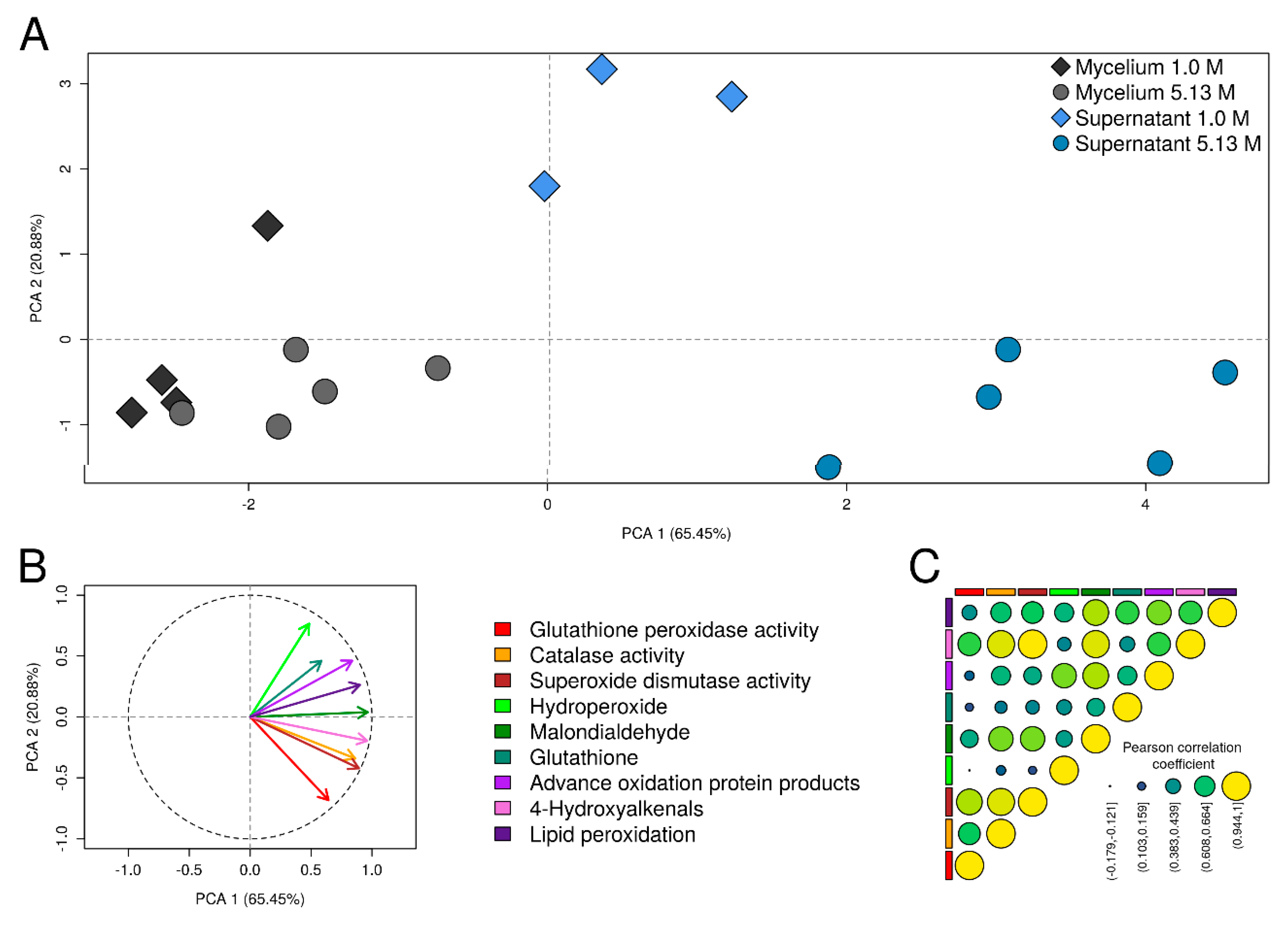
2.4. Oxidative Stress Biomarkers in Aspergillus sydowii EXF-12860 at Hypersaline Conditions
2.5. Transcriptomic Expression of Genes Involved in Cellular Oxidative Stress Defenses
2.6. Statistical Calculations
3. Results and Discussion
3.1. Morphology of Aspergillus sydowii EXF-12860 at Different NaCl Concentrations
3.2. Synthesis of Compatible Solutes by Aspergillus sydowii EXF-12860 Exposed to NaCl
3.3. Oxidative Stress Defenses Induced by NaCl in Aspergillus sydowii EXF-12860
3.4. Transcriptional Expression of Genes Related to Oxidative Stress Responses in Aspergillus sydowii EXF-12860
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Reid, D.S. Water activity: Fundamentals and Relationships. Water Activity in Foods: Fundamentals and Applications, 2nd ed.; Barbosa-Cánovas, G.V., Anthony, J.F., Jr., Schmidt, S.J., Labuza, T.P., Eds.; University of California-Davis: Davis, CA, USA, 2020; pp. 13–26. [Google Scholar]
- Ball, P.; Hallsworth, J.E. Water structure and chaotropicity: Their uses, abuses and biological implications. Phys. Chem. Chem. Phys. 2015, 17, 8297–8305. [Google Scholar] [CrossRef]
- Stevenson, A.; Burkhardt, J.; Cockell, C.S.; Cray, J.A.; Dijksterhuis, J.; Fox-Powell, M.; Kee, T.P.; Kminek, G.; McGenity, T.J.; Timmis, K.N. Multiplication of microbes below 0.690 water activity: Implications for terrestrial and extraterrestrial life. Environ. Microbiol. 2015, 17, 257–277. [Google Scholar] [CrossRef] [PubMed]
- Hallsworth, J.E. Stress-free microbes lack vitality. Fungal Biol. 2018, 122, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, A.; Cray, J.A.; Williams, J.P.; Santos, R.; Sahay, R.; Neuenkirchen, N.; McClure, C.D.; Grant, I.R.; Houghton, J.D.; Quinn, J.P. Is there a common water-activity limit for the three domains of life? ISME J. 2015, 9, 1333–1351. [Google Scholar] [CrossRef] [PubMed]
- Gunde-Cimerman, N.; Plemenitaš, A.; Oren, A. Strategies of adaptation of microorganisms of the three domains of life to high salt concentrations. FEMS Microbiol. Rev. 2018, 42, 353–375. [Google Scholar] [CrossRef] [PubMed]
- Kis-Papo, T.; Weig, A.R.; Riley, R.; Peršoh, D.; Salamov, A.; Sun, H.; Lipzen, A.; Wasser, S.P.; Rambold, G.; Grigoriev, I.V. Genomic adaptations of the halophilic Dead Sea filamentous fungus Eurotium rubrum. Nat. Commun. 2014, 5, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Plemenitaš, A.; Lenassi, M.; Konte, T.; Kejžar, A.; Zajc, J.; Gostinčar, C.; Gunde-Cimerman, N. Adaptation to high salt concentrations in halotolerant/halophilic fungi: A molecular perspective. Front. Microbiol. 2014, 5, 199. [Google Scholar] [CrossRef] [PubMed]
- Plemenitaš, A.; Vaupotič, T.; Lenassi, M.; Kogej, T.; Gunde-Cimerman, N. Adaptation of extremely halotolerant black yeast Hortaea werneckii to increased osmolarity: A molecular perspective at a glance. Studies Mycol. 2008, 61, 67–75. [Google Scholar]
- Zajc, J.; Liu, Y.; Dai, W.; Yang, Z.; Hu, J.; Gostinčar, C.; Gunde-Cimerman, N. Genome and transcriptome sequencing of the halophilic fungus Wallemia ichthyophaga: Haloadaptations present and absent. BMC Genom. 2013, 14, 1–21. [Google Scholar] [CrossRef]
- Ge, Y.; Yu, F.; Tan, Y.; Zhang, X.; Liu, Z. Comparative transcriptome sequence analysis of sporulation-related genes of Aspergillus cristatus in response to low and high osmolarity. Curr. Microbiol. 2017, 74, 806–814. [Google Scholar] [CrossRef]
- Ding, X.; Liu, K.; Lu, Y.; Gong, G. Morphological, transcriptional, and metabolic analyses of osmotic-adapted mechanisms of the halophilic Aspergillus montevidensis ZYD4 under hypersaline conditions. Appl. Microbiol. Biotechnol. 2019, 103, 3829–3846. [Google Scholar] [CrossRef] [PubMed]
- Tafer, H.; Poyntner, C.; Lopandic, K.; Sterflinger, K.; Piñar, G. Back to the Salt Mines: Genome and Transcriptome Comparisons of the Halophilic Fungus Aspergillus salisburgensis and Its Halotolerant Relative Aspergillus sclerotialis. Genes 2019, 10, 381. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Llano, Y.; Rodríguez-Pupo, E.C.; Druzhinina, I.S.; Chenthamara, K.; Cai, F.; Gunde-Cimerman, N.; Zalar, P.; Gostinčar, C.; Kostanjšek, R.; Folch-Mallol, J.L. Stress Reshapes the Physiological Response of Halophile Fungi to Salinity. Cells 2020, 9, 525. [Google Scholar] [CrossRef] [PubMed]
- Batista-García, R.A.; Balcázar-López, E.; Miranda-Miranda, E.; Sánchez-Reyes, A.; Cuervo-Soto, L.; Aceves-Zamudio, D.; Atriztán-Hernández, K.; Morales-Herrera, C.; Rodríguez-Hernández, R.; Folch-Mallol, J. Characterization of lignocellulolytic activities from a moderate halophile strain of Aspergillus caesiellus isolated from a sugarcane bagasse fermentation. PLoS ONE 2014, 9, e105893. [Google Scholar] [CrossRef] [PubMed]
- Hickey, P.C.; Jacobson, D.J.; Read, N.D.; Glass, N.L. Live-cell imaging of vegetative hyphal fusion in Neurospora crassa. Fungal Genet. Biol. 2002, 37, 109–119. [Google Scholar] [CrossRef]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef]
- Kogej, T.; Stein, M.; Volkmann, M.; Gorbushina, A.A.; Galinski, E.A.; Gunde-Cimerman, N. Osmotic adaptation of the halophilic fungus Hortaea werneckii: Role of osmolytes and melanization. Microbiology 2007, 153, 4261–4273. [Google Scholar] [CrossRef]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef]
- Ramasarma, T.; Rao, A.V.; Devi, M.M.; Omkumar, R.; Bhagyashree, K.; Bhat, S. New insights of superoxide dismutase inhibition of pyrogallol autoxidation. Mol. Cell Biochem. 2015, 400, 277–285. [Google Scholar] [CrossRef]
- Ecobichon, D. Glutathione depletion and resynthesis in laboratory animals. Drug Chem. Toxicoly 1984, 7, 345–355. [Google Scholar] [CrossRef]
- Witko, V.; Nguyen, A.T.; Descamps-Latscha, B. Microtiter plate assay for phagocyte-derived Taurine-chloramines. J. Clin. Lab. Anal. 1992, 6, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Erdelmeier, I.; Gérard-Monnier, D.; Yadan, J.-C.; Chaudiere, J. Reactions of N-methyl-2-phenylindole with malondialdehyde and 4-hydroxyalkenals. Mechanistic aspects of the colorimetric assay of lipid peroxidation. Chem. Res. Toxicol. 1998, 11, 1184–1194. [Google Scholar] [CrossRef] [PubMed]
- Ozdemirler, G.; Mehmetcik, G.; Oztezcan, S.; Toker, G.; Sivas, A.; Uysal, M. Peroxidation potential and antioxidant activity of serum in patients with diabetes mellitus and myocard infarction. Horm. Metab. Res. 1995, 27, 194–196. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Song, L.; Florea, L. Rcorrector: Efficient and accurate error correction for Illumina RNA-seq reads. Gigascience 2015, 4, 48. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Kunčič, M.K.; Kogej, T.; Drobne, D.; Gunde-Cimerman, N. Morphological response of the halophilic fungal genus Wallemia to high salinity. Appl. Environ. Microbiol. 2010, 76, 329–337. [Google Scholar] [CrossRef]
- Liu, K.-H.; Ding, X.-W.; Narsing Rao, M.P.; Zhang, B.; Zhang, Y.-G.; Liu, F.-H.; Liu, B.-B.; Xiao, M.; Li, W.-J. Morphological and transcriptomic analysis reveals the osmoadaptive response of endophytic fungus Aspergillus montevidensis ZYD4 to high salt stress. Front. Microbiol. 2017, 8, 1789. [Google Scholar] [CrossRef]
- Matsuda, Y.; Sugiyama, F.; Nakanishi, K.; Ito, S.-i. Effects of sodium chloride on growth of ectomycorrhizal fungal isolates in culture. Mycoscience 2006, 47, 212–217. [Google Scholar] [CrossRef]
- Kelavkar, U.; Rao, K.S.; Ghhatpar, H.S. Sodium chloride stress induced morphological and ultrastructural changes in Aspergillus repens. Indian J. Exp. Biol. 1993, 31, 511–515. [Google Scholar]
- Ide-Pérez, M.R.; Fernández-López, M.G.; Sánchez-Reyes, A.; Leija, A.; Batista-García, R.A.; Folch-Mallol, J.L.; Sánchez-Carbente, M.d.R. Aromatic Hydrocarbon Removal by Novel Extremotolerant Exophiala and Rhodotorula Spp. from an Oil Polluted Site in Mexico. J. Fungi. 2020, 6, 135. [Google Scholar] [CrossRef] [PubMed]
- Gladfelter, A.S. Control of filamentous fungal cell shape by septins and formins. Nat. Rev. Microbiol. 2006, 4, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Gladfelter, A.S. Guides to the final frontier of the cytoskeleton: Septins in filamentous fungi. Curr. Opin. Microbiol. 2010, 13, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Kozubowski, L.; Heitman, J. Septins enforce morphogenetic events during sexual reproduction and contribute to virulence of Cryptococcus neoformans. Mol. Microbiol. 2010, 75, 658–675. [Google Scholar] [CrossRef] [PubMed]
- Lindsey, R.; Cowden, S.; Hernández-Rodríguez, Y.; Momany, M. Septins AspA and AspC are important for normal development and limit the emergence of new growth foci in the multicellular fungus Aspergillus nidulans. Eukaryot. Cell 2010, 9, 155–163. [Google Scholar] [CrossRef]
- Lindsey, R.; Ha, Y.; Momany, M. A septin from the filamentous fungus A. nidulans induces atypical pseudohyphae in the budding yeast S. cerevisiae. PLoS ONE 2010, 5, e9858. [Google Scholar] [CrossRef]
- Vargas-Muñiz, J.M.; Renshaw, H.; Richards, A.D.; Lamoth, F.; Soderblom, E.J.; Moseley, M.A.; Juvvadi, P.R.; Steinbach, W.J. The Aspergillus fumigatus septins play pleiotropic roles in septation, conidiation, and cell wall stress, but are dispensable for virulence. Fungal Genet. Biol. 2015, 81, 41–51. [Google Scholar] [CrossRef][Green Version]
- Zajc, J.; Kogej, T.; Galinski, E.A.; Ramos, J.; Gunde-Cimerman, N. Osmoadaptation strategy of the most halophilic fungus, Wallemia ichthyophaga, growing optimally at salinities above 15% NaCl. Appl. Environ. Microbiol. 2014, 80, 247–256. [Google Scholar] [CrossRef]
- Turk, M.; Gostinčar, C. Glycerol metabolism genes in Aureobasidium pullulans and Aureobasidium subglaciale. Fungal Biol. 2018, 122, 63–73. [Google Scholar] [CrossRef]
- Krijgsheld, P.; Bleichrodt, R.; van Veluw, G.J.; Wang, F.; Müller, W.H.; Dijksterhuis, J.; Wösten, H.A. Development in Aspergillus. Studies Mycol. 2013, 74, 1–29. [Google Scholar] [CrossRef]
- Witteveen, C.F.; Visser, J. Polyol pools in Aspergillus niger. FEMS Microbiol. Lett. 1995, 134, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Gostinčar, C.; Gunde-Cimerman, N. Overview of oxidative stress response genes in selected halophilic fungi. Genes 2018, 9, 143. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.H.A.; Alkhalifah, D.H.M.; Al Yousef, S.A.; Beemster, G.T.S.; Mousa, A.S.M.; Hozzein, W.N.; AbdElgawad, H. Salinity Stress Enhances the Antioxidant Capacity of Bacillus and Planococcus Species Isolated From Saline Lake Environment. Front. Microbiol. 2020, 11, 561816. [Google Scholar] [CrossRef] [PubMed]
- Sreenivasulu, N.; Miranda, M.; Prakash, H.S.; Wobus, U.; Weschke, W. Transcriptome changes in foxtail millet genotypes at high salinity: Identification and characterization of a PHGPX gene specifically up-regulated by NaCl in a salt-tolerant line. J. Plant Physiol. 2004, 161, 467–477. [Google Scholar] [CrossRef]
- Petrovic, U. Role of oxidative stress in the extremely salt-tolerant yeast Hortaea werneckii. FEMS Yeast Res. 2006, 6, 816–822. [Google Scholar] [CrossRef][Green Version]
- Warris, A.; Ballou, E.R. Oxidative responses and fungal infection biology. Semin. Cell Dev. Biol. 2019, 89, 34–46. [Google Scholar] [CrossRef]
- Osman, M.S.; Badawy, A.A.; Osman, A.I.; Abdel Latef, A.A.H. Ameliorative Impact of an Extract of the Halophyte Arthrocnemum macrostachyum on Growth and Biochemical Parameters of Soybean Under Salinity Stress. J. Plant Growth Regul. 2020. [Google Scholar] [CrossRef]
- Vaupotic, T.; Veranic, P.; Jenoe, P.; Plemenitas, A. Mitochondrial mediation of environmental osmolytes discrimination during osmoadaptation in the extremely halotolerant black yeast Hortaea werneckii. Fungal Genet. Biol. 2008, 45, 994–1007. [Google Scholar] [CrossRef]
- Morgan, B.; Ezeriņa, D.; Amoako, T.N.; Riemer, J.; Seedorf, M.; Dick, T.P. Multiple glutathione disulfide removal pathways mediate cytosolic redox homeostasis. Nat. Chem. Biol. 2013, 9, 119–125. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef]
- Busch, C.J.; Binder, C.J. Malondialdehyde epitopes as mediators of sterile inflammation. Biochim Biophys Acta Mol. Cell Biol. Lipids 2017, 1862, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Dobarganes, M.C.; Velasco, J. Analysis of lipid hydroperoxides. Eur. J. Lipid Sci. Technol. 2002, 104, 420–428. [Google Scholar] [CrossRef]
- Sasson, S. 4-Hydroxyalkenal-activated PPARδ complexes mediate glucohormetic and lipohormetic interactions in diabetes. Free. Radical. Biol. Med. 2018, 124, 560. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, Q.; Xi, B. Effect of NaCl salinity on the growth, metabolites, and antioxidant system of Microcystis aeruginosa. J. Freshw. Ecol. 2013, 28, 477–487. [Google Scholar] [CrossRef]
- Finkel, T. From sulfenylation to sulfhydration: What a thiolate needs to tolerate. Sci. Signal 2012, 5, pe10. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C.C.; Hampton, M.B. Thiol chemistry and specificity in redox signaling. Free. Radic. Biol. Med. 2008, 45, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Sarıkaya, E.; Doğan, S. Glutathione Peroxidase in Health and Diseases. In Glutathione Peroxidase in Health and Disease; IntechOpen: London, UK, 2020. [Google Scholar]
- Aktaş, M.; Değirmenci, U.; Ercan, S.K.; Tamer, L.; y Atik, U.; Aktaþ, M.; Deðirmenci, U.; Ercan, S.K.; Tamer, L.; Atik, U. The Comparison of Spectrophotometric and HPLC Methods in Reduced Glutathione Measurements. Türk Klinik Biyokimya Derg 2005, 3, 95–99. [Google Scholar]
- Calabrese, G.; Morgan, B.; Riemer, J. Mitochondrial Glutathione: Regulation and Functions. Antioxid Redox Signal 2017, 27, 1162–1177. [Google Scholar] [CrossRef]
- Couto, N.; Wood, J.; Barber, J. The role of glutathione reductase and related enzymes on cellular redox homoeostasis network. Free. Radic. Biol. Med. 2016, 95, 27–42. [Google Scholar] [CrossRef]
- Fujino, G.; Noguchi, T.; Takeda, K.; Ichijo, H. Thioredoxin and protein kinases in redox signaling. Semin Cancer Biol. 2006, 16, 427–435. [Google Scholar] [CrossRef]
- Matsuzawa, A. Thioredoxin and redox signaling: Roles of the thioredoxin system in control of cell fate. Arch. Biochem. Biophys. 2017, 617, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Liebthal, M.; Maynard, D.; Dietz, K.J. Peroxiredoxins and Redox Signaling in Plants. Antioxid Redox Signal 2018, 28, 609–624. [Google Scholar] [CrossRef] [PubMed]
- Esse, R.; Barroso, M.; Tavares de Almeida, I.; Castro, R. The Contribution of Homocysteine Metabolism Disruption to Endothelial Dysfunction: State-of-the-Art. Int. J. Mol. Sci. 2019, 20, 867. [Google Scholar] [CrossRef] [PubMed]
- Richard, E.; Gallego-Villar, L.; Rivera-Barahona, A.; Oyarzábal, A.; Pérez, B.; Rodríguez-Pombo, P.; Desviat, L.R. Altered Redox Homeostasis in Branched-Chain Amino Acid Disorders, Organic Acidurias, and Homocystinuria. Oxid. Med. Cell Longev. 2018, 2018, 1246069. [Google Scholar] [CrossRef] [PubMed]
- Taysi, S.; Keles, M.S.; Gumustekin, K.; Akyuz, M.; Boyuk, A.; Cikman, O.; Bakan, N. Plasma homocysteine and liver tissue S-adenosylmethionine, S-adenosylhomocysteine status in vitamin B6-deficient rats. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 154–160. [Google Scholar] [PubMed]
- Sankaranarayanan, S.; Jamshed, M.; Kumar, A.; Skori, L.; Scandola, S.; Wang, T.; Spiegel, D.; Samuel, M.A. Glyoxalase Goes Green: The Expanding Roles of Glyoxalase in Plants. Int. J. Mol. Sci. 2017, 18, 898. [Google Scholar] [CrossRef] [PubMed]
- Kaur, C.; Ghosh, A.; Pareek, A.; Sopory, S.K.; Singla-Pareek, S.L. Glyoxalases and stress tolerance in plants. Biochem. Soc. Trans. 2014, 42, 485–490. [Google Scholar] [CrossRef]
- Thornalley, P.J. The glyoxalase system: New developments towards functional characterization of a metabolic pathway fundamental to biological life. Biochem. J. 1990, 269, 1–11. [Google Scholar] [CrossRef]
- Crook, E.M.; Law, K. Glyoxalase; the role of the components. Biochem. J. 1952, 52, 492–499. [Google Scholar] [CrossRef]
- Chutoam, P.; Charoensawan, V.; Wongtrakoongate, P.; Kum-Arth, A.; Buphamalai, P.; Tungpradabkul, S. RpoS and oxidative stress conditions regulate succinyl-CoA: 3-ketoacid-coenzyme A transferase (SCOT) expression in Burkholderia pseudomallei. Microbiol. Immunol. 2013, 57, 605–615. [Google Scholar] [CrossRef]
- Folgueira, I.; Lamas, J.; Sueiro, R.A.; Leiro, J.M. Molecular characterization and gene expression modulation of the alternative oxidase in a scuticociliate parasite by hypoxia and mitochondrial respiration inhibitors. Sci. Rep. 2020, 10, 11880. [Google Scholar] [CrossRef] [PubMed]
- McDonald, A.E. Alternative oxidase: An inter-kingdom perspective on the function and regulation of this broadly distributed ’cyanide-resistant’ terminal oxidase. Funct. Plant Biol. 2008, 35, 535–552. [Google Scholar] [CrossRef] [PubMed]
- Barsottini, M.R.O.; Copsey, A.; Young, L.; Baroni, R.M.; Cordeiro, A.T.; Pereira, G.A.G.; Moore, A.L. Biochemical characterization and inhibition of the alternative oxidase enzyme from the fungal phytopathogen Moniliophthora perniciosa. Commun. Biol. 2020, 3, 263. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Wu, J.; Jamieson, P.A.; Zhang, C. Alternative Oxidase Is Involved in the Pathogenicity, Development, and Oxygen Stress Response of Botrytis cinerea. Phytopathology 2019, 109, 1679–1688. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Li, M.; Cao, Y.; Gao, P.; Cao, Y.; Wang, Y.; Jiang, Y. The alternative oxidase of Candida albicans causes reduced fluconazole susceptibility. J. Antimicrob. Chemother. 2009, 64, 764–773. [Google Scholar] [CrossRef]
- Grahl, N.; Dinamarco, T.M.; Willger, S.D.; Goldman, G.H.; Cramer, R.A. Aspergillus fumigatus mitochondrial electron transport chain mediates oxidative stress homeostasis, hypoxia responses and fungal pathogenesis. Mol. Microbiol. 2012, 84, 383–399. [Google Scholar] [CrossRef]
- Kowalski, C.H.; Beattie, S.R.; Fuller, K.K.; McGurk, E.A.; Tang, Y.W.; Hohl, T.M.; Obar, J.J.; Cramer, R.A., Jr. Heterogeneity among Isolates Reveals that Fitness in Low Oxygen Correlates with Aspergillus fumigatus Virulence. mBio 2016, 7. [Google Scholar] [CrossRef]
- Honda, Y.; Hattori, T.; Kirimura, K. Visual expression analysis of the responses of the alternative oxidase gene (aox1) to heat shock, oxidative, and osmotic stresses in conidia of citric acid-producing Aspergillus niger. J. Biosci. Bioeng. 2012, 113, 338–342. [Google Scholar] [CrossRef]
- Yukioka, H.; Inagaki, S.; Tanaka, R.; Katoh, K.; Miki, N.; Mizutani, A.; Masuko, M. Transcriptional activation of the alternative oxidase gene of the fungus Magnaporthe grisea by a respiratory-inhibiting fungicide and hydrogen peroxide. Biochim. Biophys. Acta 1998, 1442, 161–169. [Google Scholar] [CrossRef]
- Minagawa, N.; Koga, S.; Nakano, M.; Sakajo, S.; Yoshimoto, A. Possible involvement of superoxide anion in the induction of cyanide-resistant respiration in Hansenula anomala. FEBS Lett. 1992, 302, 217–219. [Google Scholar] [CrossRef]
- Martins, V.P.; Dinamarco, T.M.; Soriani, F.M.; Tudella, V.G.; Oliveira, S.C.; Goldman, G.H.; Curti, C.; Uyemura, S.A. Involvement of an alternative oxidase in oxidative stress and mycelium-to-yeast differentiation in Paracoccidioides brasiliensis. Eukaryot. Cell 2011, 10, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Bulteau, A.L.; Ikeda-Saito, M.; Szweda, L.I. Redox-dependent modulation of aconitase activity in intact mitochondria. Biochemistry 2003, 42, 14846–14855. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
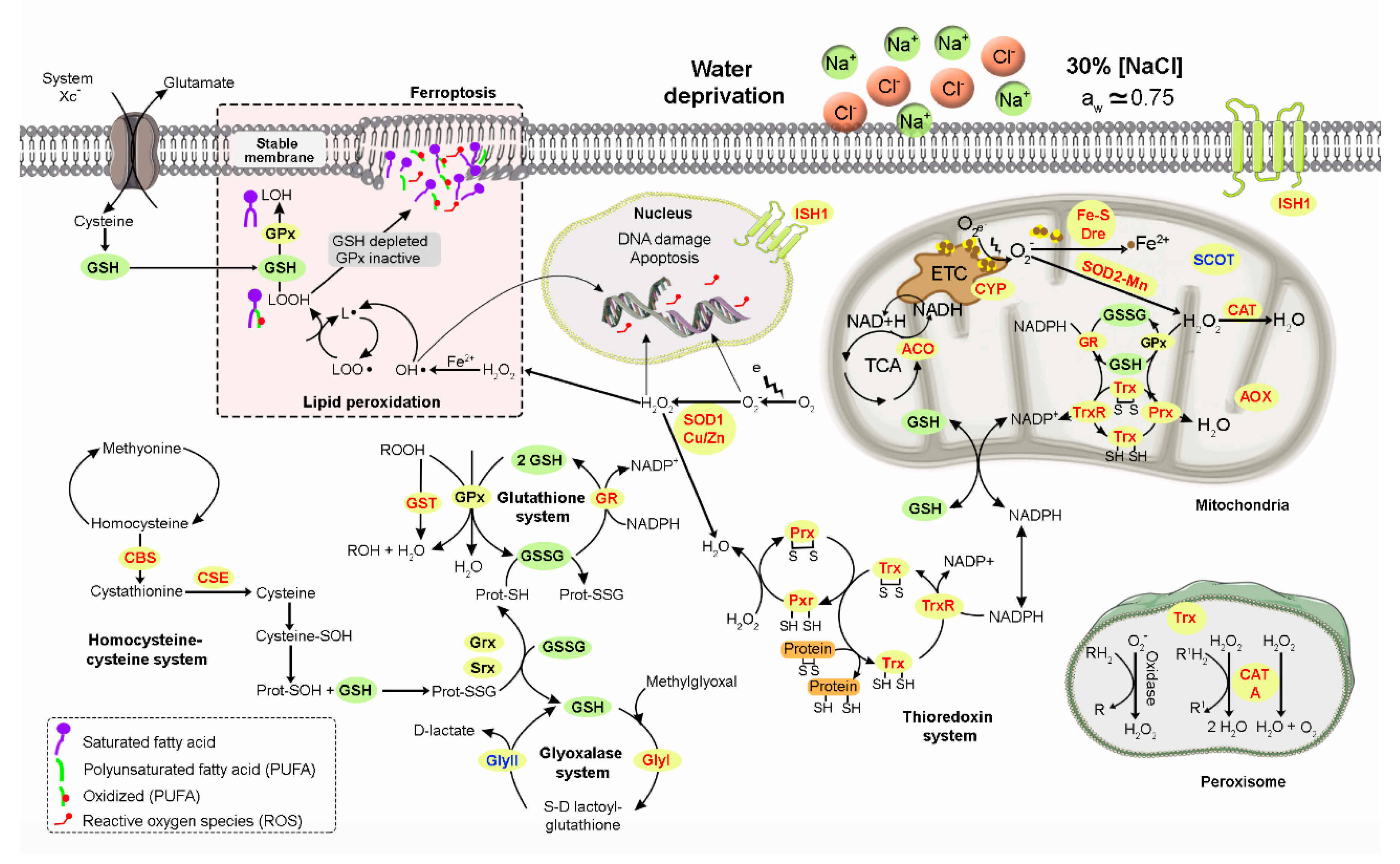{kind=link}
| Transcript ID | Annotation | Gene ID | LogFC | FDR |
|---|---|---|---|---|
| TRINITY_DN892_c0_g1_i12 | Catalase A | catA | 11.27 | 4.46 × 10−4 |
| TRINITY_DN892_c0_g1_i4 | 10.73 | 6.37 × 10−4 | ||
| TRINITY_DN892_c0_g1_i3 | −4.12 | 2.57 × 10−3 | ||
| TRINITY_DN892_c0_g1_i6 | −5.08 | 2.69 × 10−4 | ||
| TRINITY_DN892_c0_g1_i11 | −11.27 | 2.46 × 10−8 | ||
| TRINITY_DN892_c0_g1_i9 | −11.94 | 2.53 × 10−9 | ||
| TRINITY_DN892_c0_g1_i10 | −14.33 | 1.37 × 10−11 | ||
| TRINITY_DN1037_c0_g1_i5 | Glyoxalase I | gly1 | 11.15 | 4.82 × 10−4 |
| TRINITY_DN1037_c0_g1_i4 | 10.40 | 9.77 × 10−4 | ||
| TRINITY_DN2716_c0_g2_i1 | 3.85 | 5.32 × 10−2 | ||
| TRINITY_DN1197_c0_g1_i1 | Cystathionine gamma-lyase | cth | 10.51 | 8.67 × 10−4 |
| TRINITY_DN1197_c0_g1_i7 | 7.20 | 3.63 × 10−6 | ||
| TRINITY_DN1197_c0_g1_i6 | 3.66 | 8.26 × 10−3 | ||
| TRINITY_DN2112_c0_g1_i19 | Cytochrome P450 alkane hydroxylase | cyp | 9.84 | 9.10 × 10−7 |
| TRINITY_DN3588_c0_g1_i1 | Cytochrome P450 | cyp450 | 6.65 | 4.76 × 10−6 |
| TRINITY_DN2010_c0_g1_i16 | 4.89 | 3.88 × 10−4 | ||
| TRINITY_DN2010_c0_g1_i10 | 4.65 | 7.22 × 10−4 | ||
| TRINITY_DN312_c0_g1_i9 | −4.57 | 1.59 × 10−3 | ||
| TRINITY_DN312_c0_g1_i2 | −3.67 | 8.32 × 10−3 | ||
| TRINITY_DN4029_c0_g1_i2 | Aconitate hydratase | aco1 | 8.88 | 7.12 × 10−6 |
| TRINITY_DN1574_c0_g1_i2 | Alternative oxidase-domain-containing protein | aox | 5.25 | 1.47 × 10−4 |
| TRINITY_DN1793_c0_g1_i1 | Fe-S cluster assembly protein dre2 | dre2 | 5.24 | 1.51 × 10−4 |
| TRINITY_DN10053_c1_g1_i1 | Peroxirredoxin 6 | prdx6 | 5.20 | 1.89 × 10−4 |
| TRINITY_DN2598_c0_g1_i4 | Cytosolic Cu/Zn superoxide dismutase | sod1 | 5.01 | 2.74 × 10−4 |
| TRINITY_DN2598_c0_g1_i3 | 4.38 | 1.66 × 10−3 | ||
| TRINITY_DN1168_c0_g2_i5 | Stress response protein putative | ish1 | 4.84 | 8.67 × 10−4 |
| TRINITY_DN681_c0_g1_i14 | Glutaredoxin/glutathione-dependent peroxiredoxin | prx1 | 4.73 | 7.03 × 10−4 |
| TRINITY_DN1443_c0_g1_i3 | Tiorredoxin-like protein | trx | 4.39 | 1.36 × 10−3 |
| TRINITY_DN1443_c0_g1_i3 | −5.45 | 2.93 × 10−4 | ||
| TRINITY_DN2569_c2_g2_i1 | Glutathione S-transferase | gst | 4.19 | 2.90 × 10−3 |
| TRINITY_DN9329_c0_g1_i1 | −4.15 | 5.47 × 10−3 | ||
| TRINITY_DN473_c0_g1_i3 | −10.53 | 1.67 × 10−7 | ||
| TRINITY_DN902_c0_g1_i5 | Superoxide dismutase [Mn] mitochondrial | sod2 | 4.21 | 3.20 × 10−3 |
| TRINITY_DN902_c0_g1_i4 | Superoxide dismutase Fe-Mn family | sod2 | 3.65 | 7.90 × 10−3 |
| TRINITY_DN2375_c0_g1_i2 | MAP kinase kinase kinase | ask1 | −3.80 | 5.87 × 10−3 |
| TRINITY_DN38_c0_g1_i5 | Homocysteine synthase | cysD | −3.84 | 7.67 × 10−3 |
| TRINITY_DN50_c0_g1_i16 | −6.25 | 8.00 × 10−5 | ||
| TRINITY_DN1899_c0_g1_i4 | Succinyl-CoA:3-ketoacid-coenzyme A transferase | scot | −4.23 | 2.02 × 10−3 |
| TRINITY_DN1899_c0_g1_i2 | −5.14 | 2.47 × 10−4 | ||
| TRINITY_DN1899_c0_g1_i5 | −9.54 | 1.81 × 10−6 | ||
| TRINITY_DN1899_c0_g1_i6 | −9.57 | 1.71 × 10−6 | ||
| TRINITY_DN298_c0_g1_i11 | Glyoxalase II (Hydroxyacylglutathione hydrolase) | gly2 | −4.45 | 1.78 × 10−3 |
| TRINITY_DN1341_c0_g1_i2 | NAD(P)H dehydrogenase (quinone) | ipda | −8.91 | 9.70 × 10−6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiménez-Gómez, I.; Valdés-Muñoz, G.; Moreno-Perlin, T.; Mouriño-Pérez, R.R.; Sánchez-Carbente, M.d.R.; Folch-Mallol, J.L.; Pérez-Llano, Y.; Gunde-Cimerman, N.; Sánchez, N.d.C.; Batista-García, R.A. Haloadaptative Responses of Aspergillus sydowii to Extreme Water Deprivation: Morphology, Compatible Solutes, and Oxidative Stress at NaCl Saturation. J. Fungi 2020, 6, 316. https://doi.org/10.3390/jof6040316
Jiménez-Gómez I, Valdés-Muñoz G, Moreno-Perlin T, Mouriño-Pérez RR, Sánchez-Carbente MdR, Folch-Mallol JL, Pérez-Llano Y, Gunde-Cimerman N, Sánchez NdC, Batista-García RA. Haloadaptative Responses of Aspergillus sydowii to Extreme Water Deprivation: Morphology, Compatible Solutes, and Oxidative Stress at NaCl Saturation. Journal of Fungi. 2020; 6(4):316. https://doi.org/10.3390/jof6040316
Chicago/Turabian StyleJiménez-Gómez, Irina, Gisell Valdés-Muñoz, Tonatiuh Moreno-Perlin, Rosa R. Mouriño-Pérez, María del Rayo Sánchez-Carbente, Jorge Luis Folch-Mallol, Yordanis Pérez-Llano, Nina Gunde-Cimerman, Nilda del C. Sánchez, and Ramón Alberto Batista-García. 2020. "Haloadaptative Responses of Aspergillus sydowii to Extreme Water Deprivation: Morphology, Compatible Solutes, and Oxidative Stress at NaCl Saturation" Journal of Fungi 6, no. 4: 316. https://doi.org/10.3390/jof6040316
APA StyleJiménez-Gómez, I., Valdés-Muñoz, G., Moreno-Perlin, T., Mouriño-Pérez, R. R., Sánchez-Carbente, M. d. R., Folch-Mallol, J. L., Pérez-Llano, Y., Gunde-Cimerman, N., Sánchez, N. d. C., & Batista-García, R. A. (2020). Haloadaptative Responses of Aspergillus sydowii to Extreme Water Deprivation: Morphology, Compatible Solutes, and Oxidative Stress at NaCl Saturation. Journal of Fungi, 6(4), 316. https://doi.org/10.3390/jof6040316