Unravelling Diaporthe Species Associated with Woody Hosts from Karst Formations (Guizhou) in China



Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation of Fungal Material, Morphology and Culture Characteristics
2.2. Molecular Based Amplification
2.3. Sequence Alignment and Phylogenetic Analyses
3. Results
3.1. Phylogenetic Analyses

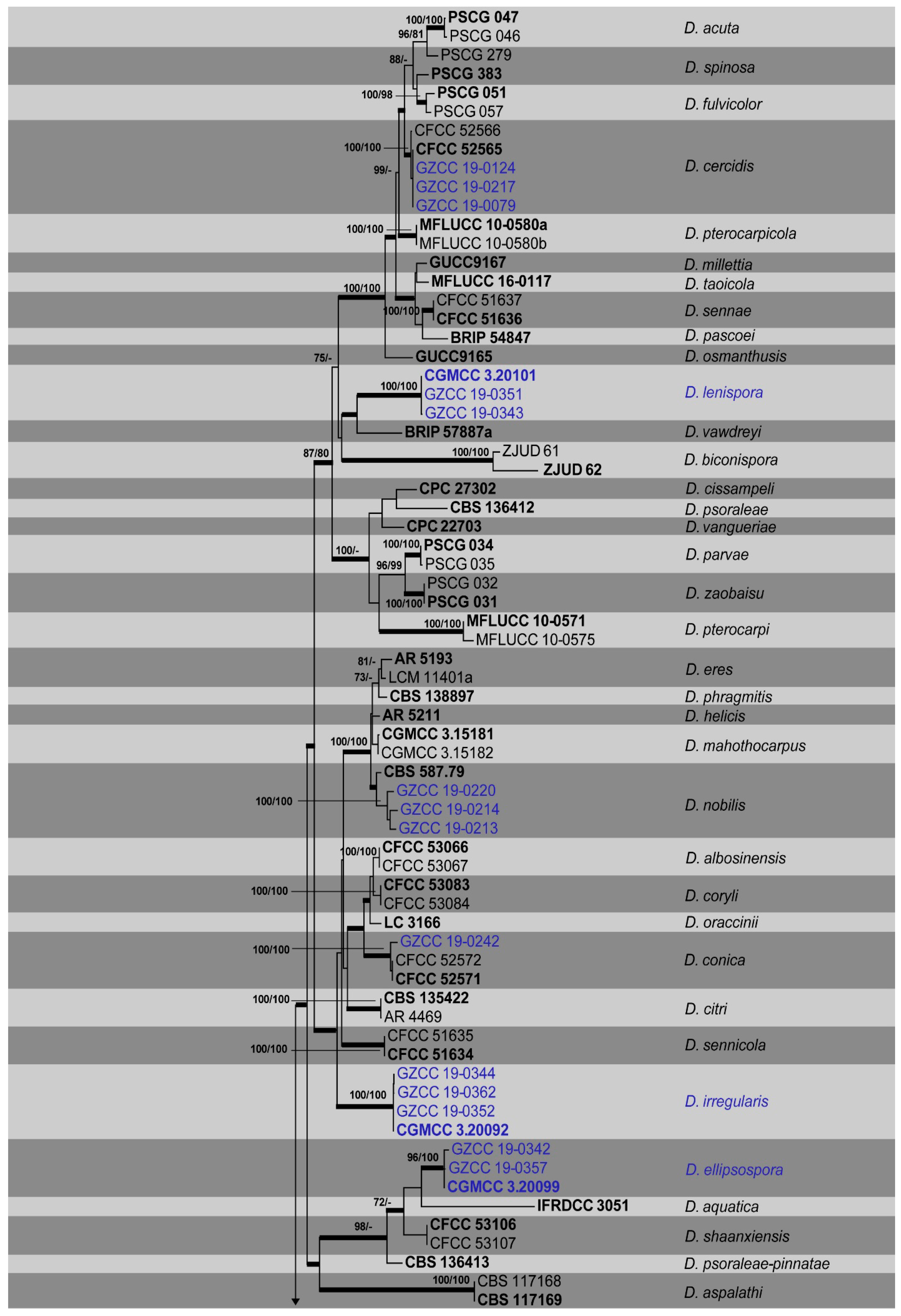{kind=link}
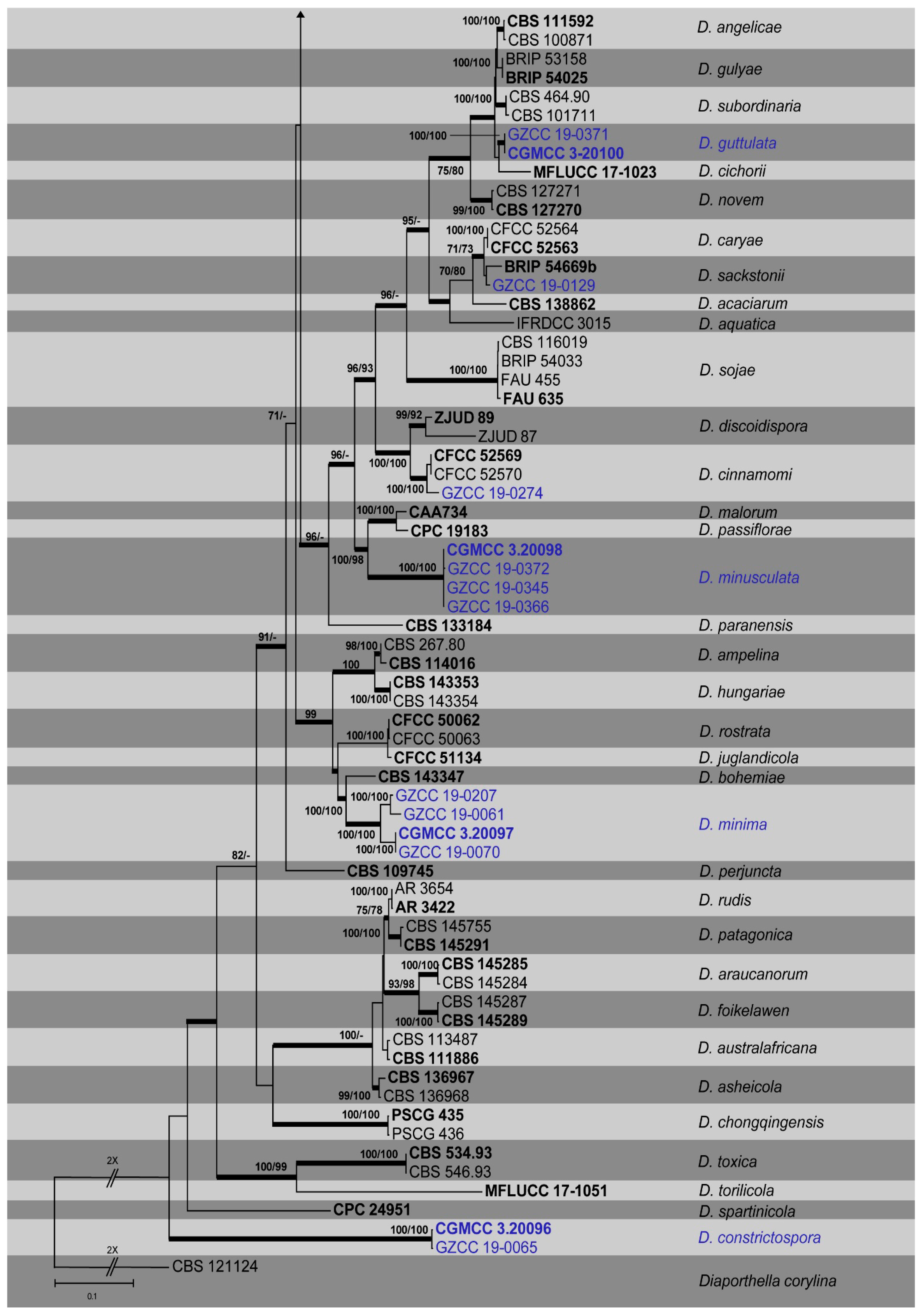{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Isolate Number | ITS | tub | tef | cal | his | Reference |
|---|---|---|---|---|---|---|---|
| Diaporthella corylina | CBS 121124 | KC343004 | KC343972 | KC343730 | KC343246 | KC343488 | Vasilyeva et al. [54] |
| Diaporthe acaciarum | CBS 138862 | KP004460 | KP004509 | N/A | N/A | KP004504 | Crous et al. [55] |
| Diaporthe acuta | PSCG 047 | MK626957 | MK691225 | MK654802 | MK691125 | MK726161 | Guo et al. [15] |
| Diaporthe acuta | PSCG 046 | MK626958 | MK691224 | MK654803 | MK691124 | MK726162 | Guo et al. [15] |
| Diaporthe albosinensis | CFCC 53066 | MK432659 | MK578059 | MK578133 | MK442979 | MK443004 | Yang et al. [14] |
| Diaporthe albosinensis | CFCC 53067 | MK432660 | MK578060 | MK578134 | MK442980 | MK443005 | Yang et al. [14] |
| Diaporthe ampelina | CBS 114016 | AF230751 | JX275452 | AY745056 | AY230751 | N/A | Mostert et al. [56] |
| Diaporthe ampelina | CBS 267.80 | KC343018 | KC343986 | KC343744 | KC343260 | KC343502 | Mostert et al. [56] |
| Diaporthe angelicae | CBS 111592 | KC343027 | KC343995 | KC343753 | KC343269 | KC343511 | Castlebury et al. [57] |
| Diaporthe angelicae | CBS 100871 | KC343025 | KC343993 | KC343751 | KC343267 | KC343509 | Castlebury et al. [57] |
| Diaporthe aquatica | IFRDCC 3015 | JQ797438 | N/A | N/A | N/A | N/A | Hu et al. [58] |
| Diaporthe aquatica | IFRDCC 3051 | JQ797437 | N/A | N/A | N/A | N/A | Hu et al. [58] |
| Diaporthe araucanorum | CBS 145285 | MN509711 | MN509722 | MN509733 | N/A | N/A | Zapata et al. [45] |
| Diaporthe araucanorum | CBS 145284 | MN509710 | MN509721 | MN509732 | N/A | N/A | Zapata et al. [45] |
| Diaporthe asheicola | CBS 136967 | KJ160562 | KJ160518 | KJ160594 | KJ160542 | N/A | Lombard et al. [59] |
| Diaporthe asheicola | CBS 136968 | KJ160563 | KJ160519 | KJ160595 | KJ160543 | N/A | Lombard et al. [59] |
| Diaporthe aspalathi | CBS 117168 | KC343035 | KC344003 | KC343761 | KC343277 | KC343519 | van Rensburg et al. [11] |
| Diaporthe aspalathi | CBS 117169 | KC343036 | KC344004 | KC343762 | KC343278 | KC343520 | van Rensburg et al. [11] |
| Diaporthe australafricana | CBS 111886 | KC343038 | KC344006 | KC343764 | KC343280 | KC343522 | Mostert et al. [56] |
| Diaporthe australafricana | CBS 113487 | KC343039 | KC344007 | KC343765 | KC343281 | KC343523 | Mostert et al. [56] |
| Diaporthe biconispora | ZJUD61 | KJ490596 | KJ490417 | KJ490475 | N/A | KJ490538 | Huang et al. [60] |
| Diaporthe biconispora | ZJUD62 | KJ490597 | KJ490418 | KJ490476 | KJ490539 | KJ490539 | Huang et al. [60] |
| Diaporthe bohemiae | CBS 143347 | MG281015 | MG281188 | MG281536 | MG281710 | MG281361 | Guarnaccia et al. [19] |
| Diaporthe caryae | CFCC 52563 | MH121498 | MH121580 | MH121540 | MH121422 | MH121458 | Yang et al. [13] |
| Diaporthe caryae | CFCC 52564 | MH121499 | MH121581 | MH121541 | MH121423 | MH121459 | Yang et al. [13] |
| Diaporthe cercidis | CFCC 52565 | MH121500 | MH121582 | MH121542 | MH121424 | MH121460 | Yang et al. [13] |
| Diaporthe cercidis | CFCC 52566 | MH121501 | MH121583 | MH121543 | MH121425 | MH121461 | Yang et al. [13] |
| Diaporthe chongqingensis | PSCG 435 | MK626916 | MK691321 | MK654866 | MK691209 | MK726257 | Guo et al. [15] |
| Diaporthe chongqingensis | PSCG 436 | MK626917 | MK691322 | MK654867 | MK691208 | MK726256 | Guo et al. [15] |
| Diaporthe cichorii | MFLUCC 17-1023 | KY964220 | KY964104 | KY964176 | KY964133 | N/A | Dissanayake et al. [9] |
| Diaporthe cinnamomi | CFCC 52569 | MH121504 | MH121586 | MH121546 | N/A | MH121464 | Yang et al. [13] |
| Diaporthe cinnamomi | CFCC 52570 | MH121505 | MH121587 | MH121547 | N/A | MH121465 | Yang et al. [13] |
| Diaporthe cissampeli | CPC 27302 | KX228273 | KX228384 | N/A | N/A | KX228366 | Crous et al. [61] |
| Diaporthe citri | CBS 135422 | KC843311 | KC843187 | KC843071 | KC843157 | MF418281 | Udayanga et al. [6] |
| Diaporthe citri | AR 4469 | KC843321 | KC843197 | KC843081 | KC843167 | N/A | Udayanga et al. [6] |
| Diaporthe conica | CFCC 52571 | MH121506 | MH121588 | MH121548 | MH121428 | MH121466 | Yang et al. [13] |
| Diaporthe conica | CFCC 52572 | MH121507 | MH121589 | MH121549 | MH121429 | MH121467 | Yang et al. [13] |
| Diaporthe coryli | CFCC 53083 | MK432661 | MK578061 | MK578135 | MK442981 | MK443006 | Yang et al. [14] |
| Diaporthe coryli | CFCC 53084 | MK432662 | MK578062 | MK578136 | MK442982 | MK443007 | Yang et al. [14] |
| Diaporthe discoidispora | ZJUD89 | KJ490624 | KJ490445 | KJ490503 | N/A | KJ490566 | Huang et al. [60] |
| Diaporthe discoidispora | ZJUD87 | KJ490622 | KJ490443 | KJ490501 | N/A | KJ490564 | Huang et al. [60] |
| Diaporthe eres | AR 5193 | KJ210529 | KJ420799 | KJ210550 | KJ434999 | KJ420850 | Udayanga et al. [7] |
| Diaporthe eres | CBS 138598 | KJ210521 | KJ420787 | KJ210545 | KJ435027 | KJ420837 | Udayanga et al. [7] |
| Diaporthe foikelawen | CBS 145289 | MN509714 | MN509725 | MN509736 | N/A | N/A | Zapata et al. [45] |
| Diaporthe foikelawen | CBS 145287 | MN509713 | MN509724 | MN509735 | N/A | N/A | Zapata et al. [45] |
| Diaporthe fulvicolor | PSCG 051 | MK626859 | MK691236 | MK654806 | MK691132 | MK726163 | Guo et al. [15] |
| Diaporthe fulvicolor | PSCG 057 | MK626858 | MK691233 | MK654810 | MK691131 | MK726164 | Guo et al. [15] |
| Diaporthe gulyae | BRIP 53158 | JF431284 | KJ197271 | N645799 | N/A | N/A | Thompson et al. [62] |
| Diaporthe gulyae | BRIP 54025 | JF431299 | KJ197272 | JN645803 | N/A | N/A | Thompson et al. [62] |
| Diaporthe helicis | AR 5211 | KJ210538 | KJ420828 | KJ210559 | KJ435043 | KJ420875 | Udayanga et al. [7] |
| Diaporthe hungariae | CBS 143353 | MG281126 | MG281299 | MG281647 | MG281823 | MG281474 | Guarnaccia et al. [19] |
| Diaporthe hungariae | CBS 143354 | MG281127 | MG281300 | MG281648 | MG281824 | MG281475 | Guarnaccia et al. [19] |
| Diaporthe juglandicola | CFCC 51134 | KU985101 | KX024634 | KX024628 | KX024616 | KX024622 | Yang et al. [18] |
| Diaporthe mahothocarpus | CGMCC 3.15181 | KC153096 | KF576312 | KC153087 | N/A | N/A | Gao et al. [63] |
| Diaporthe mahothocarpus | CGMCC 3.15182 | KC153097 | N/A | KC153088 | N/A | N/A | Gao et al. [63] |
| Diaporthe malorum | CAA734 | KY435638 | KY435668 | KY435627 | KY435658 | KY435648 | Santos et al. [16] |
| Diaporthe millettia | GUCC 9167 | MK398674 | MK502089 | MK480609 | MK502086 | N/A | Long et al. [26] |
| Diaporthe nobilis | CBS 587.79 | KC343153 | KC344121 | KC343879 | KC343395 | KC343637 | Li et al. [25] |
| Diaporthe novem | CBS 127270 | KC343155 | KC344123 | KC343881 | KC343397 | KC343640 | Santos et al. [64] |
| Diaporthe novem | CBS 127271 | KC343157 | KC344125 | KC343883 | KC343399 | KC343641 | Santos et al. [64] |
| Diaporthe oraccinii | LC 3166 | KP267863 | KP293443 | KP267937 | N/A | KP293517 | Gao et al. [63] |
| Diaporthe osmanthusis | GUCC 9165 | MK398675 | MK502090 | MK480610 | MK502087 | N/A | Long et al. [26] |
| Diaporthe paranensis | CBS 133184 | KC343171 | KC344139 | KC343897 | KC343413 | KC343655 | Gomes et al. [4] |
| Diaporthe parvae | PSCG 034 | MK626919 | MK691248 | MK654858 | N/A | MK726210 | Guo et al. [15] |
| Diaporthe parvae | PSCG 035 | MK626920 | MK691249 | MK654859 | MK691169 | MK726211 | Guo et al. [15] |
| Diaporthe pascoei | BRIP 54847 | JX862532 | KF170924 | JX862538 | N/A | N/A | Tan et al. [65] |
| Diaporthe passiflorae | CPC 19183 | JX069860 | KY435674 | KY435633 | KY435664 | KY435654 | Crous et al. [66] |
| Diaporthe patagonica | CBS 145291 | MN509717 | MN509728 | MN509739 | N/A | N/A | Zapata et al. [45] |
| Diaporthe patagonica | CBS 145755 | MN509718 | MN509729 | MN509740 | N/A | N/A | Zapata et al. [45] |
| Diaporthe perjuncta | CBS 109745 | KC343172 | KC344140 | KC343898 | KC343414 | KC343656 | van Niekerk et al. [67] |
| Diaporthe phragmitis | CBS 138897 | KP004445 | KP004507 | N/A | N/A | KP004503 | Crous et al. [55] |
| Diaporthe psoraleae | CBS 136412 | KF777158 | KF777251 | KF777245 | N/A | N/A | Crous et al. [68] |
| Diaporthe psoraleae-pinnatae | CBS 136413 | KF777159 | KF777252 | N/A | N/A | N/A | Crous et al. [68] |
| Diaporthe pterocarpicola | MFLUCC 10-0580a | JQ619887 | JX275441 | JX275403 | JX197433 | N/A | Udayanga et al. [69] |
| Diaporthe pterocarpicola | MFLUCC 10-0580b | JQ619887 | JX275441 | JX275403 | JX197433 | N/A | Udayanga et al. [69] |
| Diaporthe pterocarpi | MFLUCC 10-0571 | JX197433 | JX275460 | JX275416 | JX197451 | N/A | Udayanga et al. [69] |
| Diaporthe pterocarpi | MFLUCC 10-0575 | JQ619901 | JX275462 | JX275418 | JX197453 | N/A | Udayanga et al. [69] |
| Diaporthe rostrata | CFCC 50062 | KP208847 | KP208855 | KP208853 | KP208849 | KP208851 | Fan et al. [17] |
| Diaporthe rostrata | CFCC 50063 | KP208848 | KP208856 | KP208854 | KP208850 | KP208852 | Fan et al. [17] |
| Diaporthe rudis | AR 3422 | KC843331 | KC843177 | KC843090 | KC843146 | N/A | Udayanga et al. [6] |
| Diaporthe rudis | AR 3654 | KC843338 | KC843184 | KC843097 | KC843153 | N/A | Udayanga et al. [6] |
| Diaporthe sackstonii | BRIP 54669b | KJ197287 | KJ197267 | KJ197249 | N/A | N/A | Thompson et al. [70] |
| Diaporthe sennae | CFCC 51636 | KY203724 | KY228891 | KY228885 | KY228875 | N/A | Yang et al. [18] |
| Diaporthe sennae | CFCC 51637 | KY203725 | KY228892 | KY228886 | KY228876 | N/A | Yang et al. [18] |
| Diaporthe sennicola | CFCC 51634 | KY203722 | KY228889 | KY228883 | KY228873 | KY228879 | Yang et al. [18] |
| Diaporthe sennicola | CFCC 51635 | KY203723 | KY228890 | KY228884 | KY228874 | KY228880 | Yang et al. [18] |
| Diaporthe shaanxiensis | CFCC 53106 | MK432654 | N/A | MK578130 | MK442976 | MK443001 | Yang et al. [14] |
| Diaporthe shaanxiensis | CFCC 53107 | MK432655 | N/A | MK578131 | MK442977 | MK443002 | Yang et al. [14] |
| Diaporthe sojae | BRIP 54033 | JF431295 | N/A | KC343901 | N/A | N/A | Udayanga et al. [71] |
| Diaporthe sojae | CBS 116019 | KC343175 | KC344143 | KC343901 | KC343417 | KC343659 | Udayanga et al. [71] |
| Diaporthe sojae | FAU 455 | KJ590712 | KJ610868 | KJ590755 | KJ612109 | KJ659201 | Udayanga et al. [71] |
| Diaporthe sojae | FAU 635 | KJ590719 | KJ610875 | KJ590762 | KJ612116 | KJ659208 | Udayanga et al. [71] |
| Diaporthe spartinicola | CPC 24951 | KR611879 | KR857695 | N/A | N/A | KR857696 | Crous et al. [71] |
| Diaporthe spinosa | PSCG 383 | MK626849 | MK691234 | MK654811 | MK691129 | MK726156 | Guo et al. [15] |
| Diaporthe spinosa | PSCG 279 | MK626925 | MK691235 | MK654801 | MK691126 | MK726155 | Guo et al. [15] |
| Diaporthe subordinaria | CBS 464.90 | KC343214 | KC344182 | KC343940 | KC343456 | KC343698 | Gomes et al. [4] |
| Diaporthe subordinaria | CBS 101711 | KC343213 | KC344182 | KC343939 | KC343455 | KC343697 | Gomes et al. [4] |
| Diaporthe taoicola | MFLUCC 16-0117 | KU557567 | KU557591 | KU557635 | N/A | N/A | Dissanayake et al. [9] |
| Diaporthe torilicola | MFLUCC 17-1051 | KY964212 | KY964096 | KY964168 | KY964127 | N/A | Dissanayake et al. [9] |
| Diaporthe toxica | CBS 534.93 | KC343220 | KC344188 | KC343946 | KC343462 | KC343704 | Williamson et al. [72] |
| Diaporthe toxica | CBS 546.93 | KC343222 | KC344190 | KC343948 | KC343464 | KC343706 | Williamson et al. [72] |
| Diaporthe vangueriae | CPC 22703 | KJ869137 | KJ869247 | N/A | N/A | N/A | Crous et al. [55] |
| Diaporthe vawdreyi | BRIP 57887a | KR936126 | KR936128 | KR936129 | N/A | N/A | Crous et al. [73] |
| Diaporthe zaobaisu | PSCG 031 | MK626922 | MK691245 | MK654855 | N/A | MK726207 | Guo et al. [15] |
| Diaporthe zaobaisu | PSCG 032 | MK626923 | MK691246 | MK654856 | N/A | MK726208 | Guo et al. [15] |
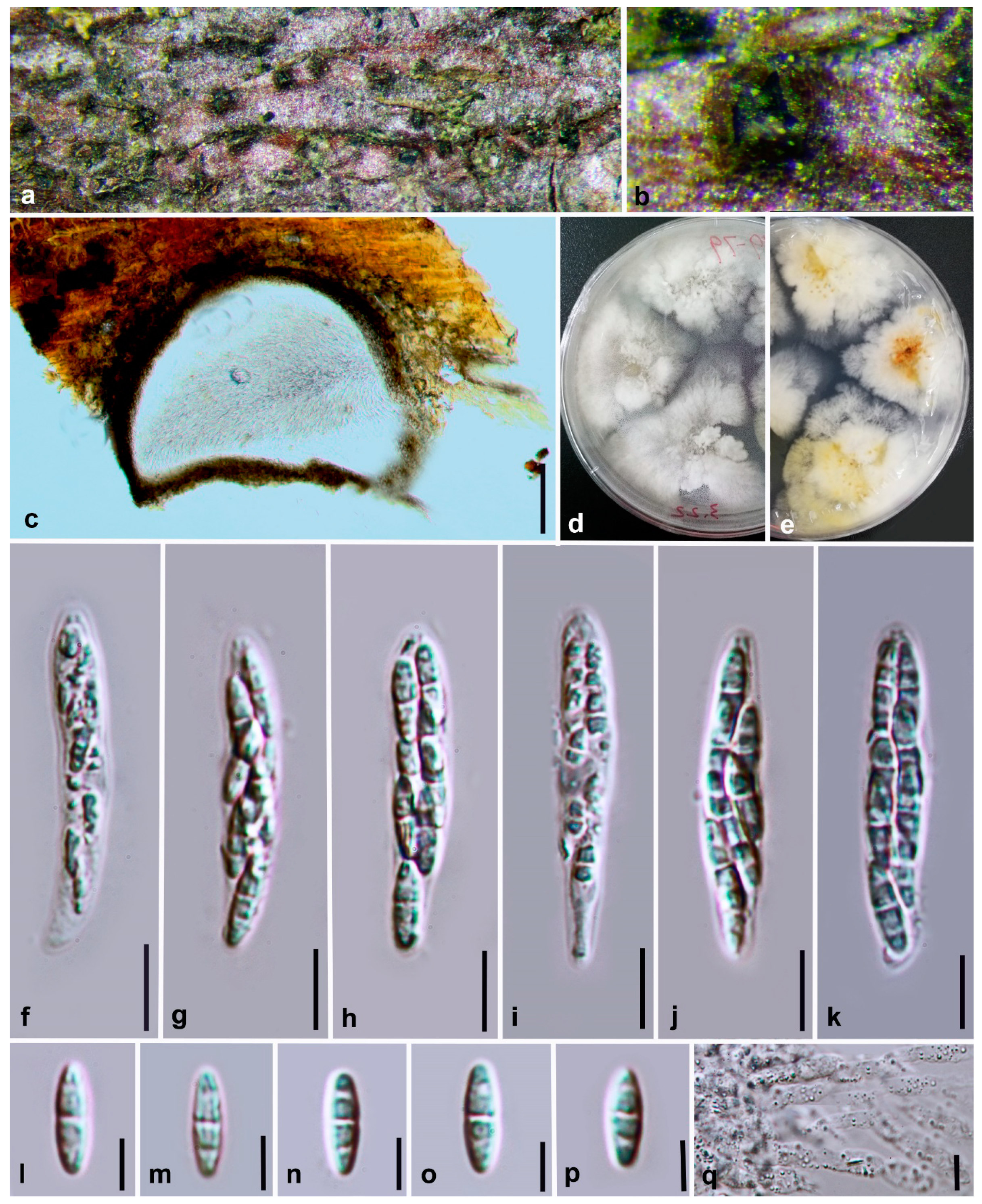
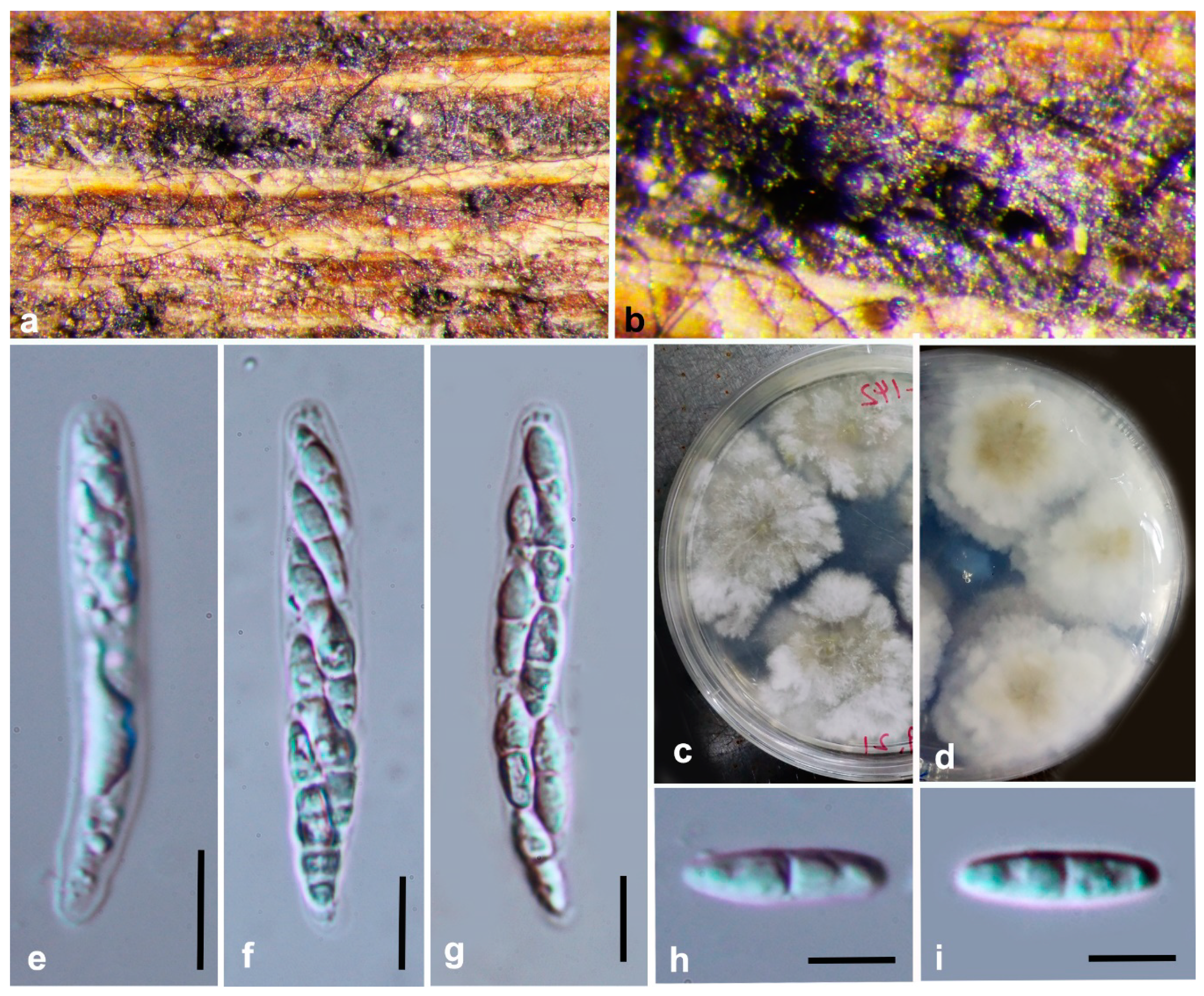
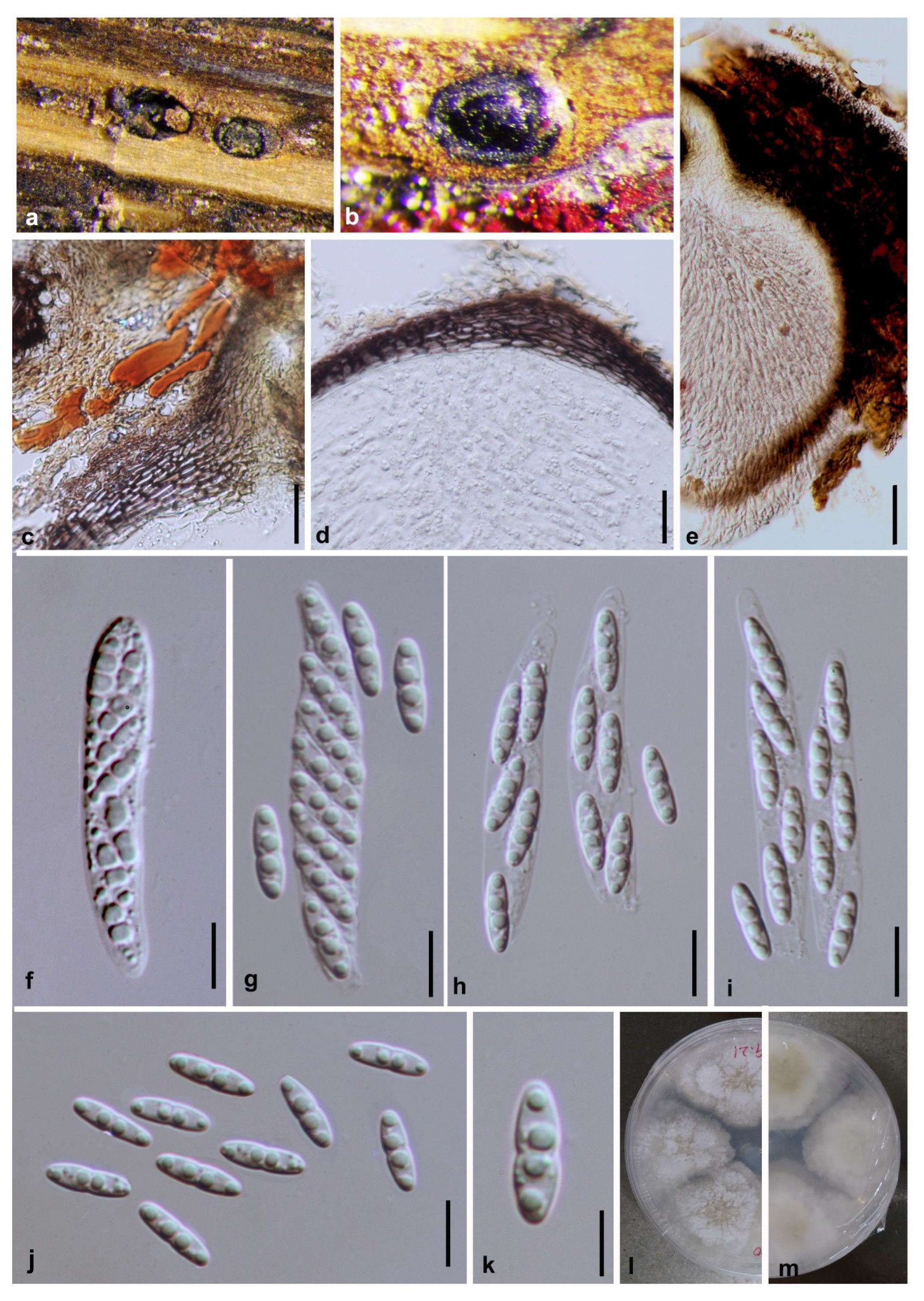
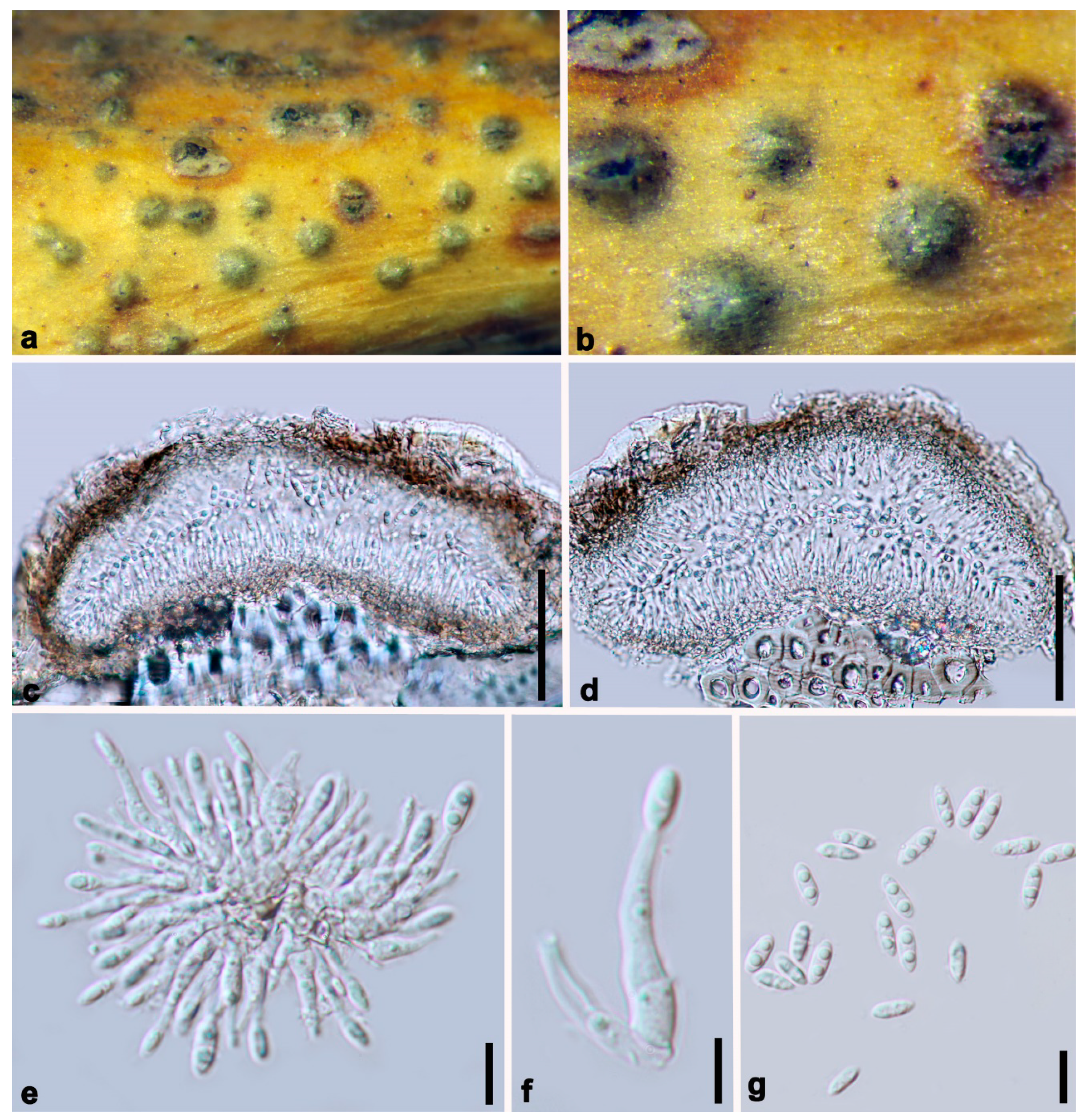
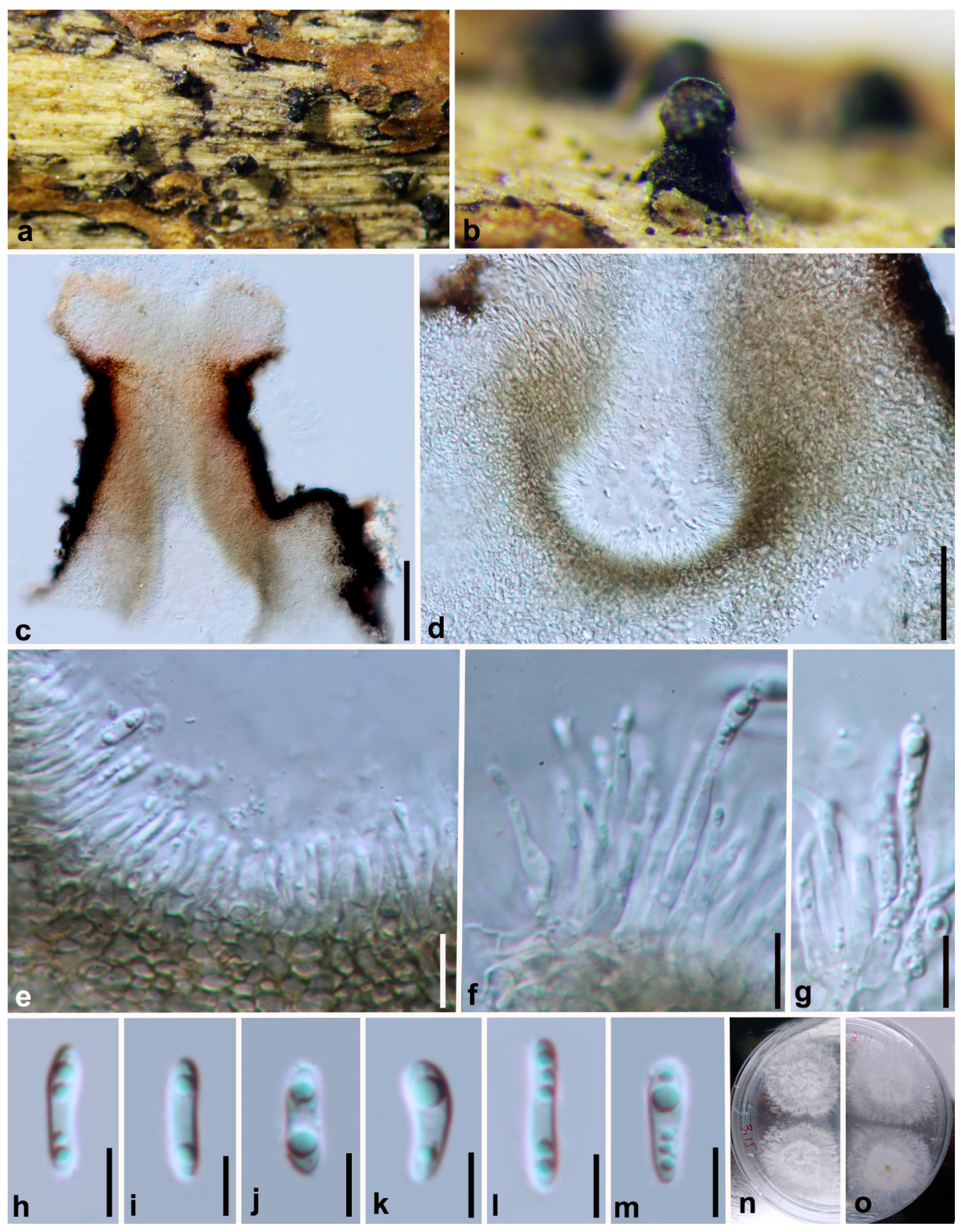
3.2. Morphology and Culture Characteristics
Taxonomy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hyde, K.D.; Nilsson, R.H.; Alias, S.A.; Ariyawansa, H.A.; Blair, J.E. One stop shop, backbone trees for important phytopathogenic genera, I. Fungal Divers. 2014, 67, 21–125. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.C.; Huang, S.K. Towards a natural classification and backbone tree for Sordariomycetes. Fungal Divers. 2015, 72, 199–301. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.C.; Bhat, D.J. Families of Sordariomycetes. Fungal Divers. 2016, 79, 1–317. [Google Scholar] [CrossRef]
- Gomes, R.R.; Glienke, C.; Videira, S.I.R.; Lombard, L.; Groenewald, J.Z. Diaporthe; a genus of endophytic; saprobic and plant pathogenic fungi. Persoonia 2013, 31, 1–41. [Google Scholar] [CrossRef]
- Rossman, A.Y.; Adams, G.C.; Cannon, P.F.; Castlebury, L.A.; Crous, P.W.; Gryzenhout, M.; Jaklitsch, W.M.; Mejia, L.C.; Stoykov, D.; Udayanga, D.; et al. Recommendations of generic names in Diaporthales competing for protection or use. IMA Fungus 2015, 6, 145–154. [Google Scholar] [CrossRef]
- Udayanga, D.; Castlebury, L.A.; Rossman, A.Y.; Hyde, K.D. Species limits in Diaporthe; molecular re-assessment of D. citri; D. cytosporella; D. foeniculina and D. rudis. Persoonia 2014, 32, 83–101. [Google Scholar] [CrossRef] [PubMed]
- Udayanga, D.; Castlebury, L.A.; Rossman, A.Y.; Chukeatirote, E.; Hyde, K.D. Insights into the genus Diaporthe; phylogenetic species delimitation in the D. eres species complex. Fungal Divers. 2014, 67, 203–229. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Liu, M.; Zhang, W.; Chen, Z.; Udayanga, D. Morphological and molecular characterization of Diaporthe species associated with grapevine trunk disease in China. Fungal Biol. 2015, 11, 283–294. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Zhang, W.; Liu, M.; Hyde, K.D.; Zhao, W.S. Diaporthe species associated with peach tree dieback in Hubei, China. Mycosphere 2017, 8, 533–549. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Phillips, A.J.L.; Yan, J.Y.; Li, X.H.; Hyde, K.D. The current status of species in Diaporthe. Mycosphere 2017, 8, 1106–1156. [Google Scholar] [CrossRef]
- van Rensburg, J.C.J.; Lamprecht, S.C.; Groenewald, J.Z.; Castlebury, L.A.; Crous, P.W. Characterization of Phomopsis spp. associated with dieback of rooibos (Aspalathus linearis) in South Africa. Stud. Mycol. 2006, 55, 65–74. [Google Scholar] [CrossRef]
- Udayanga, D.; Liu, X.Z.; McKenzie, E.H.C.; Chukeatirote, E.; Bahkali, A.H. The genus Phomopsis: Biology, applications, species concepts and names of common pathogens. Fungal Divers. 2011, 50, 189–225. [Google Scholar] [CrossRef]
- Yang, Q.; Du, Z.; Tian, C.M. Phylogeny and morphology reveal two new species of Diaporthe from Traditional Chinese Medicine in Northeast China. Phytotaxa 2018, 336, 159–170. [Google Scholar] [CrossRef]
- Yang, Q.; Jiang, N.; Tian, C.M. Three new Diaporthe species from Shaanxi Province; China. MycoKeys 2020, 67, 1–18. [Google Scholar] [CrossRef]
- Guo, Y.S.; Crous, P.W.; Bai, Q.; Fu, M.; Yang, M.M.; Wang, X.H.; Du, Y.M.; Hong, N.; Xu, W.X.; Wang, G.P. High diversity of Diaporthe species associated with pear shoot canker in China. Persoonia 2020, 45, 132–162. [Google Scholar] [CrossRef]
- Santos, L.; Phillips, A.J.L.; Crous, P.W.; Alves, A. Diaporthe species on Rosaceae with descriptions of D. pyracanthae sp. nov. and D. malorum sp. nov. Mycosphere 2017, 8, 485–511. [Google Scholar] [CrossRef]
- Fan, X.L.; Hyde, K.D.; Udayanga, D.; Wu, X.Y. Diaporthe rostrata; a novel ascomycete from Juglans mandshurica associated with walnut dieback. Mycol. Prog. 2016, 14, 82. [Google Scholar] [CrossRef]
- Yang, Q.; Fan, X.L.; Du, Z.; Tian, C.M. Diaporthe juglandicola sp. nov. (Diaporthales; Ascomycetes) evidenced by morphological characters and phylogenetic analysis. Mycosphere 2017, 8, 817–826. [Google Scholar] [CrossRef]
- Guarnaccia, V.; Groenewald, J.Z.; Woodha, J. Diaporthe diversity and pathogenicity revealed from a broad survey of grapevine diseases in Europe. Persoonia 2018, 40, 135–153. [Google Scholar] [CrossRef]
- Wang, C.; Wen, Y. Study on Nature Reserve Management Strategy Based on Externality of Public Goods. Reform Strategy 2011, 3. [Google Scholar]
- Xu, Z.L. Dashahe Nature Reserve Assessment and Strategies for Protection and Management. For. Inventory Plan. 2003, 1. [Google Scholar]
- Yao, Z.M.; Yu, D.L.; Ran, J.C. Study on the Ecotourism and Community Economic Development of Maolan National Natural Reserve in Guizhou Province. J. Anhui Agric. Sci. 2011, 3, 87–91. [Google Scholar]
- Teng, S.C. Fungi of China; Science Press: Beijing, China, 1963; pp. 1–808. (In Chinese) [Google Scholar]
- Tai, F.L. Sylloge Fungorum Sinicorum; Science Press: Beijing, China, 1979. [Google Scholar]
- Li, Y.; Tan, P.; Zhao, D.G. Diaporthe nobilis; a new record on Camellia sinensis in Guizhou Province; China. Mycosphere 2017, 8, 1–8. [Google Scholar] [CrossRef]
- Long, H.; Zhang, Q.; Hao, Y.Y.; Shao, X.Q.; Wei, X.X.; Hyde, K.D.; Wang, Y.; Zhao, D.G. Diaporthe species in south-western China. Mycokeys 2019, 57, 113–127. [Google Scholar] [CrossRef]
- Liu, J.K.; Chomnunti, P.; Cai, L.; Phookamsak, R.; Chukeatirote, E.; Jones, E.B.G.; Moslem, M.; Hyde, K.D. Phylogeny and morphology of Neodeightonia palmicola sp. nov. from palms. Sydowia 2010, 62, 261–276. [Google Scholar]
- Chomnunti, P.; Hongsanan, S.; Aguirre-Hudson, B.; Tian, Q.; Peršoh, D.; Dhami, M.K.; Alias, A.S.; Xu, J.C.; Hyde, K.D. The sooty moulds. Fungal Divers. 2014, 66, 1–36. [Google Scholar] [CrossRef]
- Rayner, R.W. A Mycological Colour Chart; Commonwealth Mycological Institute: Kew, UK, 1970. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplifcation and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. A Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous Ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- Crous, P.W.; Gams, W.; Stalpers, J.A. MycoBank: An online initiative to launch mycology into the 21st century. Stud. Mycol. 2004, 50, 19–22. [Google Scholar]
- Hall, T. Bioedit. Department of Microbiology. North Carolina State University. Available online: http://wwwmbioncsuedu/BioEdit/Bioedithtml (accessed on 29 March 2020).
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Hardy, G.E.J.; Gené, J. Fungal Planet description sheets: 716–784. Persoonia 2018, 40, 240–393. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Luangsa-ard, J.J.; Wingfield, M.J.; Carnegie, A.J.; Hernández-Restrepo, M. Fungal Planet description sheets: 785–867. Persoonia 2018, 41, 238–417. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Carnegie, A.J.; Wingfield, M.J.; Sharma, R.; Mughini, G. Fungal Planet description sheets: 868–950. Persoonia 2019, 42, 291–473. [Google Scholar] [CrossRef] [PubMed]
- Guarnaccia, V.; Crous, P.W. Emerging citrus diseases in Europe caused by species of Diaporthe. IMA Fungus 2018, 8, 317–334. [Google Scholar] [CrossRef] [PubMed]
- Milagres, C.A.; Belisário, R.; Silva, M.A.; Lisboa, D.O.; Pinho, D.B.; Furtado, G.Q. A novel species of Diaporthe causing leaf spot in Pachira glabra. Trop. Plant. Pathol. 2019, 43, 460–467. [Google Scholar] [CrossRef]
- Ozawa, K.; Mochizuki, K.; Takagi, D.; Ishida, K.; Sunada, A.; Ohkusu, K.; Kamei, K.; Hashimoto, A.; Tanaka, K. Identification and antifungal sensitivity of two new species of Diaporthe isolated. J. Infect. Chemother. 2018, 25, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Wanasinghe, D.N.; Phukhamsakda, C.; Hyde, K.D.; Jeewon, R.; Lee, H.B. Fungal diversity notes 709–839, taxonomic and phylogenetic contributions to fungal taxa with an emphasis on fungi on Rosaceae. Fungal Divers. 2018, 89, 1–236. [Google Scholar] [CrossRef]
- Yang, Q.; Fan, X.L.; Guarnaccia, V.; Tian, C.M. High diversity of Diaporthe species associated with dieback diseases in China; with twelve new species described. Mycokeys 2018, 39, 97–149. [Google Scholar] [CrossRef]
- Hyde, K.D.; Tennakoon, D.S.; Jeewon, R.; Bhat, D.J.; Maharachchikumbura, S.S.N. Fungal diversity notes 1036–1150, taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2019, 96, 1–242. [Google Scholar] [CrossRef]
- Zhou, H.; Hou, C.L. Three new species of Diaporthe from China based on morphological characters and DNA sequence data analyses. Phytotaxa 2019, 422, 157–174. [Google Scholar] [CrossRef]
- Zapata, M.; Palma, M.A.; Aninat, M.J.; Piontelli, E. Polyphasic studies of new species of Diaporthe from native forest in Chile; with descriptions of Diaporthe araucanorum sp. nov.; Diaporthe foikelawen sp. nov. and Diaporthe patagonica sp. nov. Int. J. Syst. Evol. Microbiol. 2020, 70, 3379–3390. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Toh, H. Recent evelopments in the MAFFT multiple sequence alignment program. Brief. Bioinform. 2010, 9, 286–298. [Google Scholar] [CrossRef] [PubMed]
- Swofford, D.L. PAUP*, Phylogenetic Analysis Using Parsimony, * and Other Methods; Version 4.0b10; Sinauer Associates: Sunderland, UK, 2003. [Google Scholar]
- Stamatakis, A. RAxML-VI-HPC, maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3, Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author Evolutionary Biology Centre; Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Silvestro, D.; Michalak, I. RaxmlGUI, a Graphical Front-End for RAxML. Available online: http://sourceforgenet/projects/raxmlgui/ (accessed on 29 March 2020).
- Kishino, H.; Hasegawa, M. Evaluation of the maximum likelihood estimate of the evolutionary tree topologies from DNA sequence data; and the branching order in Hominoidea. J. Mol. Evol. 1989, 29, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.A.; Bhat, J.; Buyck, B.; Cai, L.; Dai, Y.C.; Abd-Elsalam, K.A.; Ertz, D.; Hidayat, I.; et al. The Faces of Fungi database, fungal names linked with morphology; phylogeny and human impacts. Fungal Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- Vasilyeva, L.N.; Rossman, A.Y.; Farr, D.F. New species of the Diaporthales from eastern Asia and eastern North America. Mycologia 2007, 99, 916–923. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Wingfield, M.J.; Schumacher, R.K. Fungal Planet description sheets: 281–319. Persoonia 2014, 33, 212–292. [Google Scholar] [CrossRef]
- Mostert, L.; Crous, P.W.; Kang, J.C. Species of Phomopsis and a Libertella sp. occurring on grapevines with specific reference to South Africa; morphological; cultural; molecular and pathological characterization. Mycologia 2001, 93, 146–167. [Google Scholar] [CrossRef]
- Castlebury, L.A.; Farr, D.F.; Rossman, A.Y.; Jaklitsch, W.J. Diaporthe angelicae comb. nov.; a modern description and placement of Diaporthopsis in Diaporthe. Mycoscience 2003, 44, 203–208. [Google Scholar] [CrossRef]
- Hu, D.M.; Cai, L.; Hyde, K.D. Three new ascomycetes from freshwater in China. Mycologia 2012, 104, 1478–1489. [Google Scholar] [CrossRef]
- Lombard, L.; van Leeuwen, G.C.M.; Guarnaccia, V. Diaporthe species associated with Vaccinium; with specific reference to Europe. Phytopathol. Mediterr. 2014, 53, 287–299. [Google Scholar]
- Huang, F.; Udayanga, D.; Wang, X.; Hou, X.; Mei, X. Endophytic Diaporthe associated with Citrus, A phylogenetic reassessment with seven new species from China. Fungal Biol. 2015, 119, 331–347. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Richardson, D.M. Fungal Planet description sheets: 400–468. Persoonia 2016, 36, 316–458. [Google Scholar] [CrossRef]
- Thompson, S.M.; Tan, Y.P.; Young, A.J.; Neate, S.M.; Aitken, E.A. Stem cankers on sunflower (Helianthus annuus) in Australia reveal a complex of pathogenic Diaporthe (Phomopsis) species. Persoonia 2011, 27, 80–89. [Google Scholar] [CrossRef]
- Gao, Y.H.; Liu, F.; Cai, L. Unravelling Diaporthe species associated with Camellia. Syst. Biodivers. 2016, 14, 102–117. [Google Scholar] [CrossRef]
- Santos, J.M.; Vrandecic, K.; Cosic, J.; Duvnjak, T.; Phillips, A.J.L. Resolving the Diaporthe species occurring on soybean in Croatia. Persoonia 2011, 27, 9–19. [Google Scholar] [CrossRef]
- Tan, Y.P.; Edwards, J.; Grice, K.R.E.; Shivas, R.G. Molecular phylogenetic analysis reveals six new Diaporthe species from Australia. Fungal Divers. 2013, 61, 251–260. [Google Scholar] [CrossRef]
- Crous, P.W.; Summerell, B.W.; Shivas, R.G.; Burgess, T.I. Fungal Planet description sheets: 107–127. Persoonia 2012, 28, 138–182. [Google Scholar] [CrossRef]
- van Niekerk, J.M.; Groenewald, J.Z.; Farr, D.F.; Fourie, P.H.; Halleen, F.; Crous, P.W. Reassessment of Phomopsis species on grapevines. Australas. Plant. Pathol. 2005, 34, 27–39. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Guarro, J. Fungal Planet description sheets: 154–213. Persoonia 2013, 31, 188–296. [Google Scholar] [CrossRef]
- Udayanga, D.; Liu, X.Z.; MCkenzie, E.H.C.; Chukeatirote, E.; Hyde, K.D. Multi-locus phylogeny reveals three new species of Diaporthe from Thailand. Cryptogam. Mycol. 2012, 33, 295–309. [Google Scholar] [CrossRef]
- Thompson, S.M.; Tan, Y.P.; Shivas, R.G.; Neate, S.M. Green and brown bridges between weeds and crops reveal novel Diaporthe species in Australia. Persoonia 2015, 35, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Udayanga, D.; Castlebury, L.A.; Rossman, A.Y. The Diaporthe sojae species complex; phylogenetic re-assessment of pathogens associated with soybean; cucurbits and other field crops. Fungal Biol. 2015, 119, 383–407. [Google Scholar] [CrossRef] [PubMed]
- Williamson, P.M.; Higher, A.S.; Gams, W. Diaporthe toxica sp. nov. the cause of lupinosis in sheep. Mycol. Res. 1994, 98, 1364–1368. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Roux, J.J.; Richardson, D.M. Fungal Planet description sheets; 371–399. Persoonia 2015, 35, 264–327. [Google Scholar] [CrossRef]
- Crous, P.W.; Schumacher, R.K.; Wingfield, M.J.; Lombard, L. Fungal Systematics and Evolution; FUSE 1. Sydowia 2015, 67, 81–118. [Google Scholar]
- Udayanga, D.; Liu, X.Z.; Crous, P.W.; McKenzie, E.H.C. A multi-locus phylogenetic evaluation of Diaporthe (Phomopsis). Fungal Divers. 2012, 56, 157–171. [Google Scholar] [CrossRef]
- Santos, J.M.; Phillips, A.J.L. Resolving the complex of Diaporthe (Phomopsis) species occurring on Foeniculum vulgare in Portugal. Fungal Divers. 2009, 34, 111–125. [Google Scholar]
- Santos, J.M.; Correia, V.G.; Phillips, A.J.L. Primers for mating-type diagnosis in Diaporthe and Phomopsis, their use in teleomorph induction in vitro and biological species definition. Fungal Biol. 2010, 114, 255–270. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, F.; Duan, W.; Crous, P.W.; Cai, L. Diaporthe is paraphyletic. IMA Fungus 2017, 8, 153–187. [Google Scholar] [CrossRef]
- Nitschke, T. Pyrenomycetes Germanici 2; Eduard Trewendt: Breslau, Germany, 1870; pp. 161–320. [Google Scholar]
- Farr, D.F.; Rossman, A.Y. Fungal Databases; Systematic Mycology and Microbiology Laboratory; ARS; USDA. (SMML Database). Available online: https://nt.ars-grin.gov/fungaldatabases/ (accessed on 25 March 2020).









| Species | Isolate | Locality | ITS | tef | tub | cal | his |
|---|---|---|---|---|---|---|---|
| Diaporthe cercidis | GZCC 19-0079 | Guiyang Xiaochehe Wetland Park | MT385942 | MT424677 | MT424698 | MW022466 | MW022482 |
| D. cercidis | GZCC 19-0124 | Maolan Nature Reserve | MT385943 | MT424678 | MT424699 | MW022467 | MW022483 |
| D. cercidis | GZCC 19-0217 | Xingyi Wanfenglin | MT385944 | MT424679 | MT424700 | MW022468 | MW022484 |
| D. cinnamomi | GZCC 19-0274 | Maolan Nature Reserve | MT385945 | MT424680 | N/A | MT424717 | MW022485 |
| D. conica | GZCC 19-0242 | Maolan NatureReserve | MT385946 | MT424681 | MT424701 | MW022469 | MW022486 |
| D. constrictospora | CGMCC 3.20096 | Maolan Nature Reserve | MT385947 | MT424682 | MT424702 | MT424718 | MW022487 |
| D. constrictospora | GZCC 19-0065 | Guiyang Huaxi Wetland Park | MT385948 | MT424683 | MT424703 | MT424719 | N/A |
| D. ellipsospora | CGMCC 3.20099 | Xingyi Wanfenglin | MT385949 | MT424684 | MT424704 | MT424720 | MW022488 |
| D. ellipsospora | GZCC 19-0342 | Xingyi Wanfenglin | MT797176 | MT793019 | MT793030 | MT786247 | MW022489 |
| D. ellipsospora | GZCC 19-0357 | Maolan Nature Reserve | MT797177 | MT793020 | MT793031 | MT786248 | MW022490 |
| D. guttulata | CGMCC 3.20100 | Maolan Nature Reserve | MT385950 | MT424685 | MT424705 | MW022470 | MW022491 |
| D. guttulata | GZCC 19-0371 | Suiyang water nature reserve | MT797178 | MT793021 | MT793032 | MW022471 | MW022492 |
| D. irregularis | CGMCC 3.20092 | Suiyang water nature reserve | MT385951 | MT424686 | MT424706 | MT424721 | N/A |
| D. irregularis | GZCC 19-0344 | Suiyang water nature reserve | MT797179 | MT793022 | MT793033 | MT786249 | N/A |
| D. irregularis | GZCC 19-0362 | Suiyang water nature reserve | MT797180 | MT793023 | MT793034 | MT786250 | N/A |
| D. irregularis | GZCC 19-0352 | Suiyang water nature reserve | MT797181 | MT793024 | MT793035 | MT786251 | N/A |
| D. lenispora | CGMCC 3.20101 | Suiyang water nature reserve | MT385952 | MT424687 | MT424707 | MW022472 | MW022493 |
| D. lenispora | GZCC 19-0343 | Xingyi Wanfenglin | MT797182 | MT793025 | MT793036 | MW022473 | MW022494 |
| D. lenispora | GZCC 19-0351 | Xingyi Wanfenglin | MT797183 | MT793026 | MT793037 | MW022474 | MW022495 |
| D. minima | CGMCC 3.20097 | Guiyang Huaxi Wetland Park | MT385953 | MT424688 | MT424708 | MT424722 | MW022496 |
| D. minima | GZCC 19-0070 | Guiyang Huaxi Wetland Park | MT385954 | MT424689 | MT424709 | MT424723 | MW022497 |
| D. minima | GZCC 19-0061 | Guiyang Huaxi Wetland Park | MT385955 | MT424690 | MT424710 | MT424724 | MW022498 |
| D. minima | GZCC 19-0207 | Guiyang Huaxi Wetland Park | MT385956 | MT424691 | MT424711 | MT424725 | N/A |
| D. minusculata | CGMCC 3.20098 | Xingyi Wanfenglin | MT385957 | MT424692 | MT424712 | MW022475 | MW022499 |
| D. minusculata | GZCC 19-0345 | Suiyang water nature reserve | MT797184 | MT793027 | MT793038 | MW022476 | MW022500 |
| D. minusculata | GZCC 19-0366 | Suiyang water nature reserve | MT797185 | MT793028 | MT793039 | MW022477 | MW022501 |
| D. minusculata | GZCC 19-0372 | Xingyi Wanfenglin | MT797186 | MT793029 | MT793040 | MW022478 | MW022502 |
| D. nobilis | GZCC 19-0213 | Fanjing mountain | MT385958 | MT424693 | MT424713 | MT424726 | MW022503 |
| D. nobilis | GZCC 19-0220 | Xingyi Wanfenglin | MT385959 | MT424694 | MT424714 | MW022479 | MW022504 |
| D. nobilis | GZCC 19-0214 | Fanjing mountain | MT385960 | MT424695 | MT424715 | MW022480 | MW022505 |
| D. sackstonii | GZCC 19-0129 | Maolan Nature Reserve | MT385962 | MT424697 | MT424716 | MT424727 | MW022507 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dissanayake, A.J.; Chen, Y.-Y.; Liu, J.-K. Unravelling Diaporthe Species Associated with Woody Hosts from Karst Formations (Guizhou) in China. J. Fungi 2020, 6, 251. https://doi.org/10.3390/jof6040251
Dissanayake AJ, Chen Y-Y, Liu J-K. Unravelling Diaporthe Species Associated with Woody Hosts from Karst Formations (Guizhou) in China. Journal of Fungi. 2020; 6(4):251. https://doi.org/10.3390/jof6040251
Chicago/Turabian StyleDissanayake, Asha J., Ya-Ya Chen, and Jian-Kui (Jack) Liu. 2020. "Unravelling Diaporthe Species Associated with Woody Hosts from Karst Formations (Guizhou) in China" Journal of Fungi 6, no. 4: 251. https://doi.org/10.3390/jof6040251
APA StyleDissanayake, A. J., Chen, Y.-Y., & Liu, J.-K. (2020). Unravelling Diaporthe Species Associated with Woody Hosts from Karst Formations (Guizhou) in China. Journal of Fungi, 6(4), 251. https://doi.org/10.3390/jof6040251