The Role of RodA-Conserved Cysteine Residues in the Aspergillus fumigatus Conidial Surface Organization

 , , , , , ,
, , , , , , 
 and
and
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Disulfide Bonds are Necessary for the Structure of the Spontaneously Self-Assembling RodAp
3.2. Point-Mutations of the Conserved Cysteine Residues of RODA Resulted in the Lack of the Rodlet Layer on the Conidial Surface
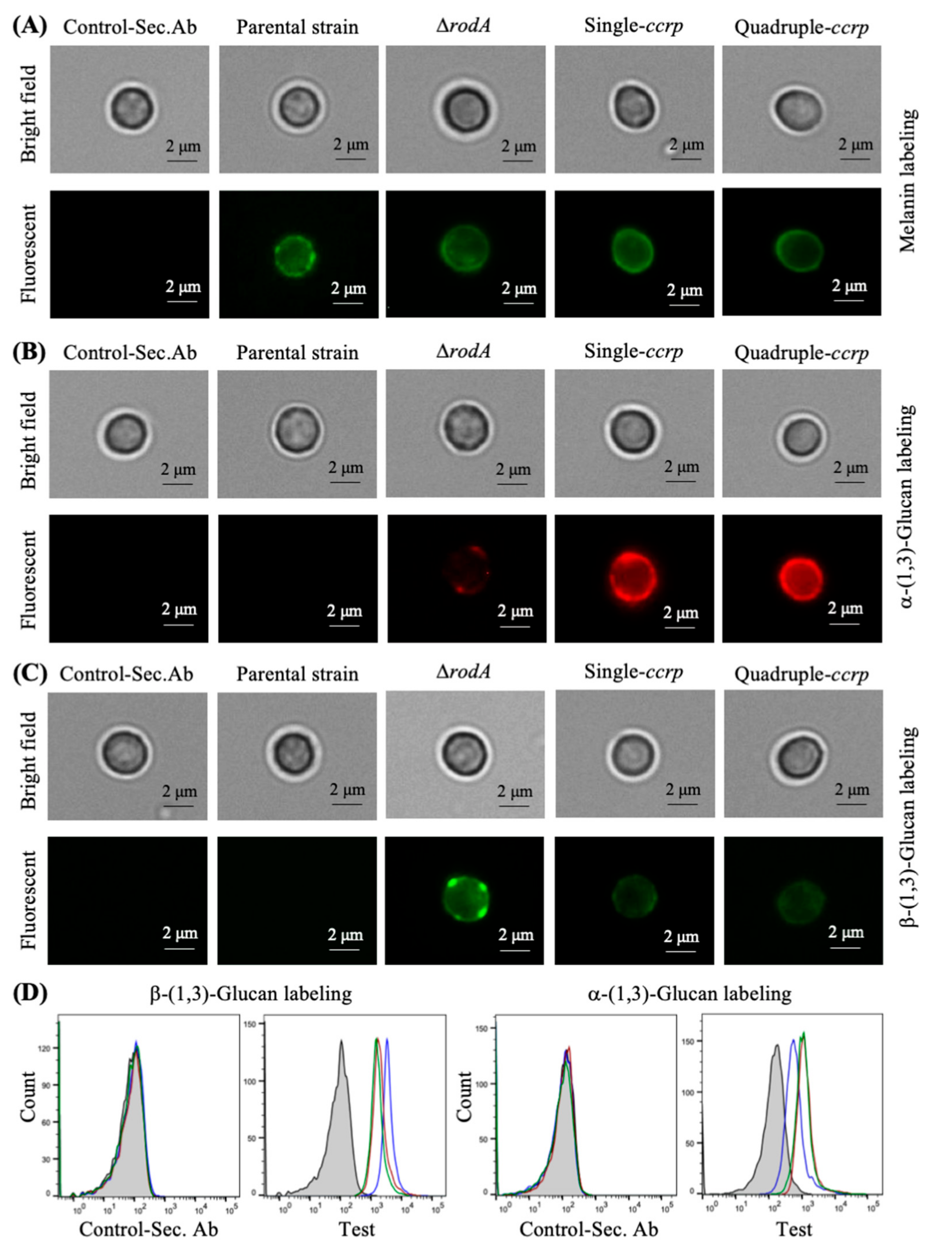
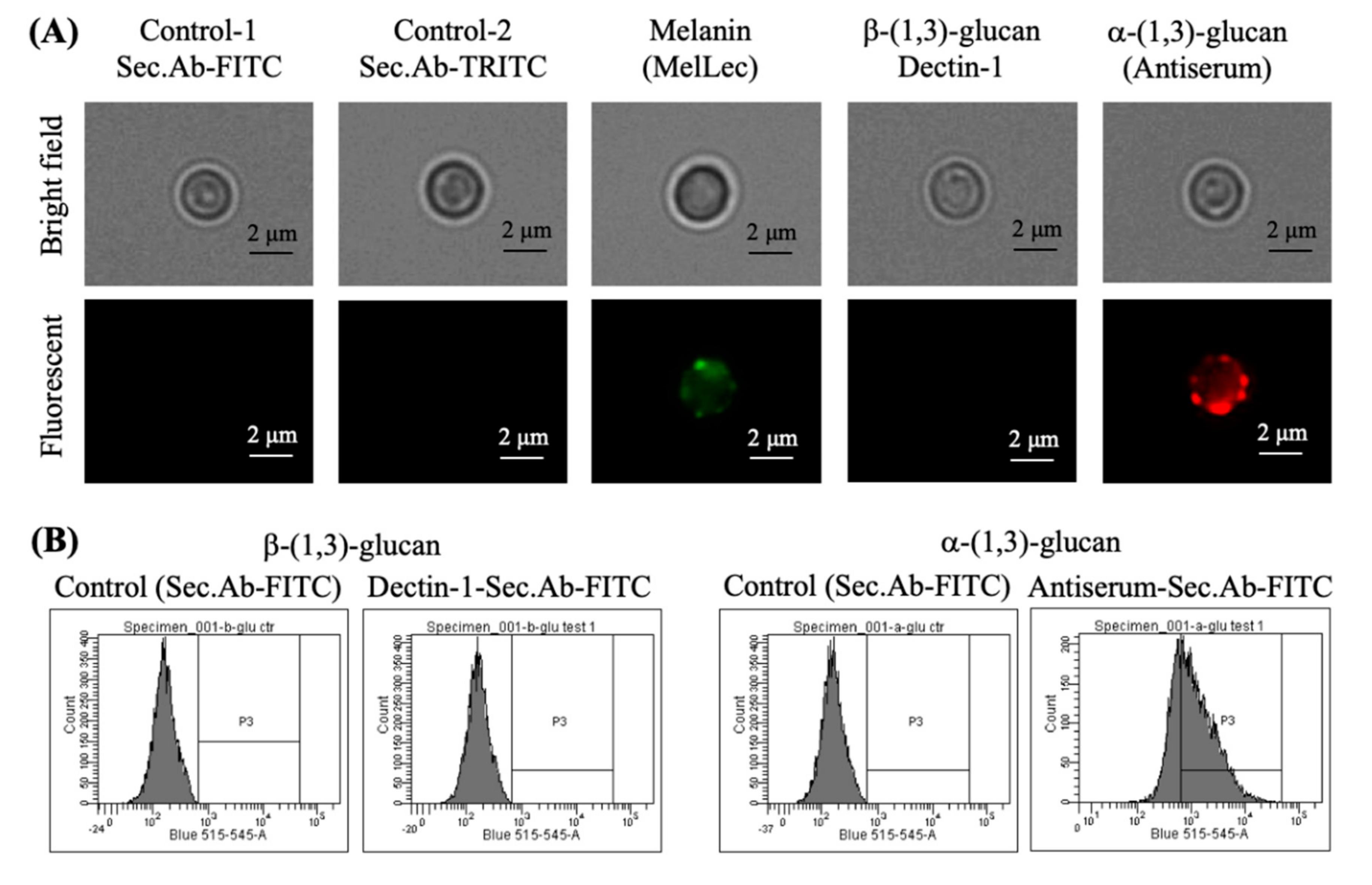
3.3. Absence of Rodlets Exposes α-(1,3)-Glucan on the Conidial Surface
3.4. The Proteins Exposed on the Ccrp-Mutant Conidial Surface are More Similar to Those of the Parental Strain
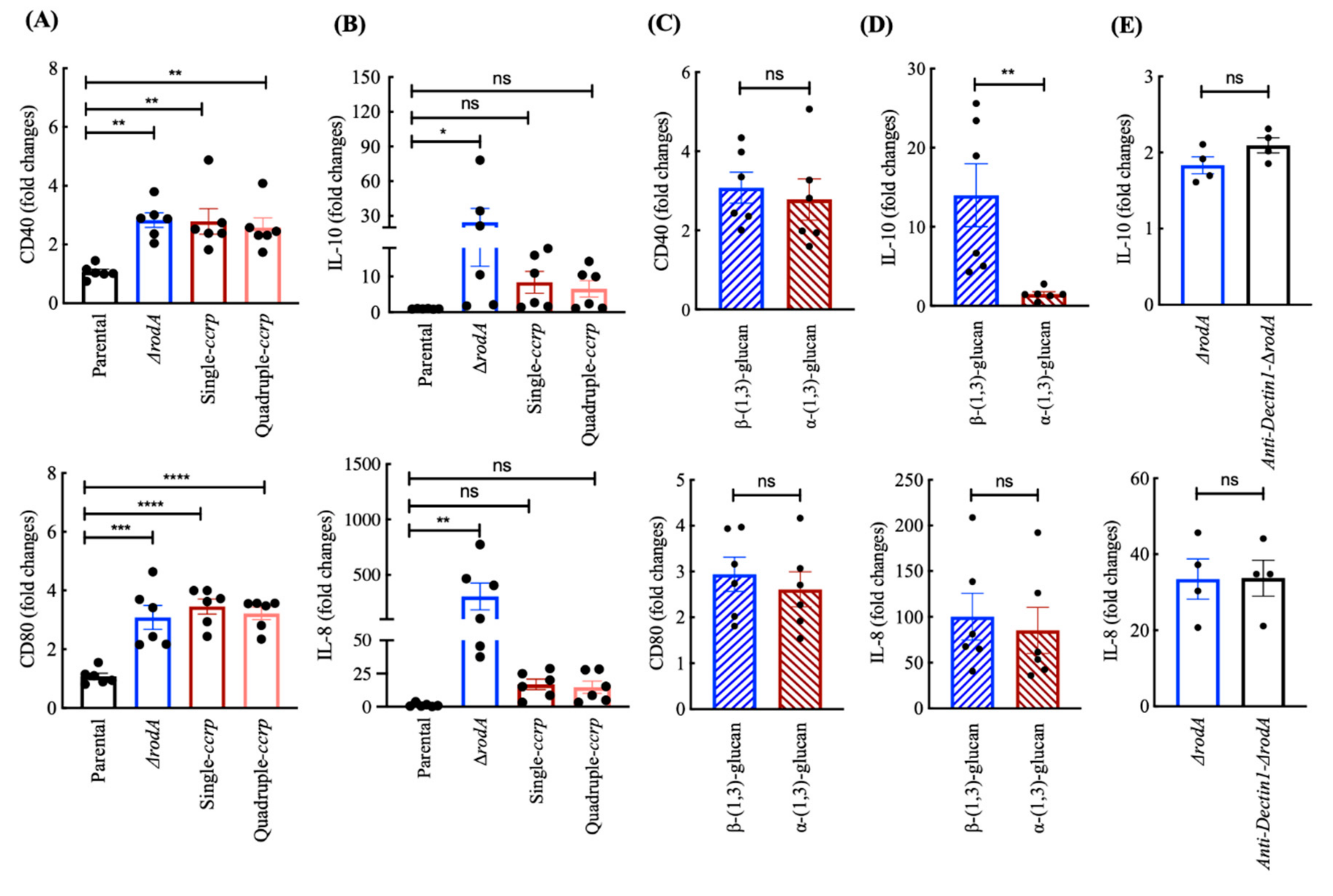
3.5. Ccrp-Mutant Conidia Are Less Stimulatory than ∆rodA Conidia
3.6. Chemical Removal of the Rodlet Layer Results in the Exposure of α-(1,3)-Glucan on the Conidial Surface
3.7. Cell Wall Components Exposed on the Ccrp-Mutant and Swollen Conidia Are Similar
4. Discussions
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Protein | Parental | ΔrodA | Single-ccrp Mutant | Quadruple-ccrp Mutant |
|---|---|---|---|---|---|
| Afu2g05635 | Afu2g05635 | 115 ± 28 | 71 ± 15 | 24 ± 21 | 0 |
| Afu5g09580 | RodA | 160 ± 112 | 0 | 36 ± 13 | 31 ± 7 |
| Afu6g14470 | Afu6g14470 | 55 ± 31 | 44 ± 10 | 15 ± 13 | 8 ± 2 |
| Afu4g09280 | Afu4g09280 | 38 ± 37 | 17 ± 12 | 7 ± 3 | 7 ± 3 |
| Afu4g09310 | Afu4g09310 | 40 ± 28 | 34 ± 26 | 34 ± 10 | 15 ± 3 |
| Afu4g09600 | Afu4g09600 | 132 ± 126 | 7 ± 7 | 0 | 0 |
| Afu1g13670 | CcpA | 33 ± 43 | 8 ± 9 | 0 | 0 |
| Afu6g03210 | ConJ | 10 ± 4 | 39 ± 26 | 20 ± 24 | 5 ± 2 |
| Afu2g09030 | DppV | 88 ± 95 | 104 ± 84 | 38 ± 16 | 23 ± 14 |
| Afu8g01980 | Afu8g01980 | 47 ± 57 | 21 ± 19 | 2 ± 2 | 0 |
| Afu2g00680 | Afu2g00680 | 35 ± 40 | 0 | 0 | 0 |
| Afu1g14450 | ExgO | 82 ± 67 | 35 ± 12 | 19 ± 20 | 2 ± 4 |
| Afu2g12630 | Aspf13 | 12 ± 14 | 0 | 0 | 0 |
| Afu2g17530 | Abr2 | 45 ± 49 | 14 ± 17 | 0 | 0 |
| Afu4g01030 | Afu4g01030 | 7 ± 8 | 0 | 0 | 0 |
| Afu7g01200 | Afu7g01200 | 19 ± 28 | 0 | 0 | 0 |
| Afu1g14560 | MsdS | 31 ± 43 | 57 ± 29 | 22 ± 20 | 10 ± 4 |
| Afu4g12450 | Afu4g12450 | 25 ± 36 | 7 ± 6 | 0 | 0 |
| Afu1g07440 | Hsp70 | 29 ± 6 | 33 ± 14 | 14 ± 8 | 5 ± 3 |
| Afu8g05410 | Afu8g05410 | 8 ± 11 | 0 | 0 | 0 |
| Afu3g15090 | Afu3g15090 | 21 ± 24 | 0 | 0 | 0 |
| Afu1g04670 | Afu1g04670 | 29 ± 26 | 0 | 0 | 0 |
| Afu5g11570 | Afu5g11570 | 17 ± 10 | 0 | 0 | 0 |
| Afu2g10670 | Afu2g10670 | 7 ± 8 | 3 ± 2 | 0 | 0 |
| Afu6g12070 | FmqD | 15 ± 21 | 11 ± 13 | 4 ± 4 | 0 |
| Afu2g03590 | Afu2g03590 | 2 ± 2 | 0 | 0 | 0 |
| Afu1g13195 | Afu1g13195 | 2 ± 2 | 3 ± 1 | 2 ± 2 | 0 |
| Afu6g07910 | Afu6g07910 | 10 ± 15 | 0 | 0 | 0 |
| Afu1g10790 | Afu1g10790 | 16 ± 23 | 5 ± 3 | 0 | 0 |
| Afu3g02270 | Cat1 | 13 ± 14 | 30 ± 10 | 9 ± 7 | 8 ± 4 |
| Afu1g10150 | Afu1g10150 | 5 ± 4 | 0 | 0 | 0 |
| Afu5g13920 | Wos2 | 3 ± 3 | 0 | 0 | 0 |
| Afu5g08020 | Afu5g08020 | 3 ± 3 | 0 | 0 | 0 |
| Afu4g06910 | Afu4g06910 | 2 ± 2 | 2 ± 3 | 0 | 0 |
| Afu3g06880 | SnxA | 0 | 19 ± 10 | 9 ± 3 | 8 ± 5 |
| Afu5g09530 | Afu5g09530 | 0 | 0 | 4 ± 4 | 2 ± 2 |
| Afu4g09320 | DppIV | 0 | 23 ± 21 | 4 ± 2 | 3 ± 3 |
| Afu5g14210 | Grg1 | 0 | 12 ± 8 | 6 ± 7 | 0 |
| Afu4g00860 | DprA | 0 | 19 ± 11 | 10 ± 13 | 0 |
| Afu5g12210 | SfaD | 0 | 0 | 1 ± 2 | 0 |
| Afu2g03980 | Afu2g03980 | 0 | 2 ± 2 | 2 ± 2 | 0 |
| Afu6g09980 | Ncr1 | 0 | 7 ± 6 | 3 ± 3 | 0 |
| Afu1g17370 | Scf1 | 0 | 8 ± 1 | 0 | 0 |
| Afu2g01050 | Afu2g01050 | 0 | 3 ± 2 | 0 | 0 |
| Afu5g03540 | Afu5g03540 | 0 | 13 ± 10 | 0 | 0 |
| Afu5g10570 | Afu5g10570 | 0 | 4 ± 5 | 0 | 0 |
| Afu1g05770 | Exg12 | 0 | 24 ± 21 | 0 | 0 |
| Afu8g05020 | NagA | 0 | 14 ± 10 | 0 | 0 |
| Afu6g13300 | Afu6g13300 | 0 | 3 ± 3 | 0 | 0 |
| Afu8g07060 | RodC | 0 | 2 ± 2 | 0 | 0 |
| Afu2g00967 | Afu2g00967 | 0 | 2 ± 2 | 0 | 0 |
| Afu3g07160 | Afu3g07160 | 0 | 4 ± 3 | 0 | 0 |
| Afu8g05320 | Afu8g05320 | 0 | 5 ± 3 | 0 | 0 |
| Afu1g06470 | Afu1g06470 | 0 | 6 ± 6 | 0 | 0 |
References
- Aimanianda, V.; Bayry, J.; Bozza, S.; Kniemeyer, O.; Perruccio, K.; Elluru, S.R.; Clavaud, C.; Paris, S.; Brakhage, A.A.; Kaveri, S.V.; et al. Surface hydrophobin prevents immune recognition of airborne fungal spores. Nature 2009, 460, 1117–1121. [Google Scholar] [CrossRef] [PubMed]
- Bertuzzi, M.; Hayes, G.E.; Icheoku, U.J.; van Rhijn, N.; Denning, D.W.; Osherov, N.; Bignell, E.M. Anti-aspergillus activities of the respiratory epithelium in health and disease. J. Fungi 2018, 4, 8. [Google Scholar] [CrossRef] [PubMed]
- van de Veerdonk, F.L.; Gresnigt, M.S.; Romani, L.; Netea, M.G.; Latge, J.P. Aspergillus fumigatus morphology and dynamic host interactions. Nat. Rev. Microbiol. 2017, 15, 661–674. [Google Scholar] [CrossRef]
- Hohl, T.M.; Van Epps, H.L.; Rivera, A.; Morgan, L.A.; Chen, P.L.; Feldmesser, M.; Pamer, E.G. Aspergillus fumigatus triggers inflammatory responses by stage-specific beta-glucan display. PLoS Pathog. 2005, 1, e30. [Google Scholar] [CrossRef] [PubMed]
- Slesiona, S.; Gressler, M.; Mihlan, M.; Zaehle, C.; Schaller, M.; Barz, D.; Hube, B.; Jacobsen, I.D.; Brock, M. Persistence versus escape: Aspergillus terreus and Aspergillus fumigatus employ different strategies during interactions with macrophages. PLoS ONE 2012, 7, e31223. [Google Scholar] [CrossRef]
- Bayry, J.; Aimanianda, V.; Guijarro, J.I.; Sunde, M.; Latge, J.P. Hydrophobins—Unique fungal proteins. PLoS Pathog. 2012, 8, e1002700. [Google Scholar] [CrossRef]
- Pille, A.; Kwan, A.H.; Cheung, I.; Hampsey, M.; Aimanianda, V.; Delepierre, M.; Latge, J.P.; Sunde, M.; Guijarro, J.I. (1)H, (13)C and (15)N resonance assignments of the RodA hydrophobin from the opportunistic pathogen Aspergillus fumigatus. Biomol. NMR Assign. 2015, 9, 113–118. [Google Scholar] [CrossRef]
- Jensen, B.G.; Andersen, M.R.; Pedersen, M.H.; Frisvad, J.C.; Sondergaard, I. Hydrophobins from Aspergillus species cannot be clearly divided into two classes. BMC Res. Notes 2010, 3, 344. [Google Scholar] [CrossRef]
- Valsecchi, I.; Lai, J.I.; Stephen-Victor, E.; Pille, A.; Beaussart, A.; Lo, V.; Pham, C.L.L.; Aimanianda, V.; Kwan, A.H.; Duchateau, M.; et al. Assembly and disassembly of aspergillus fumigatus conidial rodlets. Cell Surf. 2019, 5, 100023. [Google Scholar] [CrossRef]
- da Silva Ferreira, M.E.; Kress, M.R.; Savoldi, M.; Goldman, M.H.; Hartl, A.; Heinekamp, T.; Brakhage, A.A.; Goldman, G.H. The akuB(KU80) mutant deficient for nonhomologous end joining is a powerful tool for analyzing pathogenicity in Aspergillus fumigatus. Eukaryot Cell 2006, 5, 207–211. [Google Scholar] [CrossRef]
- Thau, N.; Monod, M.; Crestani, B.; Rolland, C.; Tronchin, G.; Latge, J.P.; Paris, S. Rodletless mutants of Aspergillus fumigatus. Infect. Immun. 1994, 62, 4380–4388. [Google Scholar] [CrossRef]
- Rosas, A.L.; Nosanchuk, J.D.; Gomez, B.L.; Edens, W.A.; Henson, J.M.; Casadevall, A. Isolation and serological analyses of fungal melanins. J. Immunol. Methods 2000, 244, 69–80. [Google Scholar] [CrossRef]
- Stappers, M.H.T.; Clark, A.E.; Aimanianda, V.; Bidula, S.; Reid, D.M.; Asamaphan, P.; Hardison, S.E.; Dambuza, I.M.; Valsecchi, I.; Kerscher, B.; et al. Recognition of DHN-melanin by a C-type lectin receptor is required for immunity to Aspergillus. Nature 2018, 555, 382–386. [Google Scholar] [CrossRef] [PubMed]
- Komarova, B.S.; Orekhova, M.V.; Tsvetkov, Y.E.; Beau, R.; Aimanianda, V.; Latge, J.P.; Nifantiev, N.E. Synthesis of a pentasaccharide and neoglycoconjugates related to fungal alpha-(1–>3)-glucan and their use in the generation of antibodies to trace Aspergillus fumigatus cell wall. Chemistry 2015, 21, 1029–1035. [Google Scholar] [CrossRef] [PubMed]
- Voltersen, V.; Blango, M.G.; Herrmann, S.; Schmidt, F.; Heinekamp, T.; Strassburger, M.; Kruger, T.; Bacher, P.; Lother, J.; Weiss, E.; et al. Proteome analysis reveals the conidial surface protein ccpa essential for virulence of the pathogenic fungus aspergillus fumigatus. mBio 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Blango, M.G.; Pschibul, A.; Rivieccio, F.; Kruger, T.; Rafiq, M.; Jia, L.J.; Zheng, T.; Goldmann, M.; Voltersen, V.; Li, J.; et al. Dynamic surface proteomes of allergenic fungal conidia. J. Proteome Res. 2020, 19, 2092–2104. [Google Scholar] [CrossRef]
- Brakhage, A.A.; Van den Brulle, J. Use of reporter genes to identify recessive trans-acting mutations specifically involved in the regulation of Aspergillus nidulans penicillin biosynthesis genes. J. Bacteriol. 1995, 177, 2781–2788. [Google Scholar] [CrossRef]
- Maerker, C.; Rohde, M.; Brakhage, A.A.; Brock, M. Methylcitrate synthase from Aspergillus fumigatus. Propionyl-CoA affects polyketide synthesis, growth and morphology of conidia. FEBS J. 2005, 272, 3615–3630. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef]
- Holla, S.; Sharma, M.; Vani, J.; Kaveri, S.V.; Balaji, K.N.; Bayry, J. GM-CSF along with IL-4 but not alone is indispensable for the differentiation of human dendritic cells from monocytes. J. Allergy Clin. Immunol. 2014, 133, 1500–1502.e1. [Google Scholar] [CrossRef]
- Stephen-Victor, E.; Karnam, A.; Fontaine, T.; Beauvais, A.; Das, M.; Hegde, P.; Prakhar, P.; Holla, S.; Balaji, K.N.; Kaveri, S.V.; et al. Aspergillus fumigatus cell wall alpha-(1,3)-glucan stimulates regulatory T-Cell polarization by Inducing PD-L1 expression on human dendritic cells. J. Infect. Dis. 2017, 216, 1281–1294. [Google Scholar] [CrossRef] [PubMed]
- Steele, C.; Rapaka, R.R.; Metz, A.; Pop, S.M.; Williams, D.L.; Gordon, S.; Kolls, J.K.; Brown, G.D. The beta-glucan receptor dectin-1 recognizes specific morphologies of Aspergillus fumigatus. PLoS Pathog. 2005, 1, e42. [Google Scholar] [CrossRef] [PubMed]
- Pihet, M.; Vandeputte, P.; Tronchin, G.; Renier, G.; Saulnier, P.; Georgeault, S.; Mallet, R.; Chabasse, D.; Symoens, F.; Bouchara, J.P. Melanin is an essential component for the integrity of the cell wall of Aspergillus fumigatus conidia. BMC Microbiol. 2009, 9, 177. [Google Scholar] [CrossRef] [PubMed]
- Bozza, S.; Clavaud, C.; Giovannini, G.; Fontaine, T.; Beauvais, A.; Sarfati, J.; D’Angelo, C.; Perruccio, K.; Bonifazi, P.; Zagarella, S.; et al. Immune sensing of Aspergillus fumigatus proteins, glycolipids, and polysaccharides and the impact on Th immunity and vaccination. J. Immunol. 2009, 183, 2407–2414. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, X.M.; Heflin, K.E.; Lavigne, L.M.; Yu, K.; Kim, M.; Salomon, A.R.; Reichner, J.S. Lectin site ligation of CR3 induces conformational changes and signaling. J. Biol. Chem. 2012, 287, 3337–3348. [Google Scholar] [CrossRef]
- Fontaine, T.; Beauvais, A.; Loussert, C.; Thevenard, B.; Fulgsang, C.C.; Ohno, N.; Clavaud, C.; Prevost, M.C.; Latge, J.P. Cell wall alpha1-3glucans induce the aggregation of germinating conidia of Aspergillus fumigatus. Fungal Genet. Biol. 2010, 47, 707–712. [Google Scholar] [CrossRef]
- Beauvais, A.; Bozza, S.; Kniemeyer, O.; Formosa, C.; Balloy, V.; Henry, C.; Roberson, R.W.; Dague, E.; Chignard, M.; Brakhage, A.A.; et al. Deletion of the alpha-(1,3)-glucan synthase genes induces a restructuring of the conidial cell wall responsible for the avirulence of Aspergillus fumigatus. PLoS Pathog. 2013, 9, e1003716. [Google Scholar] [CrossRef]
- Dichtl, K.; Samantaray, S.; Aimanianda, V.; Zhu, Z.; Prevost, M.C.; Latge, J.P.; Ebel, F.; Wagener, J. Aspergillus fumigatus devoid of cell wall beta-1,3-glucan is viable, massively sheds galactomannan and is killed by septum formation inhibitors. Mol. Microbiol. 2015, 95, 458–471. [Google Scholar] [CrossRef]
- Henry, C.; Fontaine, T.; Heddergott, C.; Robinet, P.; Aimanianda, V.; Beau, R.; Beauvais, A.; Mouyna, I.; Prevost, M.C.; Fekkar, A.; et al. Biosynthesis of cell wall mannan in the conidium and the mycelium of Aspergillus fumigatus. Cell Microbiol. 2016, 18, 1881–1891. [Google Scholar] [CrossRef]
- Herrero, A.B.; Magnelli, P.; Mansour, M.K.; Levitz, S.M.; Bussey, H.; Abeijon, C. KRE5 gene null mutant strains of Candida albicans are avirulent and have altered cell wall composition and hypha formation properties. Eukaryot. Cell 2004, 3, 1423–1432. [Google Scholar] [CrossRef]
- Gilbert, N.M.; Donlin, M.J.; Gerik, K.J.; Specht, C.A.; Djordjevic, J.T.; Wilson, C.F.; Sorrell, T.C.; Lodge, J.K. KRE genes are required for beta-1,6-glucan synthesis, maintenance of capsule architecture and cell wall protein anchoring in Cryptococcus neoformans. Mol. Microbiol. 2010, 76, 517–534. [Google Scholar] [CrossRef] [PubMed]
- Dennehy, K.M.; Willment, J.A.; Williams, D.L.; Brown, G.D. Reciprocal regulation of IL-23 and IL-12 following co-activation of Dectin-1 and TLR signaling pathways. Eur. J. Immunol. 2009, 39, 1379–1386. [Google Scholar] [CrossRef] [PubMed]
- Kang, X.; Kirui, A.; Muszynski, A.; Widanage, M.C.D.; Chen, A.; Azadi, P.; Wang, P.; Mentink-Vigier, F.; Wang, T. Molecular architecture of fungal cell walls revealed by solid-state NMR. Nat. Commun. 2018, 9, 2747. [Google Scholar] [CrossRef]
- Chai, L.Y.; Kullberg, B.J.; Vonk, A.G.; Warris, A.; Cambi, A.; Latge, J.P.; Joosten, L.A.; van der Meer, J.W.; Netea, M.G. Modulation of Toll-like receptor 2 (TLR2) and TLR4 responses by Aspergillus fumigatus. Infect. Immun. 2009, 77, 2184–2192. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valsecchi, I.; Stephen-Victor, E.; Wong, S.S.W.; Karnam, A.; Sunde, M.; Guijarro, J.I.; Rodríguez de Francisco, B.; Krüger, T.; Kniemeyer, O.; Brown, G.D.; et al. The Role of RodA-Conserved Cysteine Residues in the Aspergillus fumigatus Conidial Surface Organization. J. Fungi 2020, 6, 151. https://doi.org/10.3390/jof6030151
Valsecchi I, Stephen-Victor E, Wong SSW, Karnam A, Sunde M, Guijarro JI, Rodríguez de Francisco B, Krüger T, Kniemeyer O, Brown GD, et al. The Role of RodA-Conserved Cysteine Residues in the Aspergillus fumigatus Conidial Surface Organization. Journal of Fungi. 2020; 6(3):151. https://doi.org/10.3390/jof6030151
Chicago/Turabian StyleValsecchi, Isabel, Emmanuel Stephen-Victor, Sarah Sze Wah Wong, Anupama Karnam, Margaret Sunde, J. Iñaki Guijarro, Borja Rodríguez de Francisco, Thomas Krüger, Olaf Kniemeyer, Gordon D. Brown, and et al. 2020. "The Role of RodA-Conserved Cysteine Residues in the Aspergillus fumigatus Conidial Surface Organization" Journal of Fungi 6, no. 3: 151. https://doi.org/10.3390/jof6030151
APA StyleValsecchi, I., Stephen-Victor, E., Wong, S. S. W., Karnam, A., Sunde, M., Guijarro, J. I., Rodríguez de Francisco, B., Krüger, T., Kniemeyer, O., Brown, G. D., Willment, J. A., Latgé, J.-P., Brakhage, A. A., Bayry, J., & Aimanianda, V. (2020). The Role of RodA-Conserved Cysteine Residues in the Aspergillus fumigatus Conidial Surface Organization. Journal of Fungi, 6(3), 151. https://doi.org/10.3390/jof6030151