Metabolites from Clonostachys Fungi and Their Biological Activities





Abstract
1. Introduction
2. Nitrogen-Containing Metabolites and Their Biological Activities
2.1. Linear Oligopeptides
2.2. Cyclopeptides
2.3. Piperazines
2.4. Other Nitrogen-Containing Metabolites
3. Polyketides and Their Biological Activities
3.1. Pyranones
3.2. Quinones
3.3. Sorbicillinoids
3.4. Other Polyketides
4. Terpenoids and Their Biological Activities
4.1. Monoterpenoids
4.2. Sesquiterpenoids
4.3. Diterpenoids
4.4. Triterpenoids
4.5. Polyterpenoids
4.6. Meroterpenoids
5. Miscellaneous Metabolites and Their Biological Activities
6. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Schroers, H.-J. A monograph of Bionectria (Ascomycota, Hypocreales, Bionectriaceae) and its Clonostachys anamorphs. Stud. Mycol. 2001, 46, 1–214. [Google Scholar]
- Chikanishi, T.; Hasumi, K.; Harada, T.; Kawasaki, N.; Endo, A. Clonostachin, a novel peptaibol that inhibits platelet aggregation. J. Antibiot. 1997, 50, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, M.; Raja, H.; Falkinham, J.O.; Adcock, A.F.; Kroll, D.J.; Wani, M.C.; Pearce, C.J.; Oberlies, N.H. Peptaibols, tetramic acid derivatives, isocoumarins, and sesquiterpenes from a Bionectria sp. (MSX 47401). J. Nat. Prod. 2013, 76, 1007–1015. [Google Scholar] [CrossRef] [PubMed]
- Ebrahim, W.; Kjer, J.; Amrani, M.E.; Wray, V.; Lin, W.; Ebel, R.; Lai, D.; Proksch, P. Pullularins E and F, two new peptides from the endophytic fungus Bionectria ochroleuca isolated from the mangrove plant Sonneratia caseolaris. Mar. Drugs 2012, 10, 1081–1091. [Google Scholar] [CrossRef] [PubMed]
- Arai, N.; Shiomi, K.; Yamaguchi, Y.; Masuma, R.; Iwai, Y.; Turberg, A.; Kolbl, H.; Omura, S. Argadin, a new chitinase inhibitor, produced by Clonostachys sp. FO7314. Chem. Pharm. Bull. 2000, 48, 1442–1446. [Google Scholar] [CrossRef]
- Arai, N.; Shiomi, K.; Iwai, Y.; Omura, S. Argifin, a new chitinase inhibitor, produced by Gliocladium sp. FTD-0668. J. Antibiot. 2000, 53, 609–614. [Google Scholar] [CrossRef]
- Shiomi, K.; Arai, N.; Iwai, Y.; Turberg, A.; Kolbl, H.; Omura, S. Structure of arifin, a new chitinase inhibitor produced by Gliocladium sp. Tetrahedron Lett. 2000, 41, 2141–2143. [Google Scholar] [CrossRef]
- Bal-Tembe, S.; Kundu, S.; Roy, K.; Hiremath, C.P.; Gole, G.; De Souza, E.P.; Kumar, E.K.S.V.; Gates, D.A.; Pillmoor, J.B. Activity of the ilicicolins against plant pathogenic fungi. Pestic. Sci. 1999, 55, 645–647. [Google Scholar] [CrossRef]
- Helaly, S.E.; Ashrafi, S.; Teponno, R.B.; Bernecker, S.; Dababat, A.A.; Maier, W.; Stadler, M. Nematicidal cyclic lipodepsipeptides and a xanthocillin derivative from a phaeosphariaceous fungus parasitizing eggs of the plant parasitic nematode Heterodera filipjevi. J. Nat. Prod. 2018, 81, 2228–2234. [Google Scholar] [CrossRef]
- Adachi, K.; Kanoh, K.; Wisespongp, P.; Nishijima, M.; Shizuri, Y. Clonostachysins A and B, new antidinoflagellate cyclic peptides from a marine-derived fungus. J. Antibiot. 2005, 58, 145. [Google Scholar] [CrossRef]
- Abdel-Wahab, N.M.; Harwoko, H.; Muller, W.E.G.; Hamacher, A.; Kassck, M.U.; Fouad, M.A.; Kamel, M.S.; Lin, W.; Ebrahim, W.; Liu, Z.; et al. Cyclic heptapeptides from the soil-derived fungus Clonostachys rosea. Bioorgan. Med. Chem. 2019, 27, 3954–3959. [Google Scholar] [CrossRef] [PubMed]
- Goto, T.; Kino, T.; Okuhara, M.; Tanaka, H.; Tsurumi, Y.; Takase, S. Cyclosporins A and C and Their Manufacture with Nectria. PCT International Application No. WO9213094, 6 August 1992. [Google Scholar]
- Wang, X.; Lin, M.; Xu, D.; Lai, D.; Zhou, L. Structural diversity and biological activities of fungal cyclic peptides, excluding cyclodipeptides. Molecules 2017, 22, 2069. [Google Scholar] [CrossRef] [PubMed]
- Cruz, L.J.; Insua, M.I.; Baz, J.P.; Trujillo, M.; Rodriguez-Mias, R.A.; Oliveira, E.; Giralt, E.; Albericio, F.; Canedo, L.M. IB-01212, a new cytotoxic cyclodepsipeptide isolated from the marine fungus Clonostachys sp. ESNA-A009. J. Org. Chem. 2006, 71, 3335–3338. [Google Scholar] [CrossRef]
- Luque-Ortega, J.R.; Cruz, L.J.; Albericio, F.; Rivas, L. The antitumoral depsipeptide IB-01212 kills leishmania through an apoptosis-like process involving intracellular targets. Mol. Pharm. 2010, 7, 1608–1617. [Google Scholar] [CrossRef]
- Zheng, C.-J.; Kim, C.-J.; Bae, K.S.; Kim, Y.-H.; Kim, W.-G. Bionectins A-C, epidithiodioxopiperazines with anti-MRSA activity, from Bionectria byssicola F120. J. Nat. Prod. 2006, 69, 1816–1819. [Google Scholar] [CrossRef]
- Yang, Y.-H.; Yang, D.-S.; Li, G.-H.; Pu, X.-J.; Mo, M.-H.; Zhao, P.-J. Antibacterial diketopiperazines from an endophytic fungus Bionectria sp. Y1085. J. Antibiot. 2019, 72, 752–758. [Google Scholar] [CrossRef]
- Watts, K.R.; Ratnam, J.; Ang, K.-H.; Tenney, K.; Compton, J.E.; McKerrow, J.; Crews, P. Assessing the trypanocidal potential of natural and semi-synthetic diketopiperazines from two deep water marine-derived fungi. Bioorgan. Med. Chem. 2010, 18, 2566–2574. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, T.; Watanabe, Y.; Nonaka, K.; Muramatsu, R.; Noguchi, C.; Tozawa, M.; Hokari, R.; Ishiyama, A.; Koike, R.; Matsui, H.; et al. Clonocoprogens A, B and C, new antimalarial coprogens from the Okinawan fungus Clonostachys compactiuscula FKR-0021. J. Antibiot. 2020, 73, 365–371. [Google Scholar] [CrossRef]
- Li, E.; Hou, B.; Gao, Q.; Xu, Y.; Zhang, C.; Liu, X.; Jiang, X.; Che, Y. Disulfide cleavage in dimeric epipolythiodioxopiperazine natural product diminishes its apoptosis-inducing effect but enhances autophagy in tumor cells. J. Nat. Prod. 2020, 83, 601–609. [Google Scholar] [CrossRef]
- Usami, Y.; Yamaguchi, J.; Numata, A. Gliocladins A-C and giloperazine; cytotoxic dioxo- or trioxopiperazine metabolitesfrom a Gliocladium sp. separated from a sea hare. Heterocycles 2004, 63, 1123–1129. [Google Scholar] [CrossRef]
- Song, H.C.; Shen, W.Y.; Dong, J.Y. Nematicidal metabolites from Gliocladium roseum YMF1.00133. Appl. Biochem. Microbiol. 2016, 52, 324–330. [Google Scholar] [CrossRef]
- Dong, J.-Y.; He, H.-P.; Shen, Y.-M.; Zhang, K.-Q. Nematicidal epipolysulfanyldioxopiperazines from Gliocladium roseum. J. Nat. Prod. 2005, 68, 1510–1513. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.Y.; Zhou, W.; Li, L.; Li, G.H.; Liu, Y.J.; Zhang, K.Q. A new epidithiodioxopiperazine metabolite isolated from Gliocladium roseum YMF1.00133. Chin. Chem. Lett. 2006, 17, 922–924. [Google Scholar]
- Zheng, C.-J.; Kim, Y.-H.; Kim, W.-G. Glioperazine B, as a new antimicrobial agent against Staphylococcus aureus, and glioperazine C: Two new dioxopiperazines from Bionectria byssicola. Biosci. Biotechnol. Biochem. 2007, 71, 1979–1983. [Google Scholar] [CrossRef]
- Suzuki, Y.; Takahashi, H.; Esumi, Y.; Arie, T.; Morita, T.; Koshino, H.; Uzawa, J.; Uramoto, M.; Yamaguchi, I. Haematocin, a new antifungal diketopiperazine produced by Nectria haematococca Berk. et Br. (880701a-1) causing nectria blight disease on ornamental plants. J. Antibiot. 2000, 53, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, M.; Graf, T.N.; Ayers, S.; Adcock, A.F.; Kroll, D.J.; Yang, J.; Swanson, S.M.; Munoz-Acuna, U.; De Blanco, E.J.C.; Agrawal, R.; et al. Cytotoxic epipolythiodioxopiperazine alkaloids from filamentous fungi of the Bionectriaceae. J. Antibiot. 2012, 65, 559–564. [Google Scholar] [CrossRef]
- Kamdem, R.S.T.; Wang, H.; Wafo, P.; Ebrahim, W.; Oezkaya, F.C.; Makhloufi, G.; Janiak, C.; Sureechatchaiyan, P.; Kassack, M.U.; Lin, W.; et al. Induction of new metabolites from the endophytic fungus Bionectria sp. through bacterial co-culture. Fitoterapia 2018, 124, 132–136. [Google Scholar] [CrossRef]
- Irvine, N.M.; Yerkes, C.N.; Graupner, P.R.; Roberts, R.E.; Hahn, D.R.; Pearce, C.; Gerwick, B.C. Synthesis and characterization of synthetic analogs of cinnacidin, a novel phytotoxin from Nectria sp. Pest. Manag. Sci. 2008, 64, 891–899. [Google Scholar] [CrossRef]
- Cota, B.B.; Tunes, L.G.; Maia, D.N.B.; Ramos, J.P.; Menezes de Oliveira, D.; Kohlhoff, M.; Maria de Alves, T.; Souza-Fagundes, E.M.; Campos, F.F.; Zani, C.L. Leishmanicidal compounds of Nectria pseudotrichia, an endophytic fungus isolated from the plant Caesalpinia echinata (Brazilwood). Mem. Inst. Oswaldo Cruz 2018, 113, 102–110. [Google Scholar] [CrossRef]
- Eilbert, F.; Thines, E.; Arendholz, W.R.; Sterner, O.; Anke, H. Fusarin C, (7Z)-fusarin C and (5Z)-fusarin C; inhibitors of dihydroxynaphthalene-melanin biosynthesis from Nectria coccinea (Cylindrocarpon sp.). J. Antibiot. 1997, 50, 443–445. [Google Scholar] [CrossRef]
- Wang, Y.; Ren, J.; Li, H.; Pan, Y.; Liu, X.; Che, Y.; Liu, G. The disruption of verM activates the production of gliocladiosin A and B in Clonostachys rogersoniana. Org. Biomol. Chem. 2019, 17, 6782–6785. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Ren, F.; Guo, Z.; Liu, J.; Liu, X.; Liu, G.; Che, Y. Rogersonins A and B, imidazolone N-oxide-incorporating indole alkaloids from a verG disruption mutant of Clonostachys rogersoniana. J. Nat. Prod. 2019, 82, 462–468. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, X.; Qian, F.; Zhu, T.; Xu, J.; Li, Y.; Zhang, L.; Jiao, B. Chlorinated phenolic sesquiterpenoids from the Arctic fungus Nectria sp. B-13. Biochem. Syst. Ecol. 2015, 59, 22–25. [Google Scholar] [CrossRef]
- Shibata, T.; Nakayama, O.; Tsurumi, Y.; Okuhara, M.; Terano, H.; Kohsaka, M. A new immunomodulator, FR-900483. J. Antibiot. 1988, 41, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Awakawa, T.; Kaji, T.; Wakimoto, T.; Abe, I. A heptaketide naphthaldehyde produced by a polyketide synthase from Nectria haematococca. Bioorgan. Med. Chem. Lett. 2012, 22, 4338–4340. [Google Scholar] [CrossRef] [PubMed]
- Parisot, D.; Devys, M.; Barbier, M. 5-Deoxybostrycoidin, a new metabolite produced by the fungus Nectria haematococca (Berk. and Br.) Wr. Z. Naturforsch. B 1989, 44, 1473–1474. [Google Scholar] [CrossRef]
- Fu, P.; Zhou, T.; Ren, F.; Zhu, S.; Zhang, Y.; Zhuang, W.; Che, Y. Pseudonectrins A-D, heptaketides from an endophytic fungus Nectria pseudotrichia. RSC Adv. 2019, 9, 12146–12152. [Google Scholar] [CrossRef]
- Wang, X.; Gong, X.; Li, P.; Lai, D.; Zhou, L. Structural diversity and biological activities of cyclic depsipeptides from fungi. Molecules 2018, 23, 169. [Google Scholar] [CrossRef]
- Isaka, M.; Berkaew, P.; Intereya, K.; Komwijit, S.; Sathitkunanon, T. Antiplasmodial and antiviral cyclohexadepsipeptides form the endophytic fungus Pullularia sp. BCC 8613. Tetrahedron 2007, 63, 6855–6860. [Google Scholar] [CrossRef]
- Wang, X.; Li, Y.; Zhang, X.; Lai, D.; Zhou, L. Structural diversity and biological activities of the cyclodipeptides from fungi. Molecules 2017, 22, 2026. [Google Scholar] [CrossRef]
- Salvi, A.; Amrine, C.S.M.; Austin, J.R.; Kilpatrick, K.; Russo, A.; Lantivit, D.; Calderon-Gierszal, E.; Mattes, Z.; Pearce, C.J.; Grinstaff, M.W.; et al. Verticillin A causes apoptosis and reduces tumor burden in high-grade serous ovarian cancer by inducing DNA damage. Mol. Cancer Ther. 2020, 19, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.M. The biosynthesis, molecular genetics and enzymology of the polyketide-derived metabolites. Nat. Prod. Rep. 2006, 23, 256–320. [Google Scholar] [CrossRef] [PubMed]
- Zhai, M.-M.; Qi, F.-M.; Li, J.; Jiang, C.-X.; Hou, Y.; Shi, Y.-P.; Di, D.-L.; Zhang, J.-W.; Wu, Q.-X. Isolation of secondary metabolites from the soil-derived fungus Clonostachys rosea YRS-06, a biological control agent, and evaluation of antibacterial activity. J. Agric. Food Chem. 2016, 64, 2298–2306. [Google Scholar] [CrossRef] [PubMed]
- Carey, S.T.; Nair, M.S.R. Metabolites of pyrenomycetes. V. identification of an antibiotic from two species of Nectria, as cephalochromin. Lloydia 1975, 38, 448–449. [Google Scholar]
- Hsiao, C.-J.; Hsiao, G.; Chen, W.-L.; Wang, S.-W.; Chiang, C.-P.; Liu, L.-Y.; Guh, J.-H.; Lee, T.-H.; Chung, C.-L. Cephalochromin induces G0/G1 cell cycle arrest and apoptosis in A549 human non-small-cell lung cancer cells by inflicting mitochondrial disruption. J. Nat. Prod. 2014, 77, 758–765. [Google Scholar] [CrossRef]
- Cui, H.; Liu, Y.Y.; Nie, Y.; Liu, Z.M.; Chen, S.H.; Zhang, Z.R.; Lu, Y.J.; He, L.; Huang, X.S.; She, Z.G. Polyketides from the mangrove-derived endophytic fungus Nectria sp. HN001 and their α-glucosidase inhibitory activity. Mar. Drugs 2016, 14, 86. [Google Scholar] [CrossRef]
- Meng, L.-H.; Chen, H.-Q.; Form, I.; Konuklugil, B.; Proksch, P.; Wang, B.-G. New chromone, isocoumarin, and indole alkaloid derivatives from three sponge-derived fungal strains. Nat. Prod. Commun. 2016, 11, 1293–1296. [Google Scholar] [CrossRef] [PubMed]
- Ayer, W.A.; Shewchuk, L.M. Metabolites of Nectria fuckeliana. J. Nat. Prod. 1986, 49, 847–848. [Google Scholar] [CrossRef]
- Ariefta, N.R.; Kristiana, P.; Aboshi, T.; Murayama, T.; Tawaraya, K.; Koseki, T.; Kurisawa, N.; Kimura, K.; Shiono, Y. New isocoumarins, naphthoquinones, and a cleistanthane-type diterpene from Nectria pseudotrichia 120-1NP. Fitoterapia 2018, 127, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, X.-B.; Li, L.; Li, R.-J.; Lou, H.-X. α-Pyrone derivatives from the endolichenic fungus Nectria sp. Phytochem. Lett. 2015, 12, 22–26. [Google Scholar] [CrossRef]
- Parisot, D.; Devys, M.; Ferezou, J.P.; Barbier, M. Pigments from Nectria haematococca: Anhydrofusarubin lactone and nectriafurone. Phytochemistry 1983, 22, 1301–1303. [Google Scholar] [CrossRef]
- Ayers, S.; Zink, D.L.; Mohn, K.; Powell, J.S.; Brown, C.M.; Bills, G.; Grund, A.; Thompson, D.; Singh, S.B. Anthelmintic constituents of Clonostachys candelabrum. J. Antibiot. 2010, 63, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Nair, M.S.; Anchel, M. Antibacterial quinone-hydroquinone pair from the ascomycete, Nectria coryli. Tetrahedron Lett. 1972, 13, 795–796. [Google Scholar] [CrossRef]
- Parisot, D.; Devys, M.; Barbier, M. A new deoxyfusarubin produced by the fungus Nectria haematococca. Synthesis of the two isomeric deoxyanhydronaphthopyranones from toralactone. J. Antibiot. 1992, 45, 1799–1801. [Google Scholar] [CrossRef]
- Parisot, D.; Devys, M.; Barbier, M. Fusarubinoic acid, a new naphthoquinone from the fungus Nectria haematococca. Phytochemistry 1988, 27, 3002–3004. [Google Scholar] [CrossRef]
- Carey, S.T.; Nair, M.S.R. Metabolites of Pyrenomycetes. X. Isolation of p-toluquinone and toluquinol from Nectria erubescens. J. Nat. Prod. 1979, 42, 231. [Google Scholar] [CrossRef]
- Supratman, U.; Suzuki, T.; Nakamura, T.; Yokoyama, Y.; Harneti, D.; Maharani, R.; Salam, S.; Abdullah, F.; Koseki, T.; Shiono, Y. New metabolites produced by endophyte Clonostachys rosea B5−2. Nat. Prod. Res. 2019. [Google Scholar] [CrossRef] [PubMed]
- Grabley, S.; Hammann, P.; Thiericke, R.; Wink, J.; Philipps, S.; Zeeck, A. Secondary metabolites by chemical screening. 21. Clonostachydiol, a novel anthelmintic macrodiolide from the fungus Clonostachys cylindrospora (strain FH-A 6607). J. Antibiot. 1993, 46, 343–345. [Google Scholar] [CrossRef]
- Freinkman, E.; Oh, D.-C.; Scott, J.J.; Currie, C.R.; Clardy, C. Bionectriol A, a polyketide glycoside from the fungus Bionectria sp. associated with the fungus-growing ant, Apterostigma dentigerum. Tetrahedron Lett. 2009, 50, 6834–6837. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; You, J.; King, J.B.; Cai, S.; Park, E.; Powell, D.R.; Cichewicz, R.H. Polyketide glycosides from Bionectria ochroleuca inhibit Candida albicans biofilm formation. J. Nat. Prod. 2014, 77, 2273–2279. [Google Scholar] [CrossRef] [PubMed]
- Yin, T.-P.; Xing, Y.; Cai, L.; Yu, J.; Luo, P.; Ding, Z.-T. A new polyketide glycoside from the rhizospheric Clonostachys rogersoniana associated with Panax notoginseng. J. Asian Nat. Prod. Res. 2017, 19, 1258–1263. [Google Scholar] [CrossRef] [PubMed]
- Gutiearrez, M.; Theoduloz, C.; Rodriaguez, J.; Lolas, M.; Schmeda-Hirschmann, G. Bioactive metabolites from the fungus Nectria galligena, the main apple canker agent in chile. J. Agric. Food Chem. 2005, 53, 7701–7708. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.-B.; Jiao, H.; Zhu, Y.-P.; Zhang, J.-P.; Lu, X.-L.; Liu, X.-Y. Bioactive metabolites from the Arctic fungus Nectria sp. B-13. J. Asian Nat. Prod. Res. 2019, 21, 961–969. [Google Scholar] [CrossRef]
- Ju, Y.M.; Juang, S.H.; Chen, K.J.; Lee, T.H. TMC-151 A monoacetate, a new polyketide from Bionectria ochroleuca. Z. Naturforsch. B 2007, 62, 561–564. [Google Scholar] [CrossRef]
- Kohno, J.; Nishio, M.; Sakurai, M.; Kawano, K.; Hiramatsu, H.; Kameda, N.; Kishi, N.; Yamashita, T.; Okuda, T.; Komatsubara, S. Isolation and structure determination of TMC-151s: Novel polyketide antibiotics from Gliocladium catenulatum Gilman & Abbott TC 1280. Tetrahedron 1999, 55, 7771–7786. [Google Scholar]
- Okuda, T.; Kohno, J.; Kishi, N.; Asai, Y.; Nishio, M.; Komatsubara, S. Production of TMC-151, TMC-154 and TMC-171, a new class of antibiotics, is specific to ‘Gliocladium roseum’ group. Mycoscience 2000, 41, 239–253. [Google Scholar] [CrossRef]
- Liu, Q.; Jiang, D.; Qi, Y.; Chen, C.; Xie, X.; Sun, L. Isolation, identification and activity analysis of antimicrobial compound from Bionectria ochroleuca strain Bo-1. Acta Phytophylacica Sin. 2014, 41, 41–44. [Google Scholar]
- Mao, Z.; Sun, W.; Fu, L.; Luo, H.; Lai, D.; Zhou, L. Natural dibenzo-α-pyrones and their bioactivities. Molecules 2014, 19, 5088–5108. [Google Scholar] [CrossRef]
- Meng, J.; Wang, X.; Xu, D.; Fu, X.; Zhang, X.; Lai, D.; Zhou, L.; Zhang, G. Sorbicillinoids from fungi and their bioactivities. Molecules 2016, 21, 715. [Google Scholar] [CrossRef]
- Ojima, K.; Yangchu, A.; Laksanacharoen, P.; Tasanathai, K.; Thanakitpipattana, D.; Tokuyama, H.; Isaka, M. Cordybislactone, a stereoisomer of the 14-membered bislactone clonostachydiol, form the hopper pathogenic fungus Cordyceps sp. BCC 49294: Revision of he absolute configuration of clonostachydiol. J. Antibiot. 2018, 71, 351–358. [Google Scholar] [CrossRef]
- Gong, T.; Zhen, X.; Li, B.-J.; Yang, J.-L.; Zhu, P. Two new monoterpenoid α-pyrones from a fungus Nectria sp. HLS206 associated with the marine sponge Gelliodes carnosa. J. Asian Nat. Prod. Res. 2015, 17, 633–637. [Google Scholar] [CrossRef]
- Ariefta, N.R.; Kristiana, P.; Nurjanto, H.H.; Momma, H.; Kwon, E.; Ashitani, T.; Tawaraya, K.; Murayama, T.; Koseki, T.; Furuno, H.; et al. Nectrianolins A, B, and C, new metabolites produced by endophytic fungus Nectria pseudotrichia 120-1NP. Tetrahedron Lett. 2017, 58, 4082–4086. [Google Scholar] [CrossRef]
- Dias, A.C.D.S.; Couzinet-Mossion, A.; Ruiz, N.; Lakhdar, F.; Etahiri, S.; Bertrand, S.; Ory, L.; Rousakis, C.; Pouchus, Y.F.; Nazih, E.-H.; et al. Steroids from marine-derived fungi: Evaluation of antiproliferative and antimicrobial activitie of eburicol. Mar. Drugs 2019, 17, 372. [Google Scholar] [CrossRef]
- Zhao, J.; Mou, Y.; Shan, T.; Li, Y.; Zhou, L.; Wang, M.; Wang, J. Antimicrobial metabolites from the endophytic fungus Pichia guilliermondii isolated from Paris polyphylla var. yunnanensis. Molecules 2010, 15, 7961–7970. [Google Scholar] [CrossRef]
- Kong, F.-D.; Huang, X.-L.; Ma, Q.-Y.; Xie, Q.-Y.; Wang, P.; Chen, P.-W.; Zhou, L.-M.; Yuan, J.-Z.; Dai, H.-F.; Luo, D.-Q. Helvolic acid derivatives with antibacterial activities against Streptococcus agalactiae from the marine-derived fungus Aspergillus fumigatus HNMF0047. J. Nat. Prod. 2018, 81, 1869–1876. [Google Scholar] [CrossRef] [PubMed]
- Nishida, H.; Huang, X.-H.; Tomoda, H.; Omura, S. Glisoprenins, new inhibitors of acyl-CoA: Cholesterol acyltransferase produced by Gliocladium sp. FO-1513. II. Structure elucidation of glisoprenins A and B. J. Antibiot. 1992, 45, 1669–1676. [Google Scholar] [CrossRef][Green Version]
- Sterner, O.; Thines, E.; Eilbert, F.; Anke, H. Glisoprenins C, D and E, new inhibitors of appressorium formation in Magnaporthe grisea, from cultures of Gliocladium roseum. 2. Structure determination. J. Antibiot. 1998, 51, 228–231. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Carey, S.T.; Nair, M.S.R. Metabolites from Pyrenomycetes. VIII. Identification of three metabolites from Nectria lucida as antibiotic triprenyl phenols. Lloydia 1977, 40, 602–603. [Google Scholar]
- Aldridge, D.C.; Borrow, A.; Foster, R.G.; Large, M.S.; Spencer, H.; Turner, W.B. Metabolites of Nectria coccinea. J. Chem. Soc. Perkin Trans. 1 1972, 17, 2136–2141. [Google Scholar] [CrossRef]
- Wang, R.-S.; Gong, T.; Zhu, P.; Cheng, K.-D. Isolation of metabolic products from the fungus Nectria sp. HLS206 that is associated with the marine sponge Gelliodes carnosa collected from the South China Sea. J. Chin. Pharm. Sci. 2012, 21, 183–186. [Google Scholar] [CrossRef]
- Kawahara, T.; Itoh, M.; Izumikawa, M.; Sakata, N.; Tsuchida, T.; Shin-ya, K. Cytotoxic sesquiterpenoids MBJ-0009 and MBJ-0010 from a saprobic fungus Nectria sp. f26111. J. Antibiot. 2013, 66, 567–569. [Google Scholar] [CrossRef] [PubMed]
- Sreekanth, D.; Sushim, G.K.; Syed, A.; Khan, B.M.; Ahmad, A. Molecular and morphological characterization of a taxol-producing endophytic fungus, Gliocladium sp., from Taxus baccata. Mycobiology 2011, 39, 151–157. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fex, T. Chalmicrin, a mannitol ether of methylated monocyclofarnesol, from Chalara microspora. Phytochemistry 1982, 21, 367–369. [Google Scholar] [CrossRef]
- Soliman, S.S.M.; Raizada, M.N. Sites of biosynthesis and storage of taxol in Taxus media (Rehder) plants: Mechanism of accumulation. Phytochemistry 2020, 175, 112369. [Google Scholar] [CrossRef] [PubMed]
- Flores-Bustamante, Z.R.; Rivera-Orduna, F.N.; Martinez-Cardenas, A.; Flores-Cotera, L.B. Microbial paclitaxel: Advances and perspectives. J. Antibiot. 2010, 63, 460–467. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Meng, Q.; Wang, M.; Yuan, J.; Shi, Z.; Zhang, Y.; Fu, S. Study on phenolic compounds of secondary metabolites from the fungus Gliocladium roseum CGMCC.3.3657. J. Zunyi Med. Univ. 2018, 41, 674–677. [Google Scholar]























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite Class | Metabolite Name | Fungal Species | Biological Activity | Ref. |
|---|---|---|---|---|
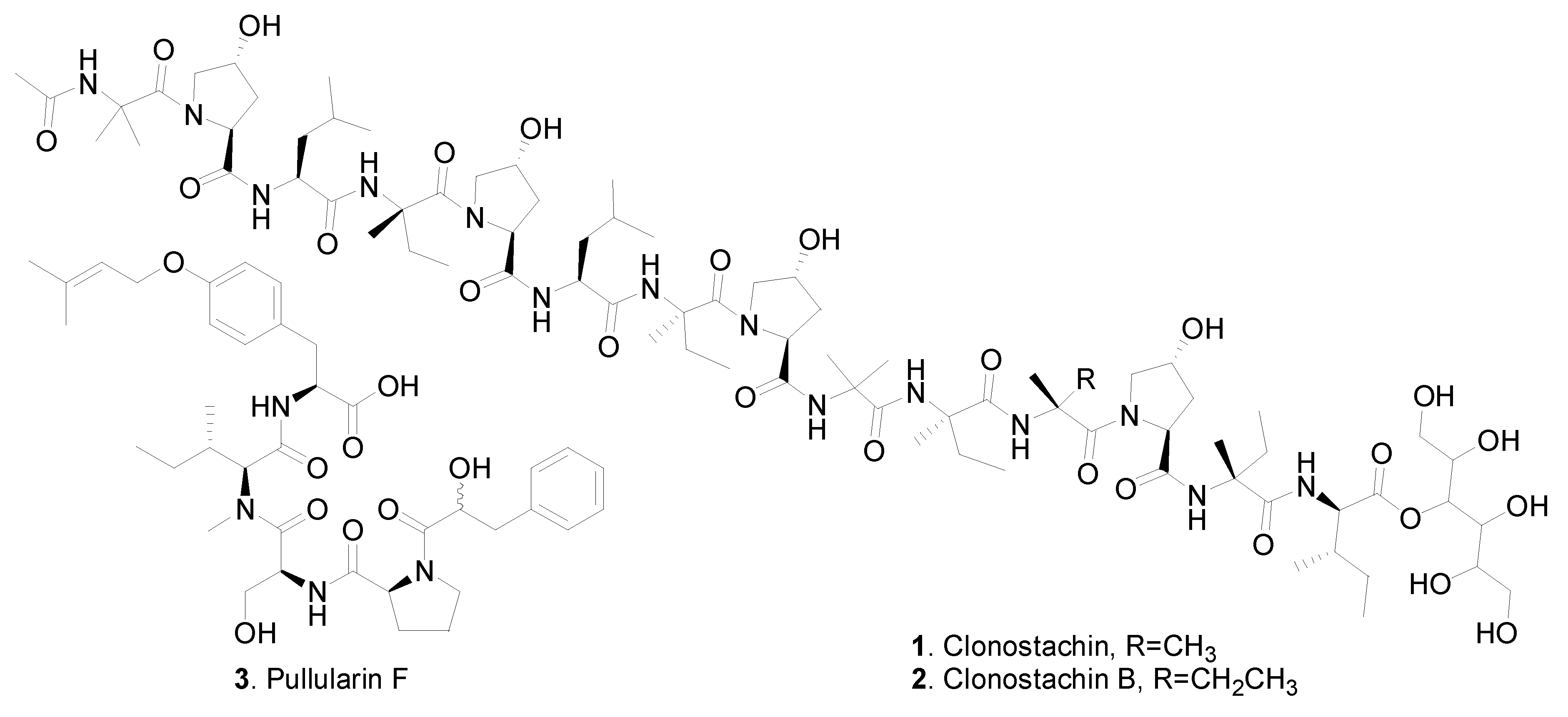
| Linear oligopeptides | Clonostachin (1) | Clonostachys sp. F5898 | Inhibition on platelet aggregation | [2] |
| Bionectria sp. MSX 47401 | - | [3] | ||
| Clonostachin B (2) | Bionectria sp. MSX 47401 | - | [3] | |
| Pullularin F (3) | Bionectria ochroleuca | - | [4] | |
| Cyclopeptides | Argadin (4) | Clonostachys sp. FO-7314 | Inhibitory activity on chitinase | [5] |
| Argifin (5) | Gliocladium sp. FTD-0668 | Inhibitory activity on chitinase | [6,7] | |
| Arthrichitin (6) | Nectria sp. | - | [8] | |
| Antifungal activity on the yeasts | [9] | |||
| Clonostachysin A (7) | Clonostachys rogersoniana | Anti-dinoflagellate activity | [10] | |
| Clonostachysin B (8) | Clonostachys rogersoniana | Anti-dinoflagellate activity | [10] | |
| Cyclo-(Gly-D-Leu-D-allo-Ile-L-Val-L-Val-D-Trp-β-Ala) (9) | Clonostachys rosea | Cytotoxic activity | [11] | |
| Cyclo-(Gly-D-Leu-L-Val-L-Val-L-Val-D-Trp-β-Ala) (10) | Clonostachys rosea | - | [11] | |
| Cyclo-(Gly-D-Leu-D-allo-Ile-D-allo-Ile-L-Val-D-Trp-β-Ala) (11) | Clonostachys rosea | - | [11] | |
| Cyclosporin A (12) | Nectria sp. F-4908 | - | [12] | |
| Immunosuppressive and antifungal activities | [13] | |||
| Cyclosporin C (13) | Nectria sp. F-4908 | - | [12] | |
| Immunosuppressive and antifungal activities | [13] | |||
| IB-01212 (14) | Clonostachys sp. ESNA-A009 | Cytotoxic activity | [14] | |
| Antileishmanial activity | [15] | |||
| Pullularin A (15) | Bionectria ochroleuca | Cytotoxic activity | [4] | |
| Pullularin C (16) | Bionectria ochroleuca | Cytotoxic activity | [4] | |
| Pullularin E (17) | Bionectria ochroleuca | - | [4] | |
| Piperazines | Bionectin A (18) | Bionectria byssicola F120 | Antibacterial activity | [16] |
| Bionectin B (19) | Bionectria byssicola F120 | Antibacterial activity | [16] | |
| Bionectin C (20) | Bionectria byssicola F120 | - | [16] | |
| Bionectin D (21) | Bionectria sp. Y1085 | Antibacterial activity | [17] | |
| Bionectin E (22) | Bionectria sp. Y1085 | Antibacterial activity | [17] | |
| 3,6-Bis(methylthio)-cyclo(alanyltryptophyl) (23) | Nectria inventa | Trypanocidal activity | [18] | |
| Chaetocin (24) | Nectria inventa | Trypanocidal activity | [18] | |
| Chetoseminudin B (25) | Nectria inventa | Trypanocidal activity | [18] | |
| Clonocoprogen A (26) | Clonostachys compactiuscula FKR-0021 | Antimalarial activity | [19] | |
| Clonocoprogen B (27) | Clonostachys compactiuscula FKR-0021 | Antimalarial activity | [19] | |
| Clonocoprogen C (28) | Clonostachys compactiuscula FKR-0021 | Antimalarial activity | [19] | |
| N14-Plmitoylcoprogen (29) | Clonostachys compactiuscula FKR-0021 | Antimalarial activity | [19] | |
| Cyclo (L-Pro-L-Leu) (30) | Bionectria sp. Y1085 | - | [17] | |
| Dioxopiperazine (31) | Bionectria sp. Y1085 | - | [17] | |
| Gliocladicillin A (32) | Bionectria sp. Y1085 | - | [17] | |
| Gliocladicillin C (33) | Bionectria sp. Y1085 | Antibacterial activity | [17] | |
| Clonostachys rogersoniana | Cytotoxic activity | [20] | ||
| Gliocladin A (34) | Glicladium roseum OUPS-N132 | - | [21] | |
| Gliocladin B (35) | Glicladium roseum OUPS-N132 | - | [21] | |
| Gliocladin C (36) | Glicladium roseum OUPS-N132 | Cytotoxic activity | [21] | |
| Gliocladium roseum YMF1.00133 | Antinematodal activity | [22] | ||
| Gliocladine A (37) | Gliocladium roseum 1A | Antinematodal activity | [23] | |
| Gliocladine B (38) | Gliocladium roseum YMF1.00133 | Antinematodal activity | [23] | |
| Bionectria sp. Y1085 | - | [17] | ||
| Gliocladine C (39) | Gliocladium roseum 1A | Antinematodal activity | [23] | |
| Gliocladine D (40) | Gliocladium roseum 1A | Antinematodal activity | [23] | |
| Gliocladine E (41) | Gliocladium roseum 1A | Antinematodal activity | [23] | |
| Glioclatine (42) | Gliocladium roseum YMF1.00133 | Antinematodal activity | [24] | |
| Glioperazine (43) | Gliocladium sp. OUPS-N132 | - | [21] | |
| Bionectria byssicola F120 | - | [25] | ||
| Clonostachys rosea | - | [11] | ||
| Glioperazine B (44) | Bionectria byssicola F120 | Antimicrobial activity | [25] | |
| Glioperazine C (45) | Bionectria byssicola F120 | - | [25] | |
| Haematocin (46) | Nectria haematococca | Antifuangl activity | [26] | |
| Lasiodipline D (47) | Bionectria sp. Y1085 | - | [17] | |
| Sch52900 (48) | Gliocladium roseum 1A | Antinematodal activity | [23] | |
| Sch52901 (49) | Gliocladium roseum 1A | Antinematodal activity | [23] | |
| Verticillin A (50) | Gliocladium roseum 1A | Antinematodal activity | [23] | |
| Bionectria sp. Y1085 | - | [17] | ||
| 11′-Deoxyverticillin A (51) | Gliocladium roseum 1A | Antinematodal activity | [23] | |
| 11,11′-Dideoxyverticillin A (52) | Bionectria sp. Y1085 | - | [17] | |
| Verticillin B (53) | Nectria inventa | Trypanocidal activity | [18] | |
| Verticillin D (54) | Bionectria byssicola F120 | - | [16] | |
| Bionectria ochroleuca | Cytotoxic activity | [4] | ||
| Clonostachys rosea | Cytotoxic activity | [11] | ||
| Verticillin G (55) | Bionectria byssicola F120 | Antibacterial activity | [25] | |
| Verticillin H (56) | Bionectriaceous strains MSX 64546 and MSX 59553 | Cytotoxic activity | [27] | |
| Other nitrogen-containing metabolites | N-Benzyl-3-phenyllactamide (57) | Clonostachys compactiuscula FKR-0021 | - | [19] |
| N-Benzyl-3-phenylpropanamide (58) | Clonostachys compactiuscula FKR-0021 | - | [19] | |
| Bionectriamine A (59) | Co-cultivation of Bionectria sp. with Bacillus subtilis or Streptomyceslividans | - | [28] | |
| Bionectriamine B (60) | Co-cultivation of Bionectria sp. with Bacillus subtilis or Streptomyceslividans | - | [28] | |
| Cinnacidin (61) | Nectria sp. DA060097 | Phytotoxic activity | [29] | |
| Clonostalactam (62) | Clonostachys rosea | - | [11] | |
| Cytochalasin D (63) | Nectria pseudotrichia | Weak leishmanicidal activity | [30] | |
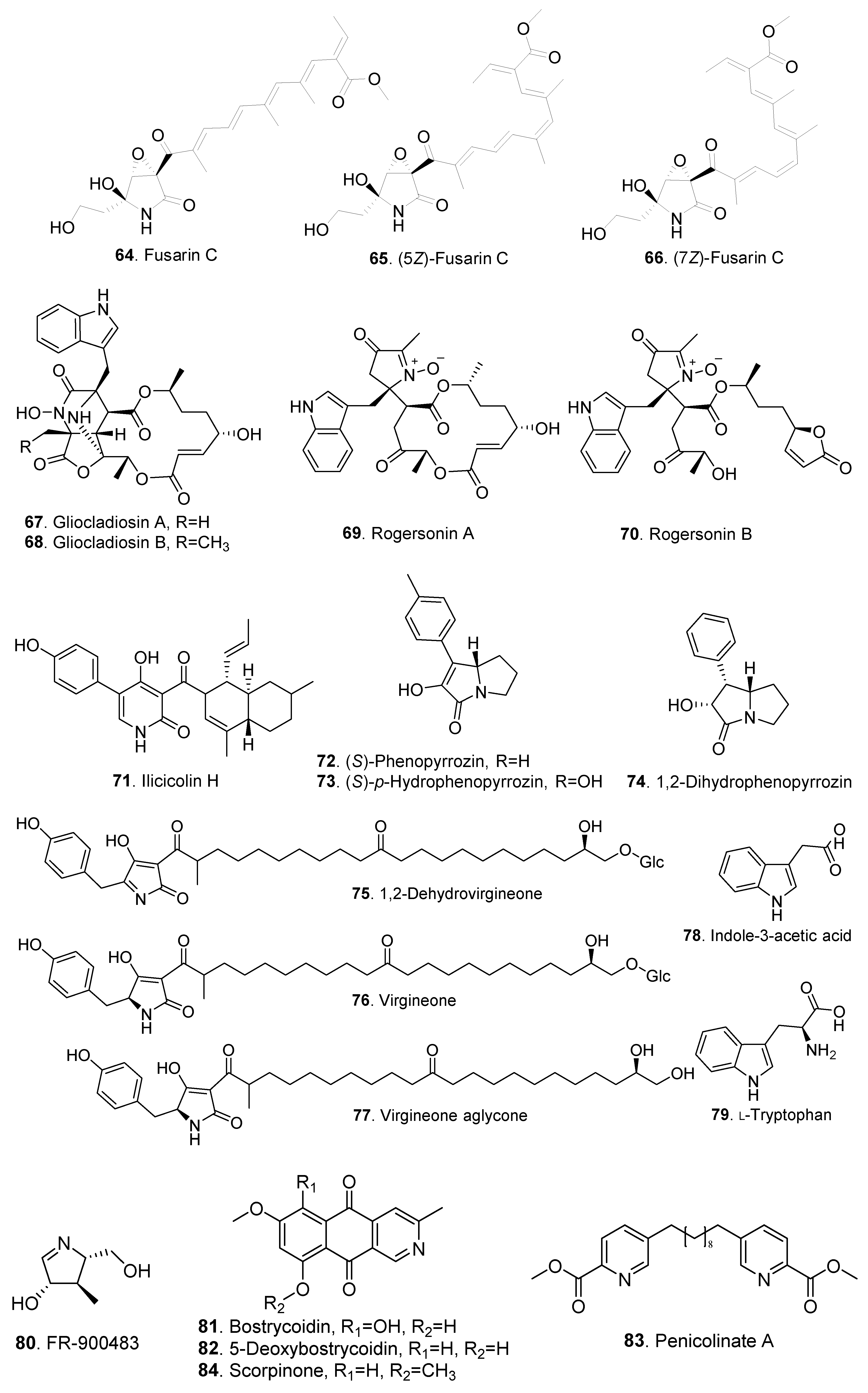
| Fusarin C (64) | Nectria coccinea A56-9 | Antifungal activity | [31] | |
| (5Z)-Fusarin C (65) | Nectria coccinea A56-9 | Antifungal activity | [31] | |
| (7Z)-Fusarin C (66) | Nectria coccinea A56-9 | Antifungal activity | [31] | |
| Gliocladiosin A (67) | verM disruption mutant of Clonostachys rogersoniana | Antibacterial activity | [32] | |
| Gliocladiosin B (68) | verM disruption mutant of Clonostachys rogersoniana | Antibacterial activity | [32] | |
| Rogersonin A (69) | verG disruption mutant of Clonostachys rogersoniana | - | [33] | |
| Rogersonin B (70) | verG disruption mutant of Clonostachys rogersoniana | - | [33] | |
| Ilicicolin H (71) | Nectria sp. B-13 | - | [34] | |
| (S)-Phenopyrrozin (72) | Bionectria sp. | - | [28] | |
| (S)-p-Hydroxyphenopyrrozin (73) | Bionectria sp. | - | [28] | |
| 1,2-Dihydrophenopyrrozin (74) | Bionectria sp. | - | [28] | |
| 1,2-Dehydrovirgineone (75) | Bionectria sp. MSX 47401 | Antibacterial activity | [3] | |
| Virgineone (76) | Bionectria sp. MSX 47401 | Antibacterial and antifungal activities | [3] | |
| Virgineone aglycone (77) | Bionectria sp. MSX 47401 | Antibacterial activity | [3] | |
| Indole-3-acetic acid (78) | Bionectria sp. Y1085 | - | [17] | |
| L-Tryptophan (79) | Bionectria sp. Y1085 | - | [17] | |
| FR-900483 (80) | Nctria lucida F-4490 | Immunostimulatory activity | [35] | |
| Bostrycoidin (81) | Nectria haematococca | - | [36] | |
| 5-Deoxybostrycoidin (82) | Nectria haematococca | - | [37] | |
| Penicolinate A (83) | Bionectria sp. | Cytotoxic activity | [28] | |
| Scorpinone (84) | Nectria pseudotrichia | - | [38] |
| Metabolite Class | Metabolite Name | Fungal Species | Biological Activity | Ref. |
|---|---|---|---|---|
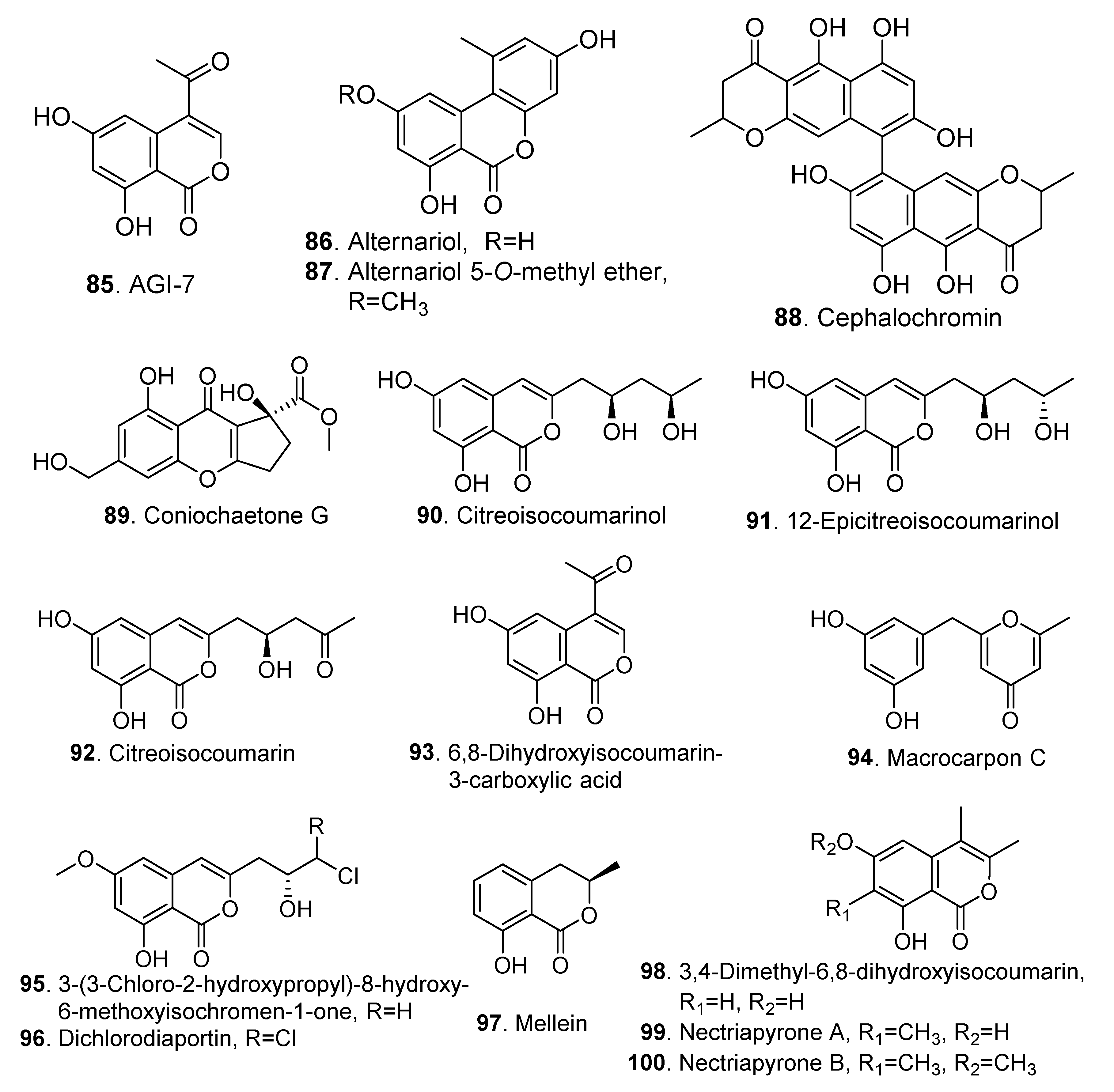
| Pyranones | AGI-7 (85) | Bionectria sp. MSX 47401 | - | [3] |
| Alternariol (86) | Clonostachys rosea YRS-06 | - | [44] | |
| Alternariol 5-O-methyl ether (87) | Clonostachys rosea YRS-06 | - | [44] | |
| Cephalochromin (88) | Nectria viridescens | - | [45] | |
| Cytotoxic activity | [46] | |||
| Coniochaetone G (89) | Clonostachys compactiuscula FKR-0021 | - | [19] | |
| Citreoisocoumarinol (90) | Nectria sp. HN001 | Inhibitory activity on α-glucosidase | [47] | |
| 12-Epicitreoisocoumarinol (91) | Nectria sp. HN001 | - | [47] | |
| Citreoisocoumarin (92) | Nectria sp. HN001 | Inhibitory activity on α-glucosidase | [47] | |
| 6,8-Dihydroxyisocoumarin -3-carboxylic acid (93) | Co-cultivation of Bionectria sp. with Bacillus subtilis or Streptomyceslividans | - | [28] | |
| Macrocarpon C (94) | Nectria sp. HN001 | Inhibitory activity on α-glucosidase | [47] | |
| 3-(3-Chloro-2-hydroxypropyl)-8-hydroxy-6-methoxyisochromen-1-one (95) | Clonostachys sp. AP4.1 | - | [48] | |
| Dichlorodiaportin (96) | Clonostachys sp. AP4.1 | - | [48] | |
| Mellein (97) | Nectria fuckeliana | - | [49] | |
| 3,4-Dimethyl-6,8-dihydroxyisocoumarin (98) | Nectria pseudotrichia 120-1NP | - | [50] | |
| Nectriapyrone A (99) | Nectria pseudotrichia 120-1NP | - | [50] | |
| Nectriapyrone B (100) | Nectria pseudotrichia 120-1NP | - | [50] | |
| (S)-4-Methoxy-6-pentanoyl-5,6-dihydro-2H-pyran-2-one (101) | Nectria sp. | - | [51] | |
| Necpyrone A (102) | Nectria sp. | - | [51] | |
| Necpyrone B (103) | Nectria sp. | - | [51] | |
| Necpyrone C (104) | Nectria sp. | - | [51] | |
| Necpyrone D (105) | Nectria sp. | - | [51] | |
| Necpyrone E (106) | Nectria sp. | - | [51] | |
| LL-P880α (107) | Nectria sp. | - | [51] | |
| LL-P880β (108) | Nectria sp. | - | [51] | |
| (1S, 2R)-1-Hydroxy-1-((S)-4-methoxy-6-oxo-3,6-dihydro-2H-pyran-2-yl)-pentan-2-yl acetate (109) | Nectria sp. | - | [51] | |
| LL-P880γ (110) | Nectria sp. | - | [51] | |
| PC-2 (111) | Nectria sp. | - | [51] | |
| Quinones | Anhydrofusarubin lactone (112) | Nectria haematococca | - | [52] |
| Aurantiogliocladin (113) | Clonostachys candelabrum | - | [53] | |
| 2,5-Dimethyoxy-3,6-dimethyl-1,4-benzoquinone (114) | Nectria coryli | Antibacterial activity | [54] | |
| Nectria fuckeliana | - | [49] | ||
| Fusarubin (115) | Nectria haematococca | - | [36] | |
| 4-Deoxyfusarubin (116) | Nectria haematococca | - | [55] | |
| 4-Deoxyanhydrofusarubin (117) | Nectria haematococca | - | [55] | |
| 5-Deoxyfusarubin (118) | Nectria haematococca | - | [55] | |
| 5-Deoxyanhydrofusarubin (119) | Nectri haematococca | - | [55] | |
| Fusarubinoic acid (120) | Nectria haematococca | - | [56] | |
| 13-Hydroxynorjavanicin (121) | Nectria haematococca | - | [56] | |
| Herbarin (122) | Nectria pseudotrichia 120-1NP | Antibacterial and phytotoxic activities | [50] | |
| Nectria pseudotrichia | Cytotoxic activity | [38] | ||
| O-Methylherbarin (123) | Nectria pseudotrichia 120-1NP | Cytotoxic activity | [50] | |
| Dehydroherbarin (124) | Nectria pseudotrichia | Cytotoxi activity | [38] | |
| 2-Acetoxyl-5,7-dimethoxy-3-methyl-1,4-naphthoquinone (125) | Nectria pseudotrichia | Cytotoxic activity | [38] | |
| Pseudonectrin A (126) | Nectria pseudotrichia | Cytotoxic activity | [38] | |
| Pseudonectrin B (127) | Nectria pseudotrichia | Cytotoxic activity | [38] | |
| Pseudonectrin C (128) | Nectria pseudotrichia | Cytotoxic activity | [38] | |
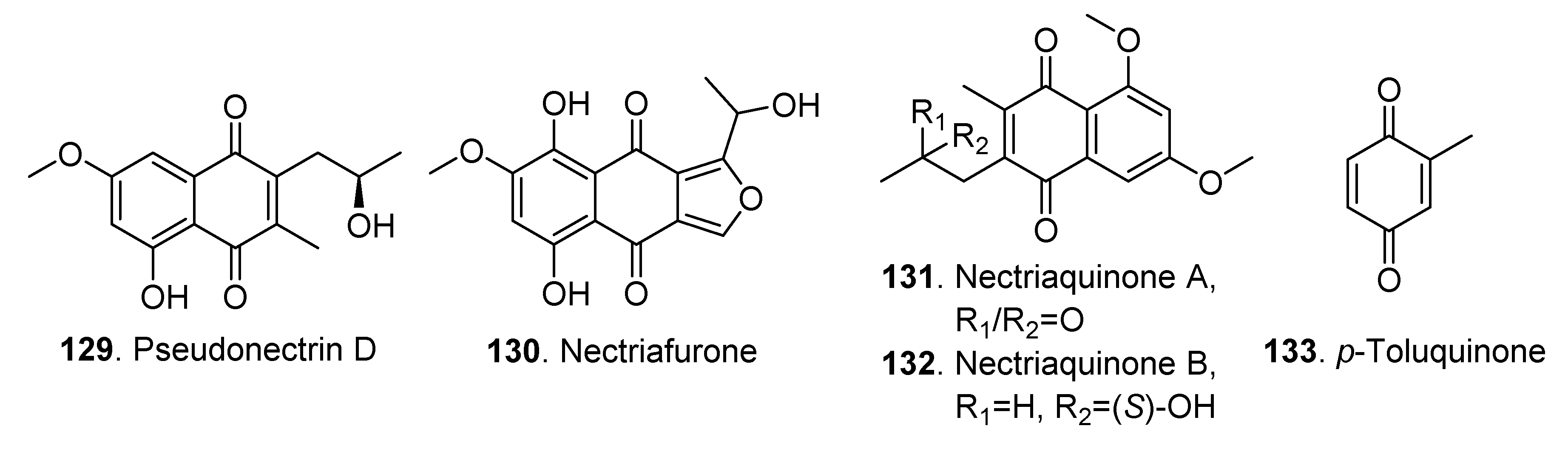
| Pseudonectrin D (129) | Nectria pseudotrichia | - | [38] | |
| Nectriafurone (130) | Nectria haematococca | - | [52] | |
| Nectriaquinone A (131) | Nectria pseudotrichia 120-1NP | Cytotoxic activity | [50] | |
| Nectriaquinone B (132) | Nectria pseudotrichia 120-1NP | Antibacterial and cytotoxic activities | [50] | |
| P-Toluquinone (133) | Nectria erubescens | - | [57] | |
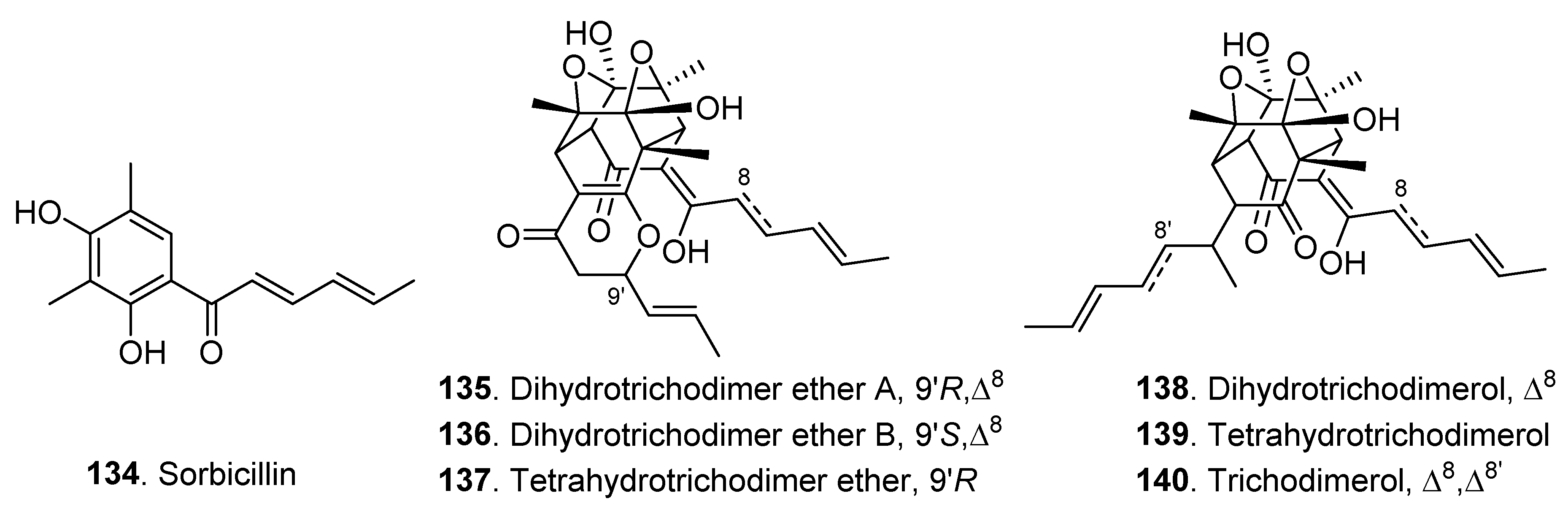
| Sorbicillinoids | Sorbicillin (134) | Clonostachys rosea YRS-06 | - | [44] |
| Dihydrotrichodimer ether A (135) | Clonostachys rosea YRS-06 | Antibacterial activity | [44] | |
| Dihydrotrichodimer ether B (136) | Clonostachys rosea YRS-06 | Antibacterial activity | [44] | |
| Tetrahydrotrichodimer ether (137) | Clonostachys rosea YRS-06 | - | [44] | |
| Dihydrotrichodimerol (138) | Clonostachys rosea YRS-06 | Antibacterial activity | [44] | |
| Tetrahydrotrichodimerol (139) | Clonostachys rosea YRS-06 | Antibacterial activity | [44] | |
| Trichodimerol (140) | Clonostachys rosea YRS-06 | - | [44] | |
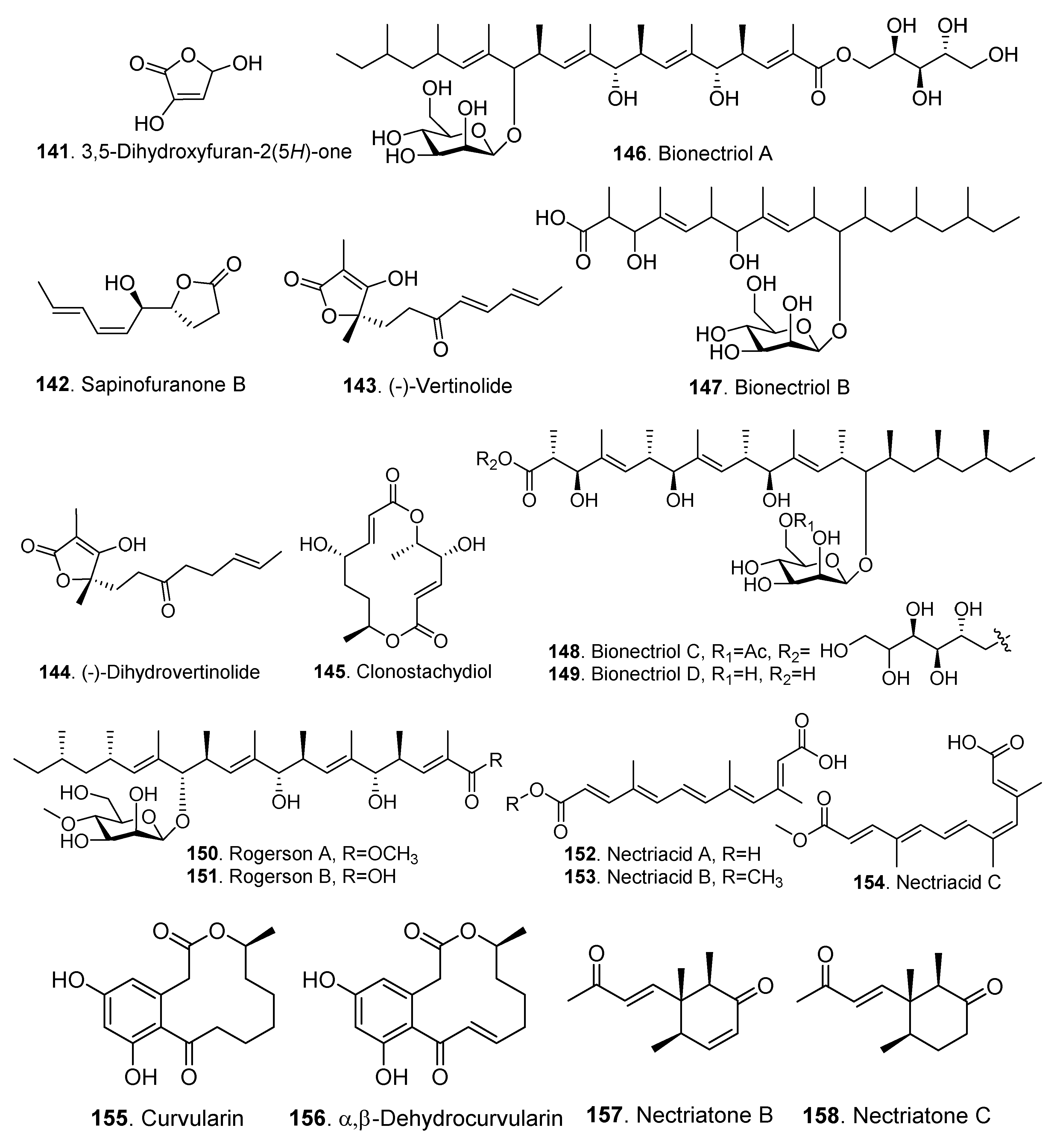
| Other polyketides | 3,5-Dihydroxyfuran-2(5H)-one (141) | Gliocladium roseum 1A | - | [23] |
| Clonostachys rosea | - | [11] | ||
| Sapinofuranone B (142) | Gliocladium roseum 1A | - | [23] | |
| Clonostachys rosea | - | [11] | ||
| (-)-Vertinolide (143) | Clonostachys rosea B5-2 | - | [58] | |
| (-)-Dihydrovertinolide (144) | Clonostachys rosea B5-2 | Phytotoxic activity | [58] | |
| Clonostachydiol (145) | Clonostachys cylindrospora FH-A 6607 | Anthelmintic activity | [59] | |
| Bionectriol A (146) | Bionectria sp. | - | [60] | |
| Bionectriol B (147) | Bionectria ochroleuca | - | [61] | |
| Bionectriol C (148) | Bionectria ochroleuca | Antifungal activity | [61] | |
| Bionectriol D (149) | Bionectria ochroleuca | - | [61] | |
| Rogerson A (150) | Clonostachys rogersoniana | - | [62] | |
| Rogerson B (151) | Clonostachys rogersoniana | - | [62] | |
| Nectriacid A (152) | Nectria sp. HN001 | - | [47] | |
| Nectriacid B (153) | Nectria sp. HN001 | Inhibitory activity on α-glucosidase | [47] | |
| Nectriacid C (154) | Nectria sp. HN001 | Inhibitory activity on α-glucosidase | [47] | |
| Curvularin (155) | Clonostachys compactiuscula FKR-0021 | - | [19] | |
| α,β-Dehydrocurvularin (156) | Nectria glligena | Cytotoxic and phytotoxic activities | [63] | |
| Nectriatone B (157) | Nectria sp. B-13 | - | [64] | |
| Nectriatone C (158) | Nectria sp. B-13 | - | [64] | |
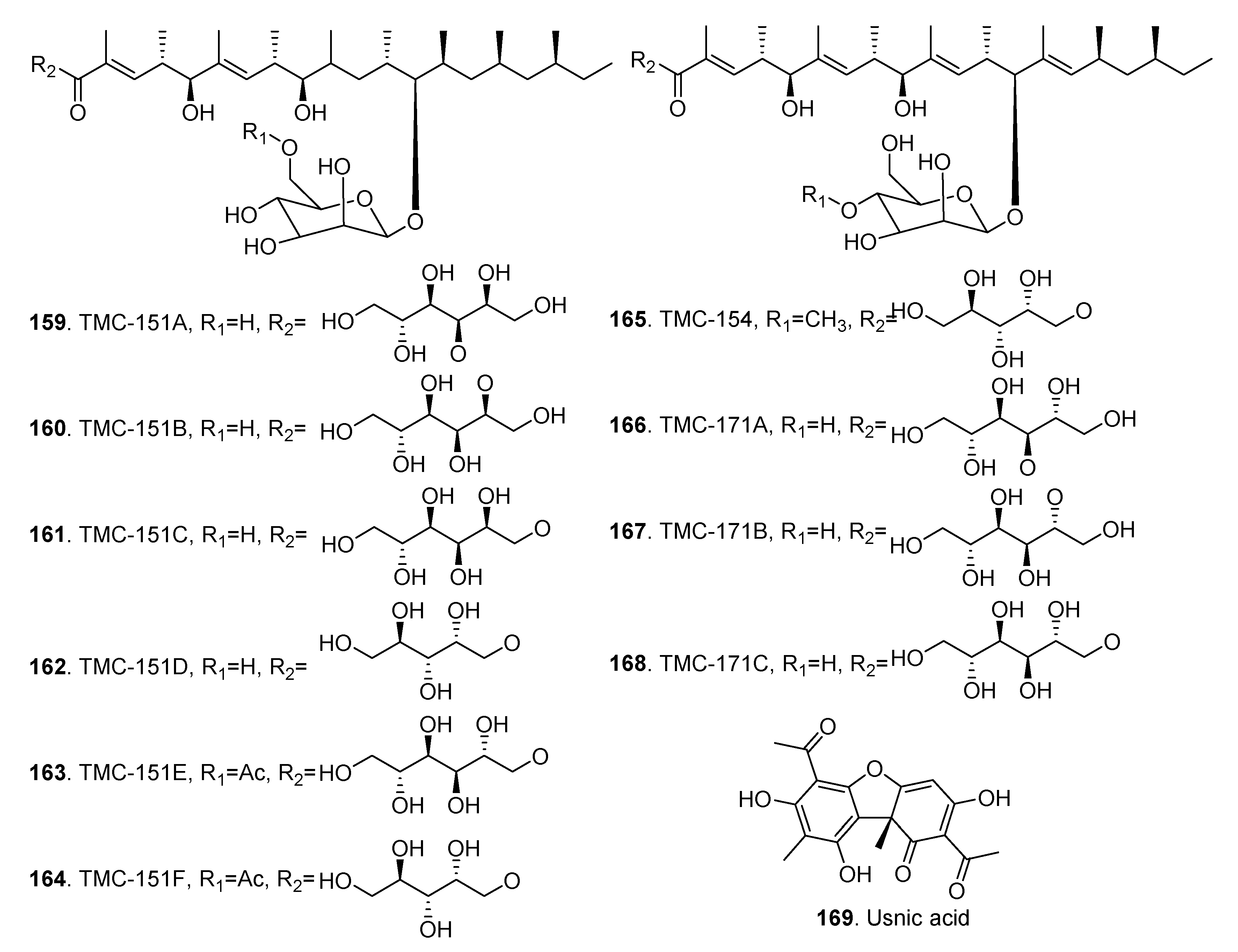
| TMC-151A (159) | Clonostachys rosea | - | [65] | |
| Gliocladium catenulatum | Moderate cytotoxicity to tumor cells | [66] | ||
| Bionectria ochroleuca | - | [67] | ||
| TMC-151B (160) | Clonostachys rosea | - | [67] | |
| Gliocladium catenulatum | Moderate cytotoxicity to tumor cells | [66] | ||
| TMC-151C (161) | Clonostachys rosea | - | [67] | |
| Gliocladium catenulatum | Moderate cytotoxicity to tumor cells | [66] | ||
| TMC-151D (162) | Clonostachys rosea | - | [67] | |
| Gliocladium catenulatum | Moderate cytotoxicity to tumor cells | [66] | ||
| TMC-151E (163) | Clonostachys rosea | - | [67] | |
| Gliocladium catenulatum | Moderate cytotoxicity to tumor cells | [66] | ||
| Bionectria ochroleuca | Antifungal activity | [61] | ||
| TMC-151F (164) | Clonostachys rosea | - | [67] | |
| Gliocladium catenulatum | Moderate cytotoxicity to tumor cells | [66] | ||
| Bionectria ochroleuca | Antifungal activity | [61] | ||
| TMC-154 (165) | Gliocladium roseum | - | [67] | |
| TMC-171A (166) | Gliocladium roseum | - | [67] | |
| TMC-171B (167) | Gliocladium roseum | - | [67] | |
| TMC-171C (168) | Gliocladium roseum | - | [67] | |
| Usnic acid (169) | Bionectria ochroleuca Bo-1 | Antibacterial activity | [68] |
| Metabolite Class | Metabolite Name | Fungal Species | Biological Activity | Ref. |
|---|---|---|---|---|
| Monoterpenoids | Nectriapyrone (170) | Nectria sp. HLS206 | - | [72] |
| Nectriapyrone C (171) | Nectria sp. HLS206 | - | [72] | |
| Nectriapyrone D (172) | Nectria sp. HLS206 | - | [72] | |
| Sesquiterpenoids | 5,6-Dihydroxybisabolol (173) | Bionectria sp. MSX 47401 | - | [3] |
| Nectrianolin C (174) | Nectria pseudotrichia 120-1NP | Cytotoxic activity | [73] | |
| 10-Acetyl trichoderonic acid A (175) | Nectria pseudotrichia | Leishmanicidal activity | [30] | |
| Hydroheptelidic acid (176) | Nectria pseudotrichia | Leishmanicidal activity | [30] | |
| Ophioceric acid (177) | Clonostachys compactiuscula FKR-0021 | - | [19] | |
| Xylaric acid D (178) | Nectria pseudotrichia | - | [30] | |
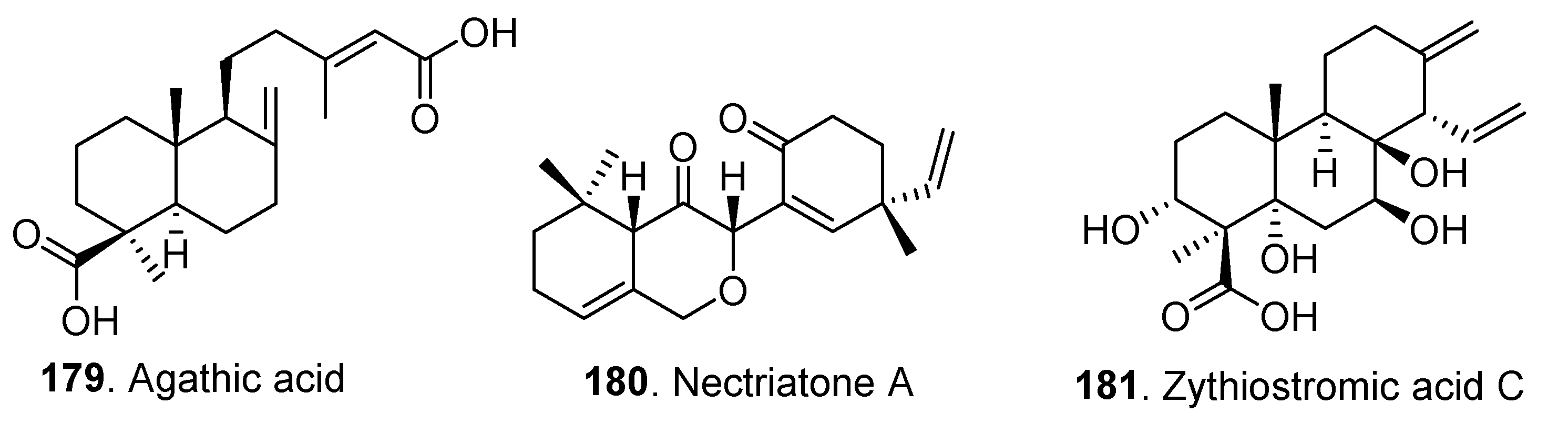
| Diterpenoids | Agathic acid (179) | Bionectria sp. | - | [28] |
| Nectriatone A (180) | Nectria sp. B-13 | Cytotoxic activity | [64] | |
| Zythiostromic acid C (181) | Nectria pseudotrichia 120-1NP | - | [50] | |
| Triterpentoids | Eburicol (182) | Clonostachys rosea MMS1090 | Cytotoxic activity | [74] |
| Helvolic acid (183) | Nectria sp. | - | [51] | |
| Antimicrobial activity | [75,76] | |||
| Polyterpenoids | Glisoprenin A (184) | Gliocladium sp. FO-1513 | Inhibition on acyl-CoA:cholesterol acyltransferase | [77] |
| Glisoprenin B (185) | Gliocladium sp. FO-1513 | Inhibition on acyl-CoA:cholesterol acyltransferase | [77] | |
| Glisoprenin C (186) | Gliocladium roseum HA190-95 | Inhibition on appressorium formation of Magnaporthe grisea | [78] | |
| Glisoprenin D (187) | Gliocladium roseum HA190-95 | Inhibition on appressorium formation of Magnaporthe grisea | [78] | |
| Glisoprenin E (188) | Gliocladium roseum HA190-95 | Inhibition on appressorium formation of Magnaporthe grisea | [78] | |
| Bionectin F (189) | Bionectria sp. Y1085 | - | [17] | |
| Meroterpenoids | Ascochlorin = Ilicicolin D = LL-Z 1272γ (190) | Nectria lucida | - | [79] |
| Nectria sp. | Antifungal activity | [8] | ||
| Nectria sp. B-13 | - | [34] | ||
| Nectria sp. B-13 | Cytotoxic activity | [64] | ||
| Dechloroilicicolin D = Cylindrol (191) | Nectria sp. | Antifungal activity | [8] | |
| 3-Bromoascochlorin (192) | Nectria coccinea | - | [80] | |
| Chloronectrin (193) | Nectria coccinea | - | [80] | |
| Deacetylchloronectrin (194) | Nectria sp. B-13 | - | [34] | |
| Dechlorodihydroascochlorin = Dechloro-12,13-dihydroascochlorin = LL-Z 1272ε (195) | Nectria lucida | - | [79] | |
| Nectria sp. B-13 | - | [34] | ||
| Ilicicolin C = LL-Z 1272δ (196) | Nectria sp. B-13 | - | [34] | |
| Nectria galligena | Inhibitory activity on AChE and α-glucuronidase | [63] | ||
| Ilicicolin E = 8′,9′-Dehydroascochorin = Cylindrochlorin (197) | Nectria sp. B-13 | Antibacterial activity | [34] | |
| Nectria sp. | Antifungal activity | [8] | ||
| Nectria sp. B-13 | Cytotoxic and antibacterial activities | [64] | ||
| Nectria galligena | Inhibitory activity on AChE and α-glucuronidase | [63] | ||
| Ilicicolin F = LL-Z 1272ζ (198) | Nectria sp. B-13 | - | [34] | |
| Nectria galligena | - | [63] | ||
| Nectria sp. | Antifungal activity | [8] | ||
| Nectrianolin A (199) | Nectria pseudotrichia 120-1NP | Cytotoxic activity against HL60 and HeLa cells | [73] | |
| Nectrianolin B (200) | Nectria pseudotrichia 120-1NP | Cytotoxic activity against HL60 and HeLa cells | [73] | |
| Ascofuranone (201) | Nectria sp. | Antifungal activity | [8] | |
| Chalmicrin (202) | Nectria sp. HLS206 | - | [81] | |
| Colletochlorin B (203) | Nectria sp. B-13 | - | [34] | |
| Nectria galligena | Inhibitory activity on AChE and α-glucuronidase | [63] | ||
| Colletorin B (204) | Nectria galligena | - | [63] | |
| Ilicicolin A (205) | Nectria sp. B-13 | - | [34] | |
| Ilicicolin B = LL-Z 1272β (206) | Nectria coccinea | - | [80] | |
| Nectria lucida | - | [79] | ||
| MBJ-0009 (207) | Nectria sp. f26111 | Cytotoxic activity | [82] | |
| MBJ-0010 (208) | Nectria sp. f26111 | Cytotoxic activity | [82] | |
| Taxol = Paclitaxel (209) | Gliocladium sp. | Cytotoxicity on cancer cells | [83] |
| Metabolite Name | Fungal Species | Biological Activity | Ref. |
|---|---|---|---|
| 4-Hydroxybenzoic aldehyde (210) | Gliocladium roseum CGMCC 3.3657 | - | [87] |
| 4-Hydroxy-benzoic acid (211) | Gliocladium roseum CGMCC 3.3657 | - | [87] |
| 3,4-Dihydroxy-benzoic acid (212) | Gliocladium roseum CGMCC 3.3657 | - | [87] |
| 3,5-Dihydroxy-benzoic acid (213) | Gliocladium roseum CGMCC 3.3657 | - | [87] |
| 2,5-Dimethoxy-3,6-dimethylbenzene-1,4-diol (214) | Nectria coryli | - | [54] |
| 3,5-Dihydroxybenzyl alcohol (215) | Nectria sp. B-13 | - | [34] |
| 3,5-Dihydroxytoluene (216) | Nectria sp. B-13 | - | [34] |
| 5-n-Heneicosylresorcinol (217) | Gliocladium roseum YMF1.00133 | Antinematodal activity | [22] |
| Toluquinol (218) | Nectria erubescens | - | [57] |
| Clonostach acid A (219) | Clonostachys rosea B5-2 | - | [58] |
| Clonostach acid B (220) | Clonostachys rosea B5-2 | - | [58] |
| Clonostach acid C (221) | Clonostachys rosea B5-2 | - | [58] |
| 2-Furoic acid (222) | Bionectria sp. Y1085 | - | [17] |
| 5-Hydroxymethyl furoic acid (223) | Bionectria sp. Y1085 | - | [17] |
| 2-Hydroxy-5-hydroxymethyl furan (224) | Bionectria sp. Y1085 | - | [17] |
| 3-(R)-Piliformic acid (225) | Bionectria sp. | - | [28] |
| 3-(S)-Piliformic acid (226) | Nectria pseudotrichia | Leishmanicidal activity | [30] |
| 6′-Acetoxy-piliformic acid (227) | Nectria pseudotrichia | Leishmanicidal activity | [30] |
| 5′,6′-Dehydropiliformic acid (228) | Nectria pseudotrichia | - | [30] |
| Hypocrealesate (229) | Nectria sp. HLS206 | - | [81] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, P.; Zhang, X.; Xu, D.; Zhang, B.; Lai, D.; Zhou, L. Metabolites from Clonostachys Fungi and Their Biological Activities. J. Fungi 2020, 6, 229. https://doi.org/10.3390/jof6040229
Han P, Zhang X, Xu D, Zhang B, Lai D, Zhou L. Metabolites from Clonostachys Fungi and Their Biological Activities. Journal of Fungi. 2020; 6(4):229. https://doi.org/10.3390/jof6040229
Chicago/Turabian StyleHan, Peipei, Xuping Zhang, Dan Xu, Bowen Zhang, Daowan Lai, and Ligang Zhou. 2020. "Metabolites from Clonostachys Fungi and Their Biological Activities" Journal of Fungi 6, no. 4: 229. https://doi.org/10.3390/jof6040229
APA StyleHan, P., Zhang, X., Xu, D., Zhang, B., Lai, D., & Zhou, L. (2020). Metabolites from Clonostachys Fungi and Their Biological Activities. Journal of Fungi, 6(4), 229. https://doi.org/10.3390/jof6040229