Global Sexual Fertility in the Opportunistic Pathogen Aspergillus fumigatus and Identification of New Supermater Strains

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Growth Conditions
2.2. DNA Extraction, PCR Mating-Type Diagnostic and RAPD-PCR Analysis
2.3. Crossing of A. fumigatus Isolates and Sexual Fertility
2.4. Heat Shock, Viability and Germination of Ascospores
2.5. Supermater and Pyrimidine Auxotrophic Strains
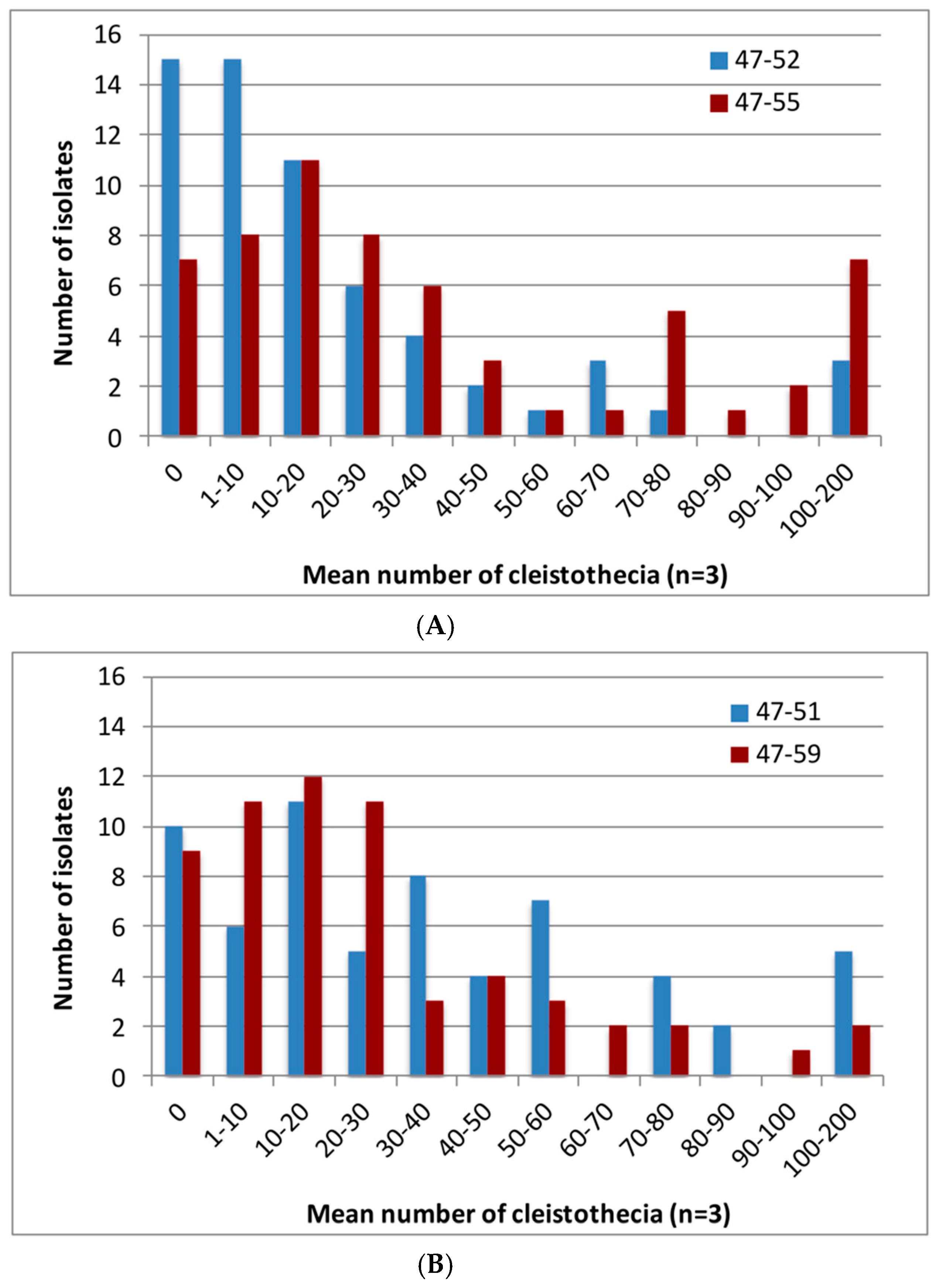
3. Results
3.1. A. fumigatus Worldwide Collection and Mating-Type Distribution
3.2. Sexual Fertility of Global A. fumigatus Isolate Collection
3.3. Sexual Fertility of Isolates Other than from Ireland
3.4. Influence of Heat Shock on Ascospore Germination
3.5. Ascospore Viability and Germination
3.6. New Supermater and Fertile Pyrimidine Auxotrophic Strains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mullins, J.; Harvey, R.; Eaton, A. Sources and incidence of airborne Aspergillus fumigatus (Fres). Clin. Allergy 1976, 6, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Kwon-Chung, K.J.; Sugui, J.A. Aspergillus fumigatus—What makes the species a ubiquitous human fungal pathogen? PLoS Pathog. 2013, 9, e1003743. [Google Scholar] [CrossRef] [PubMed]
- Raper, K.B.; Fennell, D.I. The Genus Aspergillus; Williams and Wilkins: Baltimore, MD, USA, 1965. [Google Scholar]
- Woodcock, A.A.; Steel, N.; Moore, C.B.; Howard, S.J.; Custovic, A.; Denning, D.W. Fungal contamination of bedding. Allergy 2006, 61, 140–142. [Google Scholar] [CrossRef] [PubMed]
- Latgé, J.P. The pathobiology of Aspergillus fumigatus. Trends Microbiol. 2001, 9, 382–389. [Google Scholar] [CrossRef]
- Latgé, J.P. Aspergillus fumigatus and aspergillosis. Clin. Microbiol. Rev. 1999, 12, 310–350. [Google Scholar] [CrossRef]
- Denning, D.W. Invasive aspergillosis. Clin. Infect. Dis. 1998, 26, 781–803. [Google Scholar] [CrossRef]
- Dagenais, T.R.T.; Keller, N.P. Pathogenesis of Aspergillus fumigatus in invasive aspergillosis. Clin. Micro. Revs. 2009, 22, 447–465. [Google Scholar] [CrossRef]
- Kousha, M.; Tadi, R.; Soubani, A.O. Pulmonary aspergillosis: A clinical review. Eur. Resp. Rev. 2011, 20, 156–174. [Google Scholar] [CrossRef]
- Wassano, N.S.; Goldman, G.H.; Damasio, A. Aspergillus fumigatus. Trends Microbiol. 2020, 28, 594–595. [Google Scholar] [CrossRef]
- Balloy, V.; Chignard, M. The innate immune response to Aspergillus fumigatus. Microbiol. Infect. 2009, 11, 919–927. [Google Scholar] [CrossRef]
- Lestrade, P.P.A.; Meis, J.F.; Meis, J.F.; Melchers, W.J.G.; Verweij, P. Triazole resistance in Aspergillus fumigatus: Recent insights and challenges for patient management. Clin. Microbiol. 2019, 25, 799–806. [Google Scholar] [CrossRef]
- del Palacio, A.; Alhambra, A.; Cuetara, M.S.; Ponton, J. Early diagnosis of invasive fungal infections caused by Aspergillus and other emerging mycelial fungi. Rev. Iberoam. Mycol. 2007, 24, 187–197. [Google Scholar] [CrossRef]
- Rivero-Menendez, O.; Alastruey-Izquierdo, A.; Mellado, E.; Cuenca-Estrella, M. Triazole resistance in Aspergillus spp.: A worldwide problem? J. Fungi 2016, 2, 21. [Google Scholar] [CrossRef] [PubMed]
- Verweij, P.E.; Lucas, J.A.; Arendrup, M.C.; Bowyer, P.; Brinkmann, A.J.F.; Denning, D.W.; Dyer, P.S.; Fisher, M.C.; Geenen, P.L.; Gisi, U.; et al. The one health problem of azole resistance in Aspergillus fumigatus: Current insights and future research agenda. Fungal Biol. Revs. 2020, in press. [Google Scholar]
- Arastehfar, A.; Carvalho, A.; van de Veerdonk, F.L. COVID-19 associated pulmonary aspergillosis (CAPA)-from immunology to treatment. J. Fungi. 2020, 6, 91. [Google Scholar] [CrossRef]
- Mohamed, A.; Hassan, T.; Trzos-Grzybowska, M.; Thomas, J.; Quinn, A.; O’Sullivan, M.; Griffin, A.; Rogers, T.R.; Talento, A.F. Multi-triazole-resistant Aspergillus fumigatus and SARS-CoV-2 co-infection: A lethal combination. Med. Mycol. Case Rep. 2020. [Google Scholar] [CrossRef]
- Van Arkel, A.L.E.; Rijpstra, T.A.; Belderbos, H.N.A.; Wijngaarden, P.; Verweij, P.E.; Bentvelsen, R.G. COVID-19-associated pulmonary aspergillosis. Am. J. Resp. Crit. Care Med. 2020, 202, 132–135. [Google Scholar] [CrossRef] [PubMed]
- O’Gorman, C.M.; Fuller, H.; Dyer, P.S. Discovery of a sexual cycle in the opportunistic fungal pathogen Aspergillus fumigatus. Nature 2009, 457, 471–474. [Google Scholar] [CrossRef]
- Galagan, J.E.; Calvo, S.E.; Cuomo, C.; Ma, L.J.; Wortman, J.R.; Batzoglou, S.; Lee, S.I.; Baştürkmen, M.; Spevak, C.C.; Clutterbuck, J.; et al. Sequencing of Aspergillus nidulans and comparative analysis with A. fumigatus and A. oryzae. Nature 2005, 438, 1105–1115. [Google Scholar] [CrossRef]
- Nierman, W.C.; Pain, A.; Anderson, M.J.; Wortman, J.R.; Kim, H.S.; Arroyo, J.; Berriman, M.; Abe, K.; Archer, D.B.; Bermejo, C.; et al. Genomic sequence of the pathogenic and allergenic filamentous fungus Aspergillus fumigatus. Nature 2005, 438, 1151–1156. [Google Scholar] [CrossRef]
- Paoletti, M.; Rydholm, C.; Schwier, E.U.; Anderson, M.J.; Szakacs, G.; Lutzoni, F.; Debeaupuis, J.P.; Latgé, J.P.; Denning, D.W.; Dyer, P.S. Evidence for sexuality in the opportunistic fungal pathogen Aspergillus fumigatus. Curr. Biol. 2005, 15, 1242–1248. [Google Scholar] [CrossRef] [PubMed]
- Dyer, P.S.; Inderbitzin, P.; Debuchy, R. Mating-type structure, function, regulation and evolution in the Pezizomycotina. In Growth, Differentiation and Sexuality, The Mycota I, 3rd ed.; Wendland, J., Ed.; Springer: Cham, Switzerland, 2016; pp. 351–385. [Google Scholar]
- Sugui, J.A.; Losada, L.; Wang, W.; Varga, J.; Ngamskulrungroj, P.; Abu-Asab, M.; Chang, Y.C.; O’Gorman, C.M.; Wickes, B.L.; Nierman, W.C.; et al. Identification and characterization of an Aspergillus fumigatus “Supermater” pair. mBio 2011, 2, e00234-11. [Google Scholar] [CrossRef]
- Camps, S.M.T.; Rijs, A.J.M.M.; Klaassen, C.H.W.; Meis, J.F.; O’Gorman, C.M.; Dyer, P.S.; Melcher, W.J.; Verweij, P.E. Molecular epidemiology of Aspergillus fumigatus isolates harboring the TR43/L98H azole resistance mechanism. J. Clin. Microbiol. 2012, 50, 2674–2680. [Google Scholar] [CrossRef] [PubMed]
- Dyer, P.S.; Paoletti, M. Reproduction in Aspergillus fumigatus: Sexuality in a supposedly asexual species? Med. Mycol. 2005, 43, S7–S14. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Snelders, E.; Zwaan, B.J.; Schoustra, S.E.; Meis, J.F.; Van Dijk, K.; Hagen, F.; Van Der Beek, M.T.; Kampinga, G.A.; Zoll, J.; et al. A novel environmental azole resistance mutation in Aspergillus fumigatus and a possible role of sexual reproduction in its emergence. mBio 2017, 8, e00791-17. [Google Scholar] [CrossRef] [PubMed]
- Geiser, D.M. Sexual structures in Aspergillus: Morphology, importance and genomics. Med. Mycol. 2009, 47, S21–S26. [Google Scholar] [CrossRef]
- Dijksterhuis, J. Heat-resistant ascospores. In Food Mycology, A Multifaceted Approach to Fungi and Food; Dijksterhuis, J., Samson, R.A., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 101–117. [Google Scholar]
- Szewczyk, E.; Krappmann, S. Conserved regulators of mating are essential for Aspergillus fumigatus cleistothecium formation. Euk. Cell. 2010, 9, 774–793. [Google Scholar] [CrossRef]
- Dyer, P.S.; Kück, U. Sex and the imperfect fungi. Microbiol. Spect. 2017, 5. [Google Scholar] [CrossRef]
- Seifert, K.A. Isolation of filamentous fungi. In Isolation of Biotechnological Organisms from Nature; Labeda, D.P., Ed.; McGraw-Hill: New York, NY, USA, 1990; pp. 21–51. [Google Scholar]
- O’Donnell, K.; Peterson, S.W. Isolation, preservation and taxonomy. In Biotechnology of Filamentous Fungi; Finkelstein, D.B., Ball, C., Eds.; Butterworth-Heinemann: Boston, MA, USA, 1992; pp. 7–43. [Google Scholar]
- Chazalet, V.; Debeaupuis, J.P.; Sarfati, J.; Lortholary, J.; Ribaud, P.; Shah, P.; Cornet, M.; Thien, H.V.; Gluckman, E.; Brücker, G.; et al. Molecular typing of environmental and patient isolates of Aspergillus fumigatus from various hospital settings. J. Clin. Microbiol. 1998, 36, 1494–1500. [Google Scholar] [CrossRef]
- Samson, R.A.; Varga, J.; Dyer, P.S. Morphology and reproductive mode of Aspergillus fumigatus. In Aspergillus fumigatus and Aspergillosis; Latgé, J.P., Steinbach, W.J., Eds.; ASM Press: Washington, DC, USA, 2009; pp. 7–13. [Google Scholar]
- Samson, R.A.; Hong, S.; Peterson, S.; Frisvad, J.C.; Varga, J. Polyphasic taxonomy of Aspergillus section Fumigati and its teleomorph Neosartorya. Stud. Mycol. 2007, 59, 147–203. [Google Scholar] [CrossRef]
- Swilaiman, S.S.; O’Gorman, C.M.; Balajee, S.A.; Dyer, P.S. Discovery of a sexual cycle in Aspergillus lentulus, a close relative of A. fumigatus. Euk. Cell. 2013, 12, 962–969. [Google Scholar] [CrossRef]
- Swilaiman, S.S. Sexual Potential and Population Biology of Fungal Aspergillus and Penicillium Species. Ph.D. Thesis, University of Nottingham, Nottingham, UK, 2013. [Google Scholar]
- Ashton, G.D.; Dyer, P.S. Culturing and mating of Aspergillus fumigatus. Curr. Protocols Micro. 2019, 54, e87. [Google Scholar] [CrossRef]
- Korf, R.P. Two techniques for showing spore markings in operculate discomycetes. Trans. Br. Mycol. Soc. 1952, 35, 24–25. [Google Scholar] [CrossRef]
- Brock, M.; Gehrke, A.; Sugareva, V.; Brakhage, A.A. Promoter analysis and generation of knock-out mutants in Aspergillus fumigatus. In Medical Mycology Cellular and Molecular Techniques; Kavanagh, K., Ed.; John Wiley & Sons, Ltd.: Chichester, UK, 2007; pp. 213–256. [Google Scholar]
- He, Z.M.; Price, M.S.; O’Brian, G.R.; Georgianna, D.R.; Payne, G.A. Improved protocols for functional analysis in the pathogenic fungus Aspergillus flavus. BMC Microbiol. 2007, 7, 104. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.T.; Ho, Q.N.; Do, L.T.B.X.; Mai, L.T.D.; Pham, D.N.; Tran, H.T.T.; Le, D.H.; Nguyen, H.Q.; Tran, V.T. A new and efficient approach for construction of uridine/uracil auxotrophic mutants in the filamentous fungus Aspergillus oryzae using Agrobacteium rumefaciens-mediated transformation. World J. Microbiol. Biotechnol. 2017, 33, 107. [Google Scholar] [CrossRef]
- Geib, E.; Baldeweg, F.; Doerfer, M.; Nett, M.; Brock, M. Cross-chemistry leads to product diversity from atromentin synthetases in aspergilli from Section Nigri. Cell. Chem. Biol. 2019, 26, 223–234. [Google Scholar] [CrossRef]
- Robellet, X.; Oestreicher, N.; Guitton, A.; Vélot, C. Gene silencing of transgenes inserted in the Aspergillus nidulans alcM and/or alcS loci. Curr. Genet. 2010, 56, 341–378. [Google Scholar] [CrossRef]
- Sun, X.Y.; Zhu, J.F.; Bao, L.; Hu, C.C.; Jin, C.; Harris, S.D.; Liu, H.W.; Li, S. pyrG is required for maintaining stable cellular uracil level and normal sporulation pattern under excess uracil stress in Aspergillus nidulans. Sci. China Life Sci. 2013, 56, 467–475. [Google Scholar] [CrossRef][Green Version]
- Fedorova, N.D.; Khaldi, N.; Joardar, V.S.; Maiti, R.; Amedeo, P.; Andreson, M.J.; Crabtree, J.; Silva, J.C.; Badger, J.H.; Albarraq, A.; et al. Genomic islands in the pathogenic filamentous fungus Aspergillus fumigatus. PLoS Genet. 2008, 4, e1000046. [Google Scholar] [CrossRef]
- Garcia-Rubio, R.; Monzon, S.; Alcazar-Fuoli, L.; Cuesta, I.; Mellado, E. Genome-wide comparative analysis of Aspergillus fumigatus strains: The reference genome as a matter of concern. Genes 2018, 9, 363. [Google Scholar] [CrossRef]
- Bertuzzi, M.; Van Rhijn, N.; Krappmann, S.; Bowyer, P.; Bromley, M.J.; Bignell, E.M. On the lineage of Aspergillus fumigatus isolates in common laboratory use. Med. Mycol. 2020, in press. [Google Scholar] [CrossRef]
- Alves de Castro, P.; Valero, C.; Chiaratto, J.; Colabardini, A.C.; Pardeshi, L.; Silva, L.P.; Almeida, F.; Rocha, M.C.; Silva, R.N.; Malavazi, I.; et al. Novel biological functions for the Aspergillus fumigatus NsdC transcription factor homologue involved in sexual development. Front. Fung. Biol. 2020. submitted. [Google Scholar]
- Dyer, P.S.; Hansen, J.; Delaney, A.; Lucas, J.A. Genetic control of resistance to the DMI fungicide prochloraz in the cereal eyespot pathogen Tapesia yallundae. App. Environ. Microbiol. 2000, 66, 4599–4604. [Google Scholar] [CrossRef]
- Xu, J. Estimating the spontaneous mutation rate of loss of sex in the human pathogenic fungus Cryptococcus neoformans. Genetics 2002, 162, 1157–1167. [Google Scholar]
- Takan, J.P.; Chipili, J.; Muthumeenakshi, S.; Talbot, N.J.; Manyasa, E.O.; Bandyopadhyay, R.; Sere, Y.; Nutsugah, S.K.; Talhinhas, P.; Hossain, M.; et al. Magnaporthe oryzae populations adapted to finger millet and rice exhibit distinctive patterns of genetic diversity, sexuality and host interaction. Mol. Biotechnol. 2012, 50, 145–158. [Google Scholar]
- King, K.M.; West, J.S.; Brunner, P.C.; Dyer, P.S.; Fitt, B.D.L. Evolutionary relationships between Rhynchosporium lolii sp. nov. and other Rhynchosporium species on grasses. PLoS ONE 2013, 8, e72536. [Google Scholar]
- Hughes, T.J.; O’Donnell, K.; Sink, S.; Rooney, A.P.; Scandiani, M.M.; Luque, A.; Bhattacharyya, M.K.; Huang, X. Genetic architecture and evolution of the mating type locus in fusaria that cause soybean sudden death syndrome and bean root rot. Mycologia 2014, 106, 686–697. [Google Scholar] [CrossRef]
- Short, D.P.G.; O’Donnell, K.; Thrane, U.; Nielsen, K.F.; Zhang, N.; Juba, J.H.; Geiser, D.M. Phylogenetic relationships among members of the Fusarium solani species complex in human infections and the descriptions of F. keratoplasticum sp. nov. and F. petroliphilum stat. nov. Fungal Genet. Biol. 2013, 53, 59–70. [Google Scholar] [CrossRef]
- Sugui, J.A.; Vinh, D.C.; Nardone, G.; Shea, Y.R.; Chang, Y.C.; Zelazyn, A.M.; Marr, K.A.; Holland, S.M.; Kwon-Chung, K.J. Neosartorya udagawae (Aspergillus udagawae), an emerging agent of aspergillosis: How different is it from Aspergillus fumigatus? J. Clin. Microbiol. 2010, 48, 220–228. [Google Scholar] [CrossRef]
- Horn, B.W.; Moore, G.G.; Carbone, I. Sexual reproduction in Aspergillus flavus. Mycologia 2009, 101, 423–429. [Google Scholar] [CrossRef]
- Horn, B.W.; Ramirez-Prado, J.H.; Carbone, I. Sexual reproduction and recombination in the aflatoxin-producing fungus Aspergillus parasiticus. Fungal Genet. Biol. 2009, 46, 169–175. [Google Scholar] [CrossRef]
- Dyer, P.S.; Ingram, D.S.; Johnstone, K. The control of sexual morphogenesis in the Ascomycotina. Biol. Rev. 1992, 67, 421–458. [Google Scholar] [CrossRef]
- Pöggeler, S.; Masloff, S.; Jacobsen, S.; Kück, U. Karyotype polymorphism correlates with intraspecific infertility in the homothallic ascomycete Sordaria macrospora. J. Evol. Biol. 2000, 13, 281–289. [Google Scholar] [CrossRef]
- Pyrzak, W.; Miller, K.Y.; Miller, B.L. The mating type protein Mat1-2 from asexual Aspergillus fumigatus drives sexual reproduction in fertile Aspergillus nidulans. Eukaryot. Cell 2008, 7, 1029–1040. [Google Scholar] [CrossRef]
- Fisher, M.C.; Henk, D.A. Sex, drugs and recombination: The wild life of Aspergillus. Mol. Ecol. 2012, 21, 1305–1306. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.Y.; Park, H.M. Expression of sexual genes in Aspergillus fumigatus homogeneous culture produced by vegetative mass mating. J. Microbiol. 2019, 57, 688–693. [Google Scholar] [CrossRef] [PubMed]
- King, K.M.; Hawkins, N.J.; Atkins, S.; Dyer, P.S.; West, J.S.; Fraaije, B.A. First application of loop-mediated isothermal amplification (LAMP) assays for rapid identification of mating type in the heterothallic fungus Aspergillus fumigatus. Mycoses 2019, 62, 812–817. [Google Scholar] [CrossRef] [PubMed]
- Kwon-Chung, K.J.; Sugui, J.A. Sexual reproduction in Aspergillus species of medical or economical importance: Why so fastidious? Trends Microbiol. 2009, 17, 481–487. [Google Scholar] [CrossRef]
- Dettman, J.R.; Jacobson, D.J.; Turner, E.; Pringle, A.; Taylor, J.W. Reproductive isolation and phylogenetic divergence in Neurospora: Comparing methods of species recognition in a model eukaryote. Evolution 2003, 57, 2721–2741. [Google Scholar] [CrossRef] [PubMed]
- Ashu, E.E.; Hagen, F.; Chowdhary, A.; Meis, J.F.; Xu, J. Global population genetic analysis of Aspergillus fumigatus. mSphere 2017, 2, e00019-17. [Google Scholar] [CrossRef]
- Sewell, T.R.; Zhu, J.; Rhodes, J.; Hagen, F.; Meis, J.F.; Fisher, M.C.; Jombart, T. Nonrandom distribution of azole resistance across the global population of Aspergillus fumigatus. mBio 2019, 10, e00392-19. [Google Scholar] [CrossRef]
- McEvoy, I.J.; Stuart, M.R. Temperature tolerance of Aspergillus fischeri var. glaber in canned strawberries. Irish J. Agri. Res. 1970, 9, 59–67. [Google Scholar]
- Dijksterhuis, J.; Samson, R.A. Activation of ascospores by novel food preservation techniques. In Advances in Food Mycology. Advances in Experimental Medicine and Biology; Hocking, A.D., Pitt, J.I., Samson, R.A., Thrane, U., Eds.; Springer: Boston, MA, USA, 2006; Volume 571, pp. 247–260. [Google Scholar]
- Losada, L.; Sugui, J.A.; Eckhaus, M.A. Genetic analysis using an isogenic mating pair of Aspergillus fumigatus identifies azole resistance genes and lack of MAT locus’s role in virulence. PLoS Path. 2015, 11, e1004834. [Google Scholar] [CrossRef]
- Krappmann, S. Gene targeting in filamentous fungi: The benefits of impaired repair. Fungal Biol. Rev. 2007, 21, 25–29. [Google Scholar] [CrossRef]
- Hurst, L.D.; Peck, J.R. Recent advances in understanding of the evolution and maintenance of sex. Trends Ecol. Evol. 1996, 11, 46–52. [Google Scholar] [CrossRef]
- Bain, J.M.; Tavanti, A.; Davidson, A.D.; Jacobsen, M.D.; Shaw, D.; Gow, N.A.R.; Odds, F.C. Multilocus sequence typing of the pathogenic fungus Aspergillus fumigatus. J. Clin. Microbiol. 2007, 45, 1469–1477. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Perez, S.; Blanco, J.L.; Alba, P.; Garcia, M.E. Mating type and invasiveness are significantly associated in Aspergillus fumigatus. Med. Mycol. 2010, 48, 273–277. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Number | MAT1-1 | MAT1-2 |
|---|---|---|---|
| Europe | 39 | 61.5% | 38.5% |
| North America | 29 | 48.3% | 51.7% |
| South America | 7 | 42.9% | 57.1% |
| Asia | 27 | 55.6% | 44.4% |
| Africa | 23 | 43.5% | 56.5% |
| Australia | 3 | 33.3% | 66.7% |
| MAT1-2 | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Asia | Africa | South America | North America | Europe | |||||
| MAT1-1 | India | South Africa | Zimbabwe | Brazil | USA (San Francisco) | USA (Michigan) | Ireland | ||
| Asia | India | 36 ± 7 | 81 ± 57 | 104 ± 61 | 29 ± 19 | 77 ± 47 | 15 ± 4 | 33 ± 28 | |
| China | 24 ± 10 | 113 ± 37 | 192 ± 14 | 180 ± 13 | 126 ± 22 | 98 ± 18 | 111 ± 34 | ||
| Africa | South Africa | 99 ± 12 | 2 ± 3 | 11 ± 14 | 65 ± 22 | 118 ± 26 | 25 ± 13 | 72 ± 30 | |
| North America | Hawaii | 0 | 0 | 0 | 7 ± 6 | 26 ± 5 | 0 | 72 ± 28 | |
| Europe | Portugal | 0 | 0 | 174 ± 84 | 6 ± 10 | 19 ± 14 | 0 | 22 ± 12 | |
| Germany | 0 | 0 | 24 ± 18 | 0 | 0 | 0 | 19 ± 26 | ||
| Ireland | 77 ± 77 | 25 ± 24 | 317 ± 78 | 32 ± 3 | 46 ± 66 | 84 ± 36 | 58 ± 12 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Swilaiman, S.S.; O’Gorman, C.M.; Du, W.; Sugui, J.A.; Del Buono, J.; Brock, M.; Kwon-Chung, K.J.; Szakacs, G.; Dyer, P.S. Global Sexual Fertility in the Opportunistic Pathogen Aspergillus fumigatus and Identification of New Supermater Strains. J. Fungi 2020, 6, 258. https://doi.org/10.3390/jof6040258
Swilaiman SS, O’Gorman CM, Du W, Sugui JA, Del Buono J, Brock M, Kwon-Chung KJ, Szakacs G, Dyer PS. Global Sexual Fertility in the Opportunistic Pathogen Aspergillus fumigatus and Identification of New Supermater Strains. Journal of Fungi. 2020; 6(4):258. https://doi.org/10.3390/jof6040258
Chicago/Turabian StyleSwilaiman, Sameira S., Céline M. O’Gorman, Wenyue Du, Janyce A. Sugui, Joanne Del Buono, Matthias Brock, Kyung J. Kwon-Chung, George Szakacs, and Paul S. Dyer. 2020. "Global Sexual Fertility in the Opportunistic Pathogen Aspergillus fumigatus and Identification of New Supermater Strains" Journal of Fungi 6, no. 4: 258. https://doi.org/10.3390/jof6040258
APA StyleSwilaiman, S. S., O’Gorman, C. M., Du, W., Sugui, J. A., Del Buono, J., Brock, M., Kwon-Chung, K. J., Szakacs, G., & Dyer, P. S. (2020). Global Sexual Fertility in the Opportunistic Pathogen Aspergillus fumigatus and Identification of New Supermater Strains. Journal of Fungi, 6(4), 258. https://doi.org/10.3390/jof6040258