Molecular Tools for Detection and Identification of Paracoccidioides Species: Current Status and Future Perspectives

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Paracoccidioidomycosis: An Overview
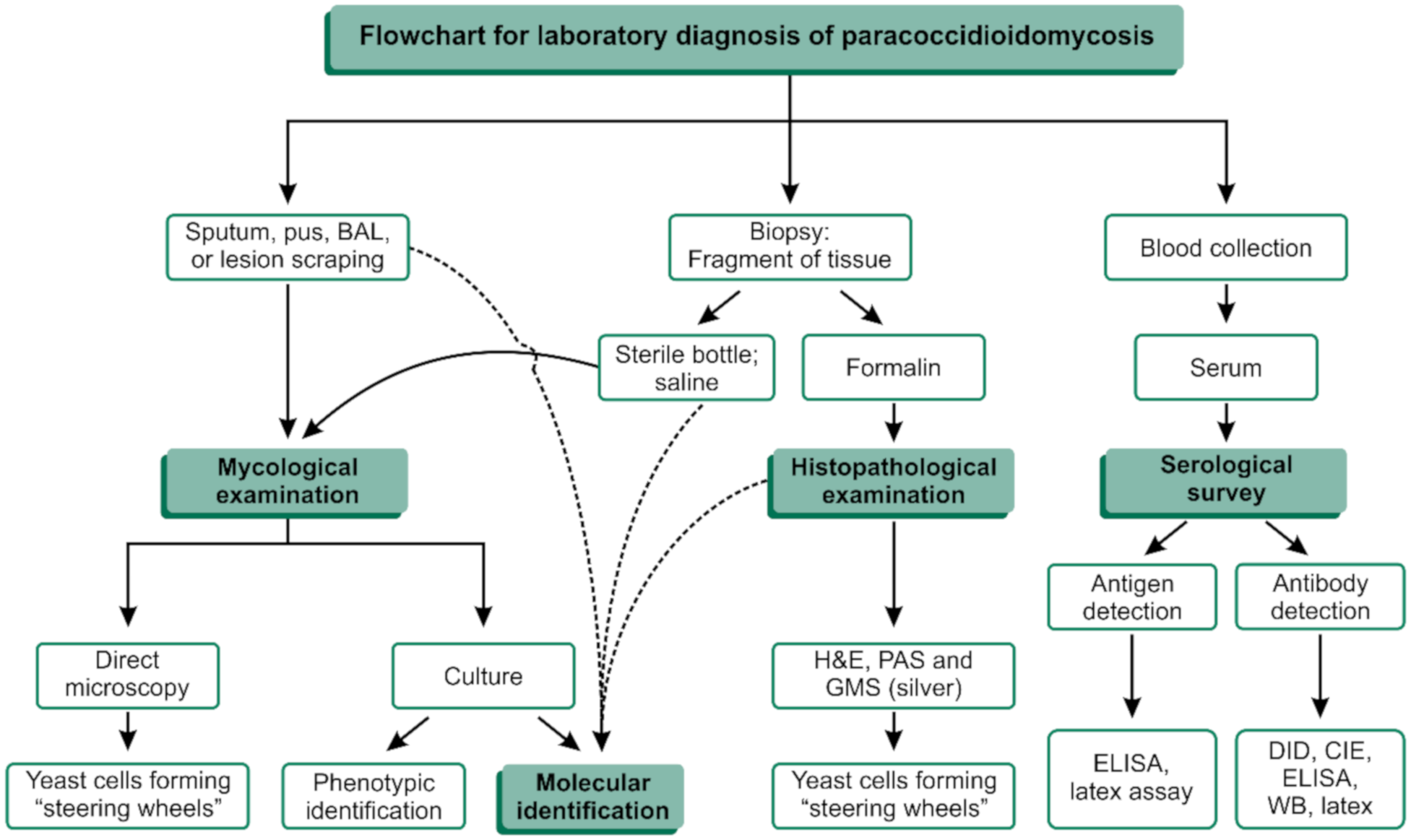
2. The Laboratory Diagnosis of PCM
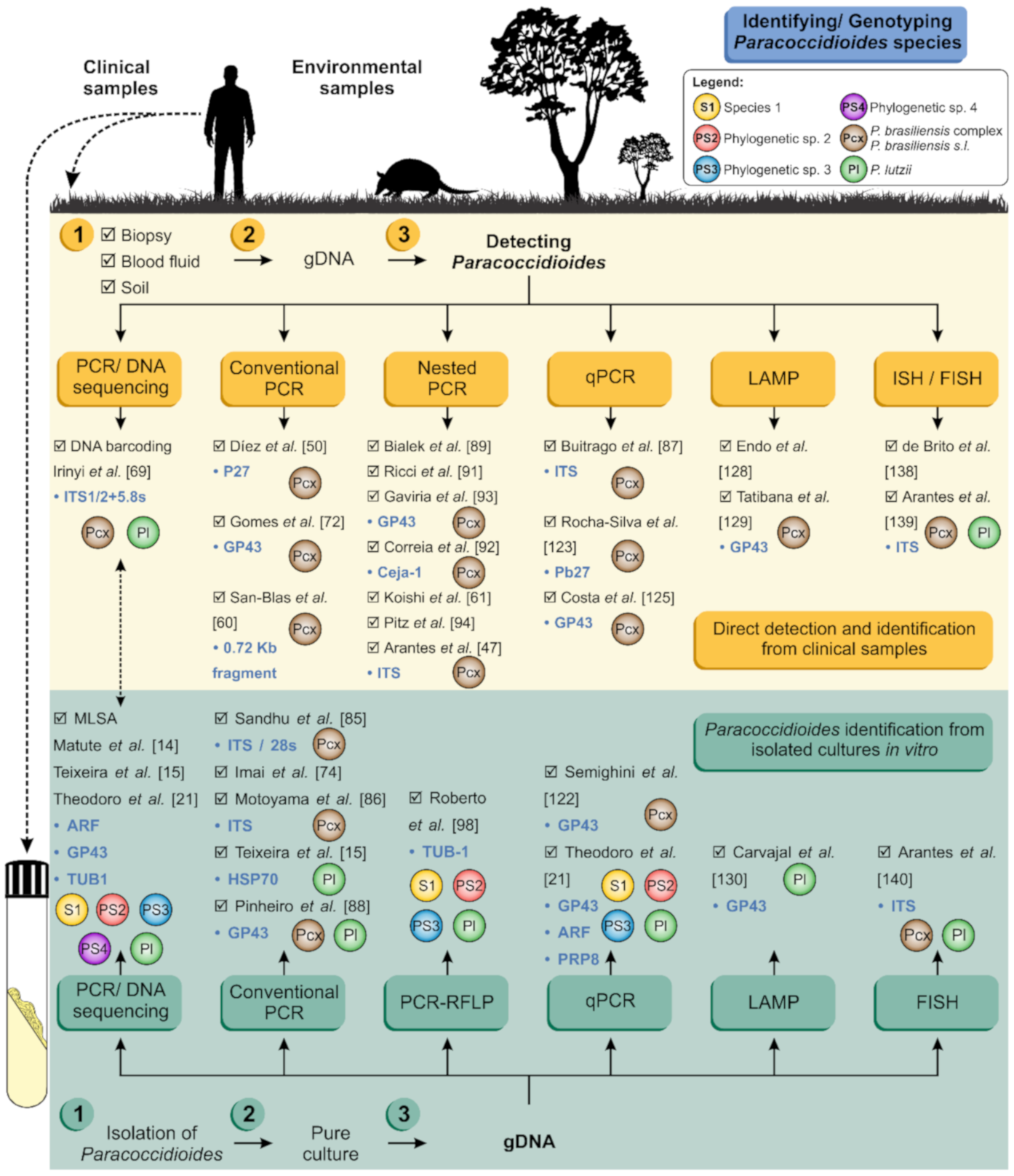
3. Molecular Diagnosis: Where Do We Stand?
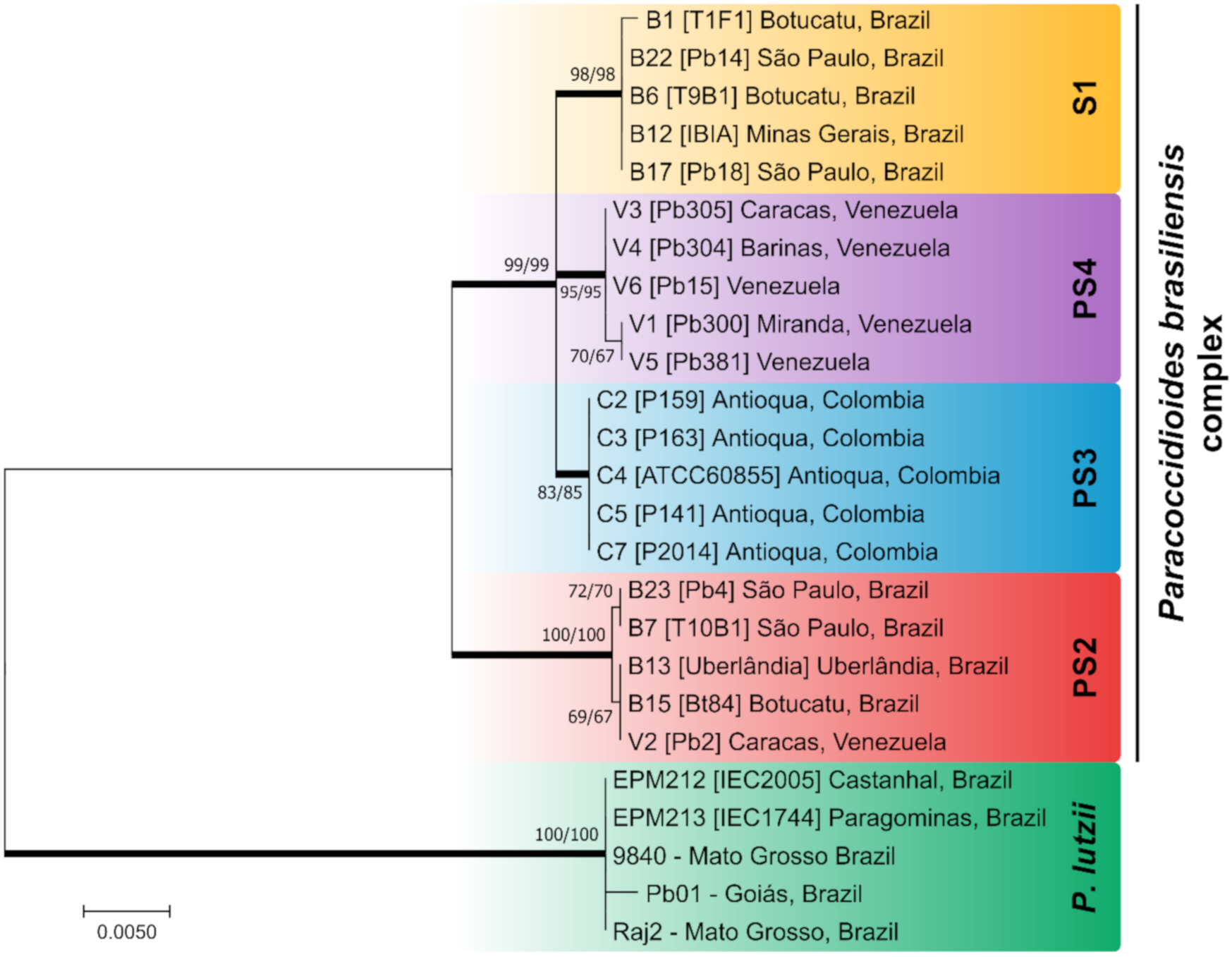
3.1. Internal Transcribed Spacer as a Barcoding Marker in Paracoccidioides
3.2. Multilocus Sequence Analysis (MLSA)
3.3. Conventional Polymerase Chain Reaction
3.4. Nested PCR and Semi-Nested PCR
3.5. Restriction Fragment Length Polymorphism and PCR-RFLP
3.6. RAPD
3.7. Microsatellite Markers
3.8. Transposable Element Amplification
3.9. Quantitative Real-Time PCR
3.10. Loop-Mediated Isothermal Amplification
3.11. Fluorescence In Situ Hybridization
3.12. Matrix-Assisted Laser Desorption Ionization Time-of-Flight Mass Spectrometry
3.13. Fourier-Transform Infrared Spectroscopy
4. Molecular Diagnosis: Where Do We Go Next?
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Shikanai-Yasuda, M.A.; Mendes, R.P.; Colombo, A.L.; Queiroz-Telles, F.; Kono, A.S.G.; Paniago, A.M.M.; Nathan, A.; Valle, A.; Bagagli, E.; Benard, G.; et al. Brazilian guidelines for the clinical management of paracoccidioidomycosis. Rev. Soc. Bras. Med. Trop. 2017, 50, 715–740. [Google Scholar] [CrossRef] [PubMed]
- Do Amaral, C.C.; Fernandes, G.F.; Rodrigues, A.M.; Burger, E.; de Camargo, Z.P. Proteomic analysis of Paracoccidioides brasiliensis complex isolates: Correlation of the levels of differentially expressed proteins with in vivo virulence. PLoS ONE 2019, 14, e0218013. [Google Scholar] [CrossRef] [PubMed]
- Ambrósio, A.; Camelo, C.; Barbosa, C.; Tomazatti, F.; Brazões, F.; Veloso, J.; Rodrigues, G.; Rodrigues, L.; Oliveira, P.; Aguiar, R.; et al. Paracoccidioidomycosis disease (Lutz-Splendore-Almeida disease): Additional workup, differential diagnosis, cure control. RMMG-Rev. Méd Minas Gerais 2014, 24, 81–92. [Google Scholar] [CrossRef]
- Restrepo, A.; Gómez, B.L.; Tobón, A. Paracoccidioidomycosis: Latin America’s own fungal disorder. Curr. Fungal Infect. Rep. 2012, 6, 303–311. [Google Scholar] [CrossRef]
- Griffiths, J.; Lopes Colombo, A.; Denning, D.W. The case for paracoccidioidomycosis to be accepted as a neglected tropical (fungal) disease. PLoS Negl. Trop. Dis. 2019, 13, e0007195. [Google Scholar] [CrossRef]
- Bellissimo-Rodrigues, F.; Machado, A.A.; Martinez, R. Paracoccidioidomycosis epidemiological features of a 1,000-cases series from a hyperendemic area on the Southeast of Brazil. Am. J. Trop. Med. Hyg. 2011, 85, 546–550. [Google Scholar] [CrossRef]
- Coutinho, Z.F.; Wanke, B.; Travassos, C.; Oliveira, R.M.; Xavier, D.R.; Coimbra, C.E., Jr. Hospital morbidity due to paracoccidioidomycosis in Brazil (1998–2006). Trop. Med. Int. Health TM IH 2015, 20, 673–680. [Google Scholar] [CrossRef]
- Martinez, R. New trends in paracoccidioidomycosis epidemiology. J. Fungi 2017, 3, 1. [Google Scholar] [CrossRef]
- Vieira, G.D.D.; Alves, T.D.C.; Lima, S.M.D.D.; Camargo, L.M.A.; Sousa, C.M.D. Paracoccidioidomycosis in a western Brazilian Amazon State: Clinical-epidemiologic profile and spatial distribution of the disease. Rev. Soc. Bras. Med. Trop. 2014, 47, 63–68. [Google Scholar] [CrossRef]
- Paniago, A.M.M.; Aguiar, J.I.A.; Aguiar, E.S.; Cunha, R.V.D.; Pereira, G.R.D.O.L.; Londero, A.T.; Wanke, B. Paracoccidioidomycosis: A clinical and epidemiological study of 422 cases observed in Mato Grosso do Sul. Rev. Soc. Bras. Med. Trop. 2003, 36, 455–459. [Google Scholar] [CrossRef]
- Garcia, N.M.; Del Negro, G.M.B.; Heins-Vaccari, E.M.; Melo, N.T.D.; Assis, C.M.D.; Lacaz, C.D.S. Paracoccidioides brasiliensis, a new strain isolated from a fecal matter of a penguin (Pygoscelis adeliae). Rev. Inst. Med. Trop. São Paulo 1993, 35, 227–235. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Moore, M. A new species of the Paracoccidioides Almeida (1930): P. cerebriformis Moore, (1935). Rev. Biol. Hig. 1935, 6, 148–162. [Google Scholar]
- Lacaz, C.D.S.; Vidal, M.S.M.; Pereira, C.N.; Heins-Vaccari, E.M.; Melo, N.T.D.; Sakai-Valente, N.; Arriagada, G.L.H. Paracoccidioides cerebriformis Moore, 1935. Mycologic and immunochemical study. Rev. Inst. Med. Trop. São Paulo 1997, 39, 141–144. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Matute, D.R.; McEwen, J.G.; Puccia, R.; Montes, B.A.; San-Blas, G.; Bagagli, E.; Rauscher, J.T.; Restrepo, A.; Morais, F.; Niño-Vega, G.; et al. Cryptic speciation and recombination in the fungus Paracoccidioides brasiliensis as revealed by gene genealogies. Mol. Biol. Evol. 2006, 23, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, M.M.; Theodoro, R.C.; de Carvalho, M.J.A.; Fernandes, L.; Paes, H.C.; Hahn, R.C.; Mendoza, L.; Bagagli, E.; San-Blas, G.; Felipe, M.S.S. Phylogenetic analysis reveals a high level of speciation in the Paracoccidioides genus. Mol. Phylogenet. Evol. 2009, 52, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Salgado-Salazar, C.; Jones, L.R.; Restrepo, Á.; McEwen, J.G. The human fungal pathogen Paracoccidioides brasiliensis (Onygenales: Ajellomycetaceae) is a complex of two species: Phylogenetic evidence from five mitochondrial markers. Cladistics 2010, 26, 613–624. [Google Scholar] [CrossRef]
- Turissini, D.A.; Gomez, O.M.; Teixeira, M.M.; McEwen, J.G.; Matute, D.R. Species boundaries in the human pathogen Paracoccidioides. Fungal Genet. Biol. 2017, 106, 9–25. [Google Scholar] [CrossRef]
- Muñoz, J.F.; Farrer, R.A.; Desjardins, C.A.; Gallo, J.E.; Sykes, S.; Sakthikumar, S.; Misas, E.; Whiston, E.A.; Bagagli, E.; Soares, C.M.A.; et al. Genome diversity, recombination, and virulence across the major lineages of Paracoccidioides. MSphere 2016, 1. [Google Scholar] [CrossRef]
- Teixeira, M.D.M.; Cattana, M.E.; Matute, D.R.; Muñoz, J.F.; Arechavala, A.; Isbell, K.; Schipper, R.; Santiso, G.; Tracogna, F.; Sosa, M.d.l.Á.; et al. Genomic diversity of the human pathogen Paracoccidioides across the South American continent. Fungal Genet. Biol. 2020, 140, 103395. [Google Scholar] [CrossRef]
- Hahn, R.C.; Rodrigues, A.M.; Fontes, C.J.; Nery, A.F.; Tadano, T.; de Padua Queiroz Junior, L.; de Camargo, Z.P. Fatal fungemia due to Paracoccidioides lutzii. Am. J. Trop. Med. Hyg. 2014, 91, 394–398. [Google Scholar] [CrossRef]
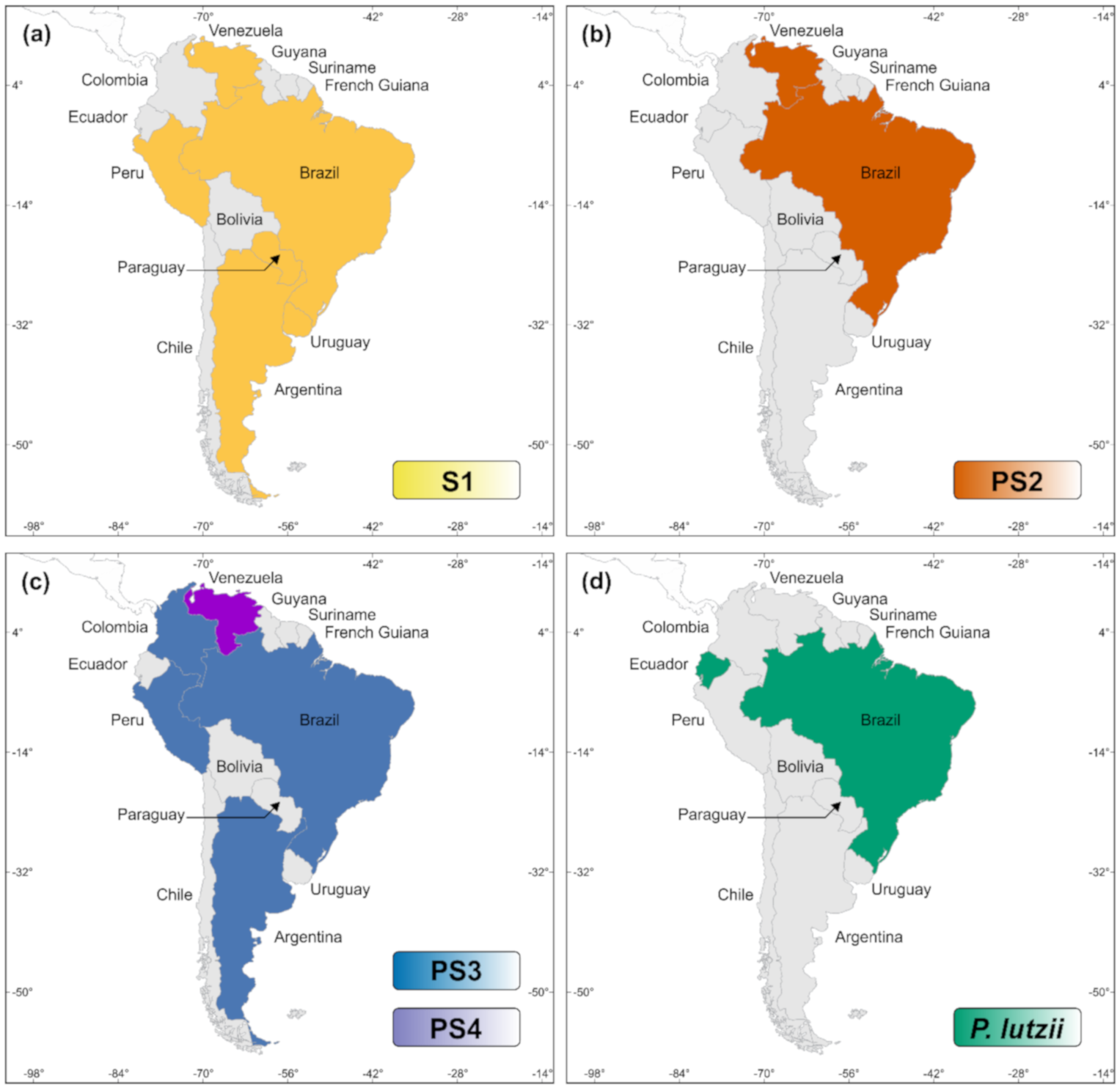
- Theodoro, R.C.; Teixeira, M.D.M.; Felipe, M.S.S.; Paduan, K.D.S.; Ribolla, P.M.; San-Blas, G.; Bagagli, E. Genus Paracoccidioides: Species recognition and biogeographic aspects. PLoS ONE 2012, 7, e37694. [Google Scholar] [CrossRef]
- Teixeira, M.M.; Theodoro, R.C.; Nino-Vega, G.; Bagagli, E.; Felipe, M.S.S. Paracoccidioides species complex: Ecology, phylogeny, sexual reproduction, and virulence. PLoS Pathog. 2014, 10, e1004397. [Google Scholar] [CrossRef]
- Cocio, T.A.; Nascimento, E.; Kress, M.R.V.Z.; Bagagli, E.; Martinez, R. Characterization of a Paracoccidioides spp. strain from southeastern Brazil genotyped as Paracoccidioides restrepiensis (PS3) and review of this phylogenetic species. Genet. Mol. Biol. 2020, 43. [Google Scholar] [CrossRef]
- Cocio, T.A.; Nascimento, E.; von Zeska Kress, M.R. Phylogenetic species of Paracoccidioides spp. isolated from clinical and environmental samples in a hyperendemic area of paracoccidioidomycosis in southeastern Brazil. J. Fungi 2020, 6, 132. [Google Scholar] [CrossRef]
- Marques-da-Silva, S.H.; Rodrigues, A.M.; de Hoog, G.S.; Silveira-Gomes, F.; de Camargo, Z.P. Occurrence of Paracoccidioides lutzii in the Amazon region: Description of two cases. Am. J. Trop. Med. Hygiene 2012, 87, 710–714. [Google Scholar] [CrossRef]
- Sarmento Tatagiba, L.; Bridi Pivatto, L.; Faccini-Martínez, Á.A.; Mendes Peçanha, P.; Grão Velloso, T.R.; Gonçalves, S.S.; Messias Rodrigues, A.; Pires Camargo, Z.; Falqueto, A. A case of paracoccidioidomycosis due to Paracoccidioides lutzii presenting sarcoid-like form. Med. Mycol. Case Rep. 2018, 19, 6–8. [Google Scholar] [CrossRef]
- Munoz, J.F.; Gallo, J.E.; Misas, E.; Priest, M.; Imamovic, A.; Young, S.; Zeng, Q.; Clay, O.K.; McEwen, J.G.; Cuomo, C.A. Genome update of the dimorphic human pathogenic fungi causing paracoccidioidomycosis. PLoS Negl. Trop Dis. 2014, 8, e3348. [Google Scholar] [CrossRef]
- Hrycyk, M.F.; Garcia Garces, H.; Bosco, S.D.M.G.; de Oliveira, S.L.; Marques, S.A.; Bagagli, E. Ecology of Paracoccidioides brasiliensis, P. lutzii and related species: Infection in armadillos, soil occurrence and mycological aspects. Med. Mycol. 2018, 56, 950–962. [Google Scholar] [CrossRef]
- Vandamme, P.; Pot, B.; Gillis, M.; de Vos, P.; Kersters, K.; Swings, J. Polyphasic taxonomy, a consensus approach to bacterial systematics. Microbiol. Rev. 1996, 60, 407–438. [Google Scholar] [CrossRef]
- Jančič, S.; Nguyen, H.D.T.; Frisvad, J.C.; Zalar, P.; Schroers, H.-J.; Seifert, K.A.; Gunde-Cimerman, N. A Taxonomic revision of the Wallemia sebi species complex. PLoS ONE 2015, 10, e0125933. [Google Scholar] [CrossRef]
- Quaedvlieg, W.; Binder, M.; Groenewald, J.Z.; Summerell, B.A.; Carnegie, A.J.; Burgess, T.I.; Crous, P.W. Introducing the consolidated species concept to resolve species in the Teratosphaeriaceae. Persoonia 2014, 33, 1–40. [Google Scholar] [CrossRef] [PubMed]
- De Hoog, G.S.; Chaturvedi, V.; Denning, D.W.; Dyer, P.S.; Frisvad, J.C.; Geiser, D.; Gräser, Y.; Guarro, J.; Haase, G.; Kwon-Chung, K.-J.; et al. Name changes in medically important fungi and their implications for clinical practice. J. Clin. Microbiol. 2015, 53, 1056–1062. [Google Scholar] [CrossRef] [PubMed]
- Hahn, R.C.; Rodrigues, A.M.; Della Terra, P.P.; Nery, A.F.; Hoffmann-Santos, H.D.; Góis, H.M.; Fontes, C.J.; de Camargo, Z.P. Clinical and epidemiological features of paracoccidioidomycosis due to Paracoccidioides lutzii. PLoS Negl. Trop Dis. 2019, 13, e0007437. [Google Scholar] [CrossRef] [PubMed]
- de Macedo, P.M.; Teixeira, M.D.M.; Barker, B.M.; Zancopé-Oliveira, R.M.; Almeida-Paes, R.; Francesconi do Valle, A.C. Clinical features and genetic background of the sympatric species Paracoccidioides brasiliensis and Paracoccidioides americana. PLoS Negl. Trop Dis. 2019, 13, e0007309. [Google Scholar] [CrossRef] [PubMed]
- Pereira, E.F.; Gegembauer, G.; Chang, M.R.; Camargo, Z.P.D.; Nunes, T.F.; Ribeiro, S.M.; Carvalho, L.R.D.; Maldonado, B.M.; Mendes, R.P.; Paniago, A.M.M. Comparison of clinico-epidemiological and radiological features in paracoccidioidomycosis patients regarding serological classification using antigens from Paracoccidioides brasiliensis complex and Paracoccidioides lutzii. PLoS Negl. Trop Dis. 2020, 14, e0008485. [Google Scholar] [CrossRef]
- Queiroz Junior Lde, P.; de Camargo, Z.P.; Tadano, T.; Rodrigues, A.M.; Takarara, D.T.; Gegembauer, G.; Araujo, L.M.; Hahn, R.C. Serological and antigenic profiles of clinical isolates of Paracoccidioides spp. from Central Western Brazil. Mycoses 2014, 57, 466–472. [Google Scholar] [CrossRef]
- Camargo, Z.P.; Rodrigues, A.M. Paracoccidioides complex. In Pocket Guide to Mycological Diagnosis; Cordeiro, R.D.A., Ed.; CRC Press: Boca Raton, FL, USA, 2019; Volume 78, pp. 125–134. [Google Scholar]
- Camargo, Z.P. Serology of paracoccidioidomycosis. Mycopathologia 2008, 165, 289–302. [Google Scholar] [CrossRef]
- de Camargo, Z.P.; Unterkircher, C.; Campoy, S.P.; Travassos, L.R. Production of Paracoccidioides brasiliensis exoantigens for immunodiffusion tests. J. Clin. Microbiol. 1988, 26, 2147–2151. [Google Scholar] [CrossRef]
- Gegembauer, G.; Araujo, L.M.; Pereira, E.F.; Rodrigues, A.M.; Paniago, A.M.; Hahn, R.C.; de Camargo, Z.P. Serology of paracoccidioidomycosis due to Paracoccidioides lutzii. PLoS Negl. Trop Dis. 2014, 8, e2986. [Google Scholar] [CrossRef]
- Bellissimo-Rodrigues, F.; Vitali, L.H.; Martinez, R. Serological diagnosis of paracoccidioidomycosis in HIV-coinfected patients. Mem. Inst. Oswaldo Cruz 2010, 105, 904–907. [Google Scholar] [CrossRef][Green Version]
- de Camargo, Z.P.; Guesdon, J.L.; Drouhet, E.; Improvisi, L. Enzyme-linked immunosorbent assay (ELISA) in the paracoccidioidomycosis. Comparison with counterimmunoelectrophoresis and erythro-immunoassay. Mycopathologia 1984, 88, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, P.O.; Rodrigues, A.M.; Fernandes, G.F.; da Silva, S.H.; Burger, E.; de Camargo, Z.P. Immunodiagnosis of paracoccidioidomycosis due to Paracoccidioides brasiliensis using a latex test: Detection of specific antibody anti-gp43 and specific antigen gp43. PLoS Negl. Trop Dis. 2015, 9, e0003516. [Google Scholar] [CrossRef] [PubMed]
- Camargo, Z.P.; Unterkircher, C.; Travassos, L.R. Identification of antigenic polypeptides of Paracoccidioides brasiliensis by immunoblotting. Med. Mycol. 1989, 27, 407–412. [Google Scholar] [CrossRef]
- Biomarkers Definitions Working Group. Biomarkers and surrogate endpoints: Preferred definitions and conceptual framework. Clin. Pharmacol. Ther. 2001, 69, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Teles, F.R.R.; Martins, M.L. Laboratorial diagnosis of paracoccidioidomycosis and new insights for the future of fungal diagnosis. Talanta 2011, 85, 2254–2264. [Google Scholar] [CrossRef] [PubMed]
- Arantes, T.D.; Theodoro, R.C.; Da Graça Macoris, S.A.; Bagagli, E. Detection of Paracoccidioides spp. in environmental aerosol samples. Med. Mycol. 2013, 51, 83–92. [Google Scholar] [CrossRef]
- Mendes, J.F.; Von Groll, A.; Poester, V.R.; Brasil, C.L.; Brandolt, T.M.; Klafke, G.B.; Nogueira, C.E.W.; Meireles, M.C.A.; Xavier, M.O. Paracoccidioides spp. in soil from the Pampa Biome in Southern Brazil. Curr. Microbiol. 2019, 76, 258–262. [Google Scholar] [CrossRef]
- Mendes, J.F.; Poester, V.R.; Groll, A.V.; Meireles, M.C.A.; Xavier, M.O. Molecular detection of Paracoccidioides in soil from an urban area of southern Brazil. Rev. Soc. Bras. Med. Trop. 2020, 53. [Google Scholar] [CrossRef]
- Díez, S.; Garcia, E.A.; Pino, P.A.; Botero, S.; Corredor, G.G.; Peralta, L.A.; Castaño, J.H.; Restrepo, A.; McEwen, J.G. PCR with Paracoccidioides brasiliensis specific primers: Potential use in ecological studies. Rev. Inst. Med. Trop Sao Paulo 1999, 41, 351–358. [Google Scholar] [CrossRef]
- Luchi, N.; Ioos, R.; Santini, A. Fast and reliable molecular methods to detect fungal pathogens in woody plants. Appl. Microbiol. Biotechnol. 2020, 104, 2453–2468. [Google Scholar] [CrossRef]
- Wickes, B.L.; Wiederhold, N.P. Molecular diagnostics in medical mycology. Nat. Commun. 2018, 9, 5135. [Google Scholar] [CrossRef]
- Teles, F.; Seixas, J. The future of novel diagnostics in medical mycology. J. Med. Microbiol. 2015, 64, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Van Stralen, K.J.; Stel, V.S.; Reitsma, J.B.; Dekker, F.W.; Zoccali, C.; Jager, K.J. Diagnostic methods I: Sensitivity, specificity, and other measures of accuracy. Kidney Int. 2009, 75, 1257–1263. [Google Scholar] [CrossRef] [PubMed]
- Halliday, C.L.; Kidd, S.E.; Sorrell, T.C.; Chen, S.C. Molecular diagnostic methods for invasive fungal disease: The horizon draws nearer? Pathology 2015, 47, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Burd, E.M. Validation of laboratory-developed molecular assays for infectious diseases. Clin. Microbiol. Rev. 2010, 23, 550–576. [Google Scholar] [CrossRef] [PubMed]
- Stone, N.R.; Gorton, R.L.; Barker, K.; Ramnarain, P.; Kibbler, C.C. Evaluation of PNA-FISH yeast traffic light for rapid identification of yeast directly from positive blood cultures and assessment of clinical impact. J. Clin. Microbiol. 2013, 51, 1301–1302. [Google Scholar] [CrossRef]
- Kiraz, N. Molecular techniques for clinical diagnostic mycology. Turk. Hij. Tecr. Biyol. Derg. 2015, 72, 263–272. [Google Scholar] [CrossRef]
- Hanson, K.E.; Caliendo, A.M.; Arias, C.A.; Englund, J.A.; Lee, M.J.; Loeb, M.; Patel, R.; El Alayli, A.; Kalot, M.A.; Falck-Ytter, Y.; et al. Infectious Diseases Society of America Guidelines on the diagnosis of Coronavirus Disease 2019. Clin. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- San-Blas, G.; Niño-Vega, G.; Barreto, L.; Hebeler-Barbosa, F.; Bagagli, E.; Olivero de Briceño, R.; Mendes, R.P. Primers for clinical detection of Paracoccidioides brasiliensis. J. Clin. Microbiol. 2005, 43, 4255–4257. [Google Scholar] [CrossRef][Green Version]
- Koishi, A.C.; Vituri, D.F.; Dionízio Filho, P.S.R.; Sasaki, A.A.; Felipe, M.S.S.; Venancio, E.J. A semi-nested PCR assay for molecular detection of Paracoccidioides brasiliensis in tissue samples. Rev. Soc. Bras. Med. Trop. 2010, 43, 728–730. [Google Scholar] [CrossRef]
- Dias, L.; de Carvalho, L.F.; Romano, C.C. Application of PCR in serum samples for diagnosis of paracoccidioidomycosis in the southern Bahia-Brazil. PLoS Negl. Trop Dis. 2012, 6, e1909. [Google Scholar] [CrossRef] [PubMed]
- Alanio, A.; Bretagne, S. Difficulties with molecular diagnostic tests for mould and yeast infections: Where do we stand? Clin. Microbiol. Infect. 2014, 20, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Sepúlveda, V.E.; Márquez, R.; Turissini, D.A.; Goldman, W.E.; Matute, D.R. Genome sequences reveal cryptic speciation in the human pathogen Histoplasma Capsulatum. mBio 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, M.M.; Theodoro, R.C.; Oliveira, F.F.; Machado, G.C.; Hahn, R.C.; Bagagli, E.; San-Blas, G.; Felipe, M.S. Paracoccidioides lutzii sp. nov.: Biological and clinical implications. Med. Mycol. 2014, 52, 19–28. [Google Scholar] [CrossRef]
- Hebert, P.D.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. Biol. Sci./R. Soc. 2003, 270, 313–321. [Google Scholar] [CrossRef]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Consortium, F.B. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Irinyi, L.; Serena, C.; Garcia-Hermoso, D.; Arabatzis, M.; Desnos-Ollivier, M.; Vu, D.; Cardinali, G.; Arthur, I.; Normand, A.C.; Giraldo, A.; et al. International Society of Human and Animal Mycology (ISHAM)-ITS reference DNA barcoding database—The quality controlled standard tool for routine identification of human and animal pathogenic fungi. Med. Mycol. 2015, 53, 313–337. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M., Gelfand, D., Shinsky, J., White, T., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Bialek, R.; Ibricevic, A.; Fothergill, A.; Begerow, D. Small subunit ribosomal DNA sequence shows Paracoccidioides brasiliensis closely related to Blastomyces dermatitidis. J. Clin. Microbiol. 2000, 38, 3190–3193. [Google Scholar] [CrossRef]
- Gomes, G.M.; Cisalpino, P.S.; Taborda, C.P.; de Camargo, Z.P. PCR for diagnosis of paracoccidioidomycosis. J. Clin. Microbiol. 2000, 38, 3478–3480. [Google Scholar] [CrossRef]
- Cisalpino, P.S.; Puccia, R.; Yamauchi, L.M.; Cano, M.I.N.; da Silveira, J.F.; Travassos, L.R. Cloning, characterization, and epitope expression of the major diagnostic antigen of Paracoccidioides brasiliensis. J. Biol. Chem. 1996, 271, 4553–4560. [Google Scholar] [CrossRef] [PubMed]
- Imai, T.; Sano, A.; Mikami, Y.; Watanabe, K.; Aoki, F.H.; Branchini, M.L.; Negroni, R.; Nishimura, K.; Miyaji, M. A new PCR primer for the identification of Paracoccidioides brasiliensis based on rRNA sequences coding the internal transcribed spacers (ITS) and 5 × 8S regions. Med. Mycol. 2000, 38, 323–326. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Iwen, P.C.; Hinrichs, S.H.; Rupp, M.E. Utilization of the internal transcribed spacer regions as molecular targets to detect and identify human fungal pathogens. Med. Mycol. 2002, 40, 87–109. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Rodrigues, A.M.; Feng, P.; Hoog, G.S. Global ITS diversity in the Sporothrix schenckii complex. Fungal Divers. 2014, 66, 153–165. [Google Scholar] [CrossRef]
- Hebeler-Barbosa, F.; Morais, F.V.; Montenegro, M.R.; Kuramae, E.E.; Montes, B.; McEwen, J.G.; Bagagli, E.; Puccia, R. Comparison of the sequences of the internal transcribed spacer regions and PbGP43 genes of Paracoccidioides brasiliensis from patients and armadillos (Dasypus novemcinctus). J. Clin. Microbiol. 2003, 41, 5735. [Google Scholar] [CrossRef] [PubMed]
- Maiden, M.C.; Bygraves, J.A.; Feil, E.; Morelli, G.; Russell, J.E.; Urwin, R.; Zhang, Q.; Zhou, J.; Zurth, K.; Caugant, D.A.; et al. Multilocus sequence typing: A portable approach to the identification of clones within populations of pathogenic microorganisms. Proc. Natl. Acad. Sci. USA 1998, 95, 3140–3145. [Google Scholar] [CrossRef] [PubMed]
- Glaeser, S.P.; Kämpfer, P. Multilocus sequence analysis (MLSA) in prokaryotic taxonomy. Syst. Appl. Microbiol. 2015, 38, 237–245. [Google Scholar] [CrossRef]
- Maiden, M.C. Multilocus sequence typing of bacteria. Annu. Rev. Microbiol. 2006, 60, 561–588. [Google Scholar] [CrossRef]
- Carrero, L.L.; Nino-Vega, G.; Teixeira, M.M.; Carvalho, M.J.; Soares, C.M.; Pereira, M.; Jesuino, R.S.; McEwen, J.G.; Mendoza, L.; Taylor, J.W.; et al. New Paracoccidioides brasiliensis isolate reveals unexpected genomic variability in this human pathogen. Fungal Genet. Biol. 2008, 45, 605–612. [Google Scholar] [CrossRef]
- Mullis, K.B.; Faloona, F.A. Specific synthesis of DNA in vitro via a polymerase-catalyzed chain reaction. Methods Enzymol. 1987, 155, 335–350. [Google Scholar] [CrossRef]
- Waters, D.L.; Shapter, F.M. The polymerase chain reaction (PCR): General methods. Methods Mol. Biol. 2014, 1099, 65–75. [Google Scholar] [CrossRef]
- Goldani, L.Z.; Maia, A.L.; Sugar, A.M. Cloning and nucleotide sequence of a specific DNA fragment from Paracoccidioides brasiliensis. J. Clin. Microbiol. 1995, 33, 1652–1654. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, G.S.; Aleff, R.A.; Kline, B.C.; da Silva Lacaz, C. Molecular detection and identification of Paracoccidioides brasiliensis. J. Clin. Microbiol. 1997, 35, 1894–1896. [Google Scholar] [CrossRef] [PubMed]
- Motoyama, A.B.; Venancio, E.J.; Brandao, G.O.; Petrofeza-Silva, S.; Pereira, I.S.; Soares, C.M.; Felipe, M.S. Molecular identification of Paracoccidioides brasiliensis by PCR amplification of ribosomal DNA. J. Clin. Microbiol. 2000, 38, 3106–3109. [Google Scholar] [CrossRef] [PubMed]
- Buitrago, M.J.; Merino, P.; Puente, S.; Gomez-Lopez, A.; Arribi, A.; Zancope-Oliveira, R.M.; Gutierrez, M.C.; Rodriguez-Tudela, J.L.; Cuenca-Estrella, M. Utility of real-time PCR for the detection of Paracoccidioides brasiliensis DNA in the diagnosis of imported paracoccidioidomycosis. Med. Mycol. 2009, 47, 879–882. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pinheiro, B.G. Development of New Molecular Tools for the Identification of Paracoccidioides spp.; Universidade Federal de São Paulo: São Paulo, Brazil, 2020. [Google Scholar]
- Bialek, R.; Ibricevic, A.; Aepinus, C.; Najvar, L.K.; Fothergill, A.W.; Knobloch, J.; Graybill, J.R. Detection of Paracoccidioides brasiliensis in tissue samples by a nested PCR assay. J. Clin. Microbiol. 2000, 38, 2940–2942. [Google Scholar] [CrossRef] [PubMed]
- Sano, A.; Yokoyama, K.; Tamura, M.; Mikami, Y.; Takahashi, I.; Fukushima, K.; Miyaji, M.; Nishimura, K. Detection of gp43 and ITS1-5.8S-ITS2 ribosomal RNA genes of Paracoccidioides brasiliensis in paraffin-embedded tissue. Jpn. J. Med. Mycol. 2001, 42, 23–27. [Google Scholar] [CrossRef]
- Ricci, G.; Zelck, U.; Mota, F.; Lass-Florl, C.; Franco, M.F.; Bialek, R. Genotyping of Paracoccidioides brasiliensis directly from paraffin embedded tissue. Med. Mycol. 2008, 46, 31–34. [Google Scholar] [CrossRef]
- Correia, J.; de Moraes Borba, C.; Reis, B.; Martins, A.; Unkles, S.; Kinghorn, J.R.; Lucena-Silva, N. The ceja-1 sequence as a potential new molecular marker for Paracoccidioides brasiliensis infection. Mycoses 2010, 53, 130–137. [Google Scholar] [CrossRef]
- Gaviria, M.; Rivera, V.; Muñoz-Cadavid, C.; Cano, L.E.; Naranjo, T.W. Validation and clinical application of a nested PCR for paracoccidioidomycosis diagnosis in clinical samples from Colombian patients. Braz. J. Infect. Dis. 2015, 19, 376–383. [Google Scholar] [CrossRef]
- Pitz Ade, F.; Koishi, A.C.; Tavares, E.R.; Andrade, F.G.; Loth, E.A.; Gandra, R.F.; Venancio, E.J. An optimized one-tube, semi-nested PCR assay for Paracoccidioides brasiliensis detection. Rev. Soc. Bras. Med. Trop. 2013, 46, 783–785. [Google Scholar] [CrossRef] [PubMed]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar] [PubMed]
- Gil-Lamaignere, C.; Roilides, E.; Hacker, J.; Müller, F.M.C. Molecular typing for fungi—A critical review of the possibilities and limitations of currently and future methods. Clin. Microbiol. Infect. 2003, 9, 172–185. [Google Scholar] [CrossRef] [PubMed]
- Nino-Vega, G.A.; Calcagno, A.M.; San-Blas, G.; San-Blas, F.; Gooday, G.W.; Gow, N.A. RFLP analysis reveals marked geographical isolation between strains of Paracoccidioides brasiliensis. Med. Mycol. 2000, 38, 437–441. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Roberto, T.N.; Rodrigues, A.M.; Hahn, R.C.; de Camargo, Z.P. Identifying Paracoccidioides phylogenetic species by PCR-RFLP of the alpha-tubulin gene. Med. Mycol. 2016, 54, 240–247. [Google Scholar] [CrossRef]
- Williams, J.G.; Kubelik, A.R.; Livak, K.J.; Rafalski, J.A.; Tingey, S.V. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res. 1990, 18, 6531–6535. [Google Scholar] [CrossRef]
- Soares, C.M.; Madlun, E.E.; da Silva, S.P.; Pereira, M.; Felipe, M.S. Characterization of Paracoccidioides brasiliensis isolates by random amplified polymorphic DNA analysis. J. Clin. Microbiol. 1995, 33, 505–507. [Google Scholar] [CrossRef]
- Calcagno, A.M.; Niño-Vega, G.; San-Blas, F.; San-Blas, G. Geographic discrimination of Paracoccidioides brasiliensis strains by randomly amplified polymorphic DNA analysis. J. Clin. Microbiol. 1998, 36, 1733–1736. [Google Scholar] [CrossRef]
- Totti, D.O.; Romanha, A.J.; Grisard, E.C.; Simpson, A.J.; Koury, M.C. Random amplified polymorphic DNA (RAPD) analysis of Paracoccidioides brasiliensis isolates. Rev. Latinoam. Microbiol. 1999, 41, 139–143. [Google Scholar]
- Hahn, R.C.; Macedo, A.M.; Fontes, C.J.; Batista, R.D.; Santos, N.L.; Hamdan, J.S. Randomly amplified polymorphic DNA as a valuable tool for epidemiological studies of Paracoccidioides brasiliensis. J. Clin. Microbiol. 2003, 41, 2849–2854. [Google Scholar] [CrossRef]
- Molinari-Madlum, E.E.; Felipe, M.S.; Soares, C.M. Virulence of Paracoccidioides brasiliensis isolates can be correlated to groups defined by random amplified polymorphic DNA analysis. Med. Mycol. 1999, 37, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Motta, T.R.; Moreira-Filho, C.A.; Mendes, R.P.; Souza, L.R.; Sugizak, M.F.; Baueb, S.; Calich, V.L.; Vaz, C.A. Evaluation of DNA polymorphisms amplified by arbitrary primers (RAPD) as genetically associated elements to differentiate virulent and non-virulent Paracoccidioides brasiliensis isolates. FEMS Immunol. Med. Microbiol. 2002, 33, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Sano, A.; Tanaka, R.; Yokoyama, K.; Franco, M.; Bagagli, E.; Montenegro, M.R.; Mikami, Y.; Miyaji, M.; Nishimura, K. Comparison between human and armadillo Paracoccidioides brasiliensis isolates by random amplified polymorphic DNA analysis. Mycopathologia 1998, 143, 165–169. [Google Scholar] [CrossRef]
- Hahn, R.C.; Macedo, A.M.; Santos, N.L.; Resende, J.C.; Hamdan, J.S. Characterization of Paracoccidioides brasiliensis atypical isolates by random amplified polymorphic DNA analysis. Rev. Iberoam. Micol. 2002, 19, 49–51. [Google Scholar] [PubMed]
- Batista Junior, J.; Berzaghi, R.; Arnaud, A.D.; Fontes, C.J.; de Camargo, Z.P.; Hahn, R.C. Simultaneous infection of human host with genetically distinct isolates of Paracoccidioides brasiliensis. Mem. Inst. Oswaldo Cruz 2010, 105, 62–65. [Google Scholar] [CrossRef] [PubMed]
- Ellegren, H. Microsatellites: Simple sequences with complex evolution. Nat. Rev. Genet. 2004, 5, 435–445. [Google Scholar] [CrossRef]
- Jeffreys, A.J.; Wilson, V.; Thein, S.L. Individual-specific ‘fingerprints’ of human DNA. Nature 1985, 316, 76–79. [Google Scholar] [CrossRef]
- Bart-Delabesse, E.; Sarfati, J.; Debeaupuis, J.-P.; Leeuwen, W.; van Belkum, A.; Bretagne, S.; Latge, J. Comparison of restriction fragment length polymorphism, microsatellite length polymorphism, and random amplification of polymorphic DNA analyses for fingerprinting Aspergillus fumigatus isolates. J. Clin. Microbiol. 2001, 39, 2683–2686. [Google Scholar] [CrossRef]
- Foulet, F.; Nicolas, N.; Eloy, O.; Botterel, F.; Gantier, J.C.; Costa, J.M.; Bretagne, S. Microsatellite marker analysis as a typing system for Candida glabrata. J. Clin. Microbiol. 2005, 43, 4574–4579. [Google Scholar] [CrossRef]
- Nascimento, E.; Martinez, R.; Lopes, A.R.; de Souza Bernardes, L.A.; Barco, C.P.; Goldman, M.H.S.; Taylor, J.W.; McEwen, J.G.; Nobrega, M.P.; Nobrega, F.G.; et al. Detection and selection of microsatellites in the genome of Paracoccidioides brasiliensis as molecular markers for clinical and epidemiological studies. J. Clin. Microbiol. 2004, 42, 5007–5014. [Google Scholar] [CrossRef]
- Matute, D.R.; Sepulveda, V.E.; Quesada, L.M.; Goldman, G.H.; Taylor, J.W.; Restrepo, A.; McEwen, J.G. Microsatellite analysis of three phylogenetic species of Paracoccidioides brasiliensis. J. Clin. Microbiol. 2006, 44, 2153–2157. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ravindran, S. Barbara McClintock and the discovery of jumping genes. Proc. Natl. Acad. Sci. USA 2012, 109, 20198–20199. [Google Scholar] [CrossRef] [PubMed]
- Alves, F.L.; Ribeiro, M.A.; Hahn, R.C.; de Melo Teixeira, M.; de Camargo, Z.P.; Cisalpino, P.S.; Marini, M.M. Transposable elements and two other molecular markers as typing tools for the genus Paracoccidioides. Med. Mycol. 2015, 53, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, C.A.; Champion, M.D.; Holder, J.W.; Muszewska, A.; Goldberg, J.; Bailao, A.M.; Brigido, M.M.; Ferreira, M.E.; Garcia, A.M.; Grynberg, M.; et al. Comparative genomic analysis of human fungal pathogens causing paracoccidioidomycosis. PLoS Genet. 2011, 7, e1002345. [Google Scholar] [CrossRef]
- Marini, M.M.; Zanforlin, T.; Santos, P.C.; Barros, R.R.; Guerra, A.C.; Puccia, R.; Felipe, M.S.; Brigido, M.; Soares, C.M.; Ruiz, J.C.; et al. Identification and characterization of Tc1/mariner-like DNA transposons in genomes of the pathogenic fungi of the Paracoccidioides species complex. BMC Genom. 2010, 11, 130. [Google Scholar] [CrossRef]
- Higuchi, R.; Fockler, C.; Dollinger, G.; Watson, R. Kinetic PCR analysis: Real-time monitoring of DNA Amplification reactions. Biotechnology 1993, 11, 1026–1030. [Google Scholar] [CrossRef]
- Chong, G.-L.M.; van de Sande, W.W.J.; Dingemans, G.J.H.; Gaajetaan, G.R.; Vonk, A.G.; Hayette, M.-P.; van Tegelen, D.W.E.; Simons, G.F.M.; Rijnders, B.J.A. Validation of a new Aspergillus real-time PCR assay for direct detection of Aspergillus and azole resistance of Aspergillus fumigatus on bronchoalveolar lavage fluid. J. Clin. Microbiol. 2015, 53, 868. [Google Scholar] [CrossRef]
- Scherer, E.; Iriart, X.; Bellanger, A.P.; Dupont, D.; Guitard, J.; Gabriel, F.; Cassaing, S.; Charpentier, E.; Guenounou, S.; Cornet, M.; et al. Quantitative PCR (qPCR) detection of Mucorales DNA in bronchoalveolar lavage fluid to diagnose pulmonary mucormycosis. J. Clin. Microbiol. 2018, 56, e00289-18. [Google Scholar] [CrossRef]
- Semighini, C.P.; de Camargo, Z.P.; Puccia, R.; Goldman, M.H.; Goldman, G.H. Molecular identification of Paracoccidioides brasiliensis by 5′ nuclease assay. Diagn Microbiol. Infect. Dis. 2002, 44, 383–386. [Google Scholar] [CrossRef]
- Rocha-Silva, F.; Gomes, L.I.; Gracielle-Melo, C.; Goes, A.M.; Caligiorne, R.B. Real Time Polymerase Chain Reaction (rt-PCR): A new patent to diagnostic purposes for paracoccidioidomycosis. Recent Pat. Endocr. Metab. Immune Drug Discov. 2017, 10, 143–149. [Google Scholar] [CrossRef]
- Rocha-Silva, F.; Guimarães, C.F.; de Oliveira Júnior, E.R.; de Figueiredo, S.M.; Caligiorne, R.B. Disseminated paracoccidioidomycosis prediagnosticated as neoplasm: An important challenge in diagnosis using rt-PCR. Med. Mycol. Case Rep. 2018, 19, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.V.; Landgraf, T.N.; Corrêa, P.C.; Souza, I.E.L.; Fernandes, F.F.; Panunto-Castelo, A. Quantitation of pulmonary fungal burden in Paracoccidioides brasiliensis-infected mice by real-time PCR. Rev. Inst. Med. Trop. Sao Paulo 2019, 61. [Google Scholar] [CrossRef]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28, E63. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, H.; Iwase, T.; Omagari, D.; Asano, M.; Nakamura, R.; Ueki, K.; Shinozuka, K.; Kaneko, T.; Tonogi, M.; Ohki, H. Rapid detection of Candida albicans in oral exfoliative cytology samples by loop-mediated isothermal amplification. J. Oral Sci. 2017, 59, 541–547. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Endo, S.; Komori, T.; Ricci, G.; Sano, A.; Yokoyama, K.; Ohori, A.; Kamei, K.; Franco, M.; Miyaji, M.; Nishimura, K. Detection of gp43 of Paracoccidioides brasiliensis by the loop-mediated isothermal amplification (LAMP) method. FEMS Microbiol. Lett. 2004, 234, 93–97. [Google Scholar] [CrossRef]
- Tatibana, B.T.; Sano, A.; Uno, J.; Kamei, K.; Igarashi, T.; Mikami, Y.; Miyaji, M.; Nishimura, K.; Itano, E.N. Detection of Paracoccidioides brasiliensis gp43 gene in sputa by loop-mediated isothermal amplification method. J. Clin. Lab. Anal. 2009, 23, 139–143. [Google Scholar] [CrossRef]
- Carvajal, D.M.L. Development of a Rapid Diagnosis Test for Identification of Paracoccidioides lutzii, Using the Loop-Mediated Isothermal Amplification (LAMP) Technique; Universidade Estadual de Campinas: Campinas, Brazil, 2018. [Google Scholar]
- Pinkel, D.; Straume, T.; Gray, J.W. Cytogenetic analysis using quantitative, high-sensitivity, fluorescence hybridization. Proc. Natl. Acad. Sci. USA 1986, 83, 2934–2938. [Google Scholar] [CrossRef]
- Da Silva, R.M., Jr.; Da Silva Neto, J.R.; Santos, C.S.; Frickmann, H.; Poppert, S.; Cruz, K.S.; Koshikene, D.; De Souza, J.V.B. Evaluation of fluorescence in situ hybridisation (FISH) for the detection of fungi directly from blood cultures and cerebrospinal fluid from patients with suspected invasive mycoses. Ann. Clin. Microbiol. Antimicrob. 2015, 14, 6. [Google Scholar] [CrossRef]
- Kempf, V.A.; Trebesius, K.; Autenrieth, I.B. Fluorescent In situ hybridization allows rapid identification of microorganisms in blood cultures. J. Clin. Microbiol. 2000, 38, 830–838. [Google Scholar] [CrossRef]
- Moter, A.; Göbel, U.B. Fluorescence in situ hybridization (FISH) for direct visualization of microorganisms. J. Microbiol. Methods 2000, 41, 85–112. [Google Scholar] [CrossRef]
- Rigby, S.; Procop, G.W.; Haase, G.; Wilson, D.; Hall, G.; Kurtzman, C.; Oliveira, K.; Von Oy, S.; Hyldig-Nielsen, J.J.; Coull, J.; et al. Fluorescence in situ hybridization with peptide nucleic acid probes for rapid identification of Candida albicans directly from blood culture bottles. J. Clin. Microbiol. 2002, 40, 2182–2186. [Google Scholar] [CrossRef] [PubMed]
- Procop, G.W. Molecular diagnostics for the detection and characterization of microbial pathogens. Clin. Infect. Dis. 2007, 45 (Suppl. 2), S99–S111. [Google Scholar] [CrossRef] [PubMed]
- Buongiorno-Nardelli, M.; Amaldi, F. Autoradiographic detection of molecular hybrids between rRNA and DNA in tissue sections. Nature 1970, 225, 946–948. [Google Scholar] [CrossRef]
- De Brito, T.; Sandhu, G.S.; Kline, B.C.; Aleff, R.A.; Sandoval, M.P.; Santos, R.T.; Brandão, A.A.H.; Lacaz, C.S. In situ hybridization in paracoccidioidomycosis. Med. Mycol. 1999, 37, 207–211. [Google Scholar] [CrossRef]
- Arantes, T.D.; Theodoro, R.C.; Teixeira Mde, M.; Bosco Sde, M.; Bagagli, E. Environmental mapping of Paracoccidioides spp. in Brazil reveals new clues into genetic diversity, biogeography and wild host association. PLoS Negl. Trop Dis. 2016, 10, e0004606. [Google Scholar] [CrossRef]
- Arantes, T.D.; Theodoro, R.C.; Teixeira, M.d.M.; Bagagli, E. Use of fluorescent oligonucleotide probes for differentiation between Paracoccidioides brasiliensis and Paracoccidioides lutzii in yeast and mycelial phase. Mem. Inst. Oswaldo Cruz 2017, 112, 140–145. [Google Scholar] [CrossRef]
- Karas, M.; Bachmann, D.; Hillenkamp, F. Influence of the wavelength in high-irradiance ultraviolet laser desorption mass spectrometry of organic molecules. Anal. Chem. 1985, 57, 2935–2939. [Google Scholar] [CrossRef]
- Claydon, M.A.; Davey, S.N.; Edwards-Jones, V.; Gordon, D.B. The rapid identification of intact microorganisms using mass spectrometry. Nat. Biotechnol. 1996, 14, 1584–1586. [Google Scholar] [CrossRef]
- Sauget, M.; Valot, B.; Bertrand, X.; Hocquet, D. Can MALDI-TOF mass spectrometry reasonably type bacteria? Trends Microbiol. 2017, 25, 447–455. [Google Scholar] [CrossRef]
- Singhal, N.; Kumar, M.; Kanaujia, P.K.; Virdi, J.S. MALDI-TOF mass spectrometry: An emerging technology for microbial identification and diagnosis. Front. Microbiol. 2015, 6, 791. [Google Scholar] [CrossRef]
- Rychert, J.; Slechta, E.S.; Barker, A.P.; Miranda, E.; Babady, N.E. Multicenter Evaluation of the Vitek MS v3.0 System for the identification of filamentous fungi. J. Clin. Microbiol. 2018, 56, e01353-17. [Google Scholar] [CrossRef]
- Lacroix, C.; Gicquel, A.; Sendid, B.; Meyer, J.; Accoceberry, I.; François, N.; Morio, F.; Desoubeaux, G.; Chandenier, J.; Kauffmann-Lacroix, C.; et al. Evaluation of two matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF MS) systems for the identification of Candida species. Clin. Microbiol. Infect. 2014, 20, 153–158. [Google Scholar] [CrossRef] [PubMed]
- De Carolis, E.; Posteraro, B.; Lass-Flörl, C.; Vella, A.; Florio, A.R.; Torelli, R.; Girmenia, C.; Colozza, C.; Tortorano, A.M.; Sanguinetti, M.; et al. Species identification of Aspergillus, Fusarium and Mucorales with direct surface analysis by matrix-assisted laser desorption ionization time-of-flight mass spectrometry. Clin. Microbiol. Infect. 2012, 18, 475–484. [Google Scholar] [CrossRef] [PubMed]
- De Almeida, J.N.; Del Negro, G.M.B.; Grenfell, R.C.; Vidal, M.S.M.; Thomaz, D.Y.; de Figueiredo, D.S.Y.; Bagagli, E.; Juliano Neto, L.; Benard, G. MALDI-TOF mass spectrometry for rapid identification of the dimorphic fungi Paracoccidioides brasiliensis and Paracoccidioides lutzii. J. Clin. Microbiol. 2015, 53, 1383–1386. [Google Scholar] [CrossRef]
- Naumann, A. Fourier Transform Infrared (FTIR) microscopy and imaging of fungi. In Advanced Microscopy in Mycology; Dahms, T., Czymmek, K., Eds.; Fungal Biology; Springer: Cham, Switzerland, 2015. [Google Scholar] [CrossRef]
- Cooley, J.W.; Tukey, J.W. An algorithm for the machine calculation of complex Fourier series. Math. Comput. 1965, 19, 297–301. [Google Scholar]
- Costa, F.; Silva, P.; Morais, C.; Arantes, T.; Milan, E.; Theodoro, R.; Lima, K. Attenuated Total Reflection Fourier Transform-Infrared (ATR-FTIR) spectroscopy as a new technology for discrimination between Cryptococcus neoformans and Cryptococcus gattii. Anal. Methods 2016, 8, 7107–7115. [Google Scholar] [CrossRef]
- Erukhimovitch, V.; Pavlov, V.; Talyshinsky, M.; Souprun, Y.; Huleihel, M. FTIR microscopy as a method for identification of bacterial and fungal infections. J. Pharm. Biomed. Anal. 2005, 37, 1105–1108. [Google Scholar] [CrossRef]
- Comparato Filho, O.O.; Morais, F.V.; Bhattacharjee, T.; Castilho, M.L.; Raniero, L. Rapid identification of Paracoccidioides lutzii and P. brasiliensis using Fourier Transform Infrared spectroscopy. J. Mol. Struct. 2019, 1177, 152–159. [Google Scholar] [CrossRef]
- Dietrich, D.; Uhl, B.; Sailer, V.; Holmes, E.E.; Jung, M.; Meller, S.; Kristiansen, G. Improved PCR performance using template DNA from formalin-fixed and paraffin-embedded tissues by overcoming PCR inhibition. PLoS ONE 2013, 8, e77771. [Google Scholar] [CrossRef]
- Brummer, E.; Castaneda, E.; Restrepo, A. Paracoccidioidomycosis: An update. Clin. Microbiol. Rev. 1993, 6, 89–117. [Google Scholar] [CrossRef]
- Morais, F.V.; Barros, T.F.; Fukada, M.K.; Cisalpino, P.S.; Puccia, R. Polymorphism in the gene coding for the immunodominant antigen gp43 from the pathogenic fungus Paracoccidioides brasiliensis. J. Clin. Microbiol. 2000, 38, 3960. [Google Scholar] [CrossRef]
- Misas, E.; Gómez, O.M.; Botero, V.; Muñoz, J.F.; Teixeira, M.M.; Gallo, J.E.; Clay, O.K.; McEwen, J.G. Updates and comparative analysis of the mitochondrial genomes of Paracoccidioides spp. using Oxford Nanopore MinION sequencing. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Kozel, T.R.; Burnham-Marusich, A.R. Point-of-Care testing for infectious diseases: Past, present, and future. J. Clin. Microbiol. 2017, 55, 2313–2320. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinheiro, B.G.; Hahn, R.C.; Camargo, Z.P.d.; Rodrigues, A.M. Molecular Tools for Detection and Identification of Paracoccidioides Species: Current Status and Future Perspectives. J. Fungi 2020, 6, 293. https://doi.org/10.3390/jof6040293
Pinheiro BG, Hahn RC, Camargo ZPd, Rodrigues AM. Molecular Tools for Detection and Identification of Paracoccidioides Species: Current Status and Future Perspectives. Journal of Fungi. 2020; 6(4):293. https://doi.org/10.3390/jof6040293
Chicago/Turabian StylePinheiro, Breno Gonçalves, Rosane Christine Hahn, Zoilo Pires de Camargo, and Anderson Messias Rodrigues. 2020. "Molecular Tools for Detection and Identification of Paracoccidioides Species: Current Status and Future Perspectives" Journal of Fungi 6, no. 4: 293. https://doi.org/10.3390/jof6040293
APA StylePinheiro, B. G., Hahn, R. C., Camargo, Z. P. d., & Rodrigues, A. M. (2020). Molecular Tools for Detection and Identification of Paracoccidioides Species: Current Status and Future Perspectives. Journal of Fungi, 6(4), 293. https://doi.org/10.3390/jof6040293