
Biocontrol Properties of Basidiomycetes: An Overview

 ,
,
Abstract
:1. Introduction
Plant Fungal Diseases
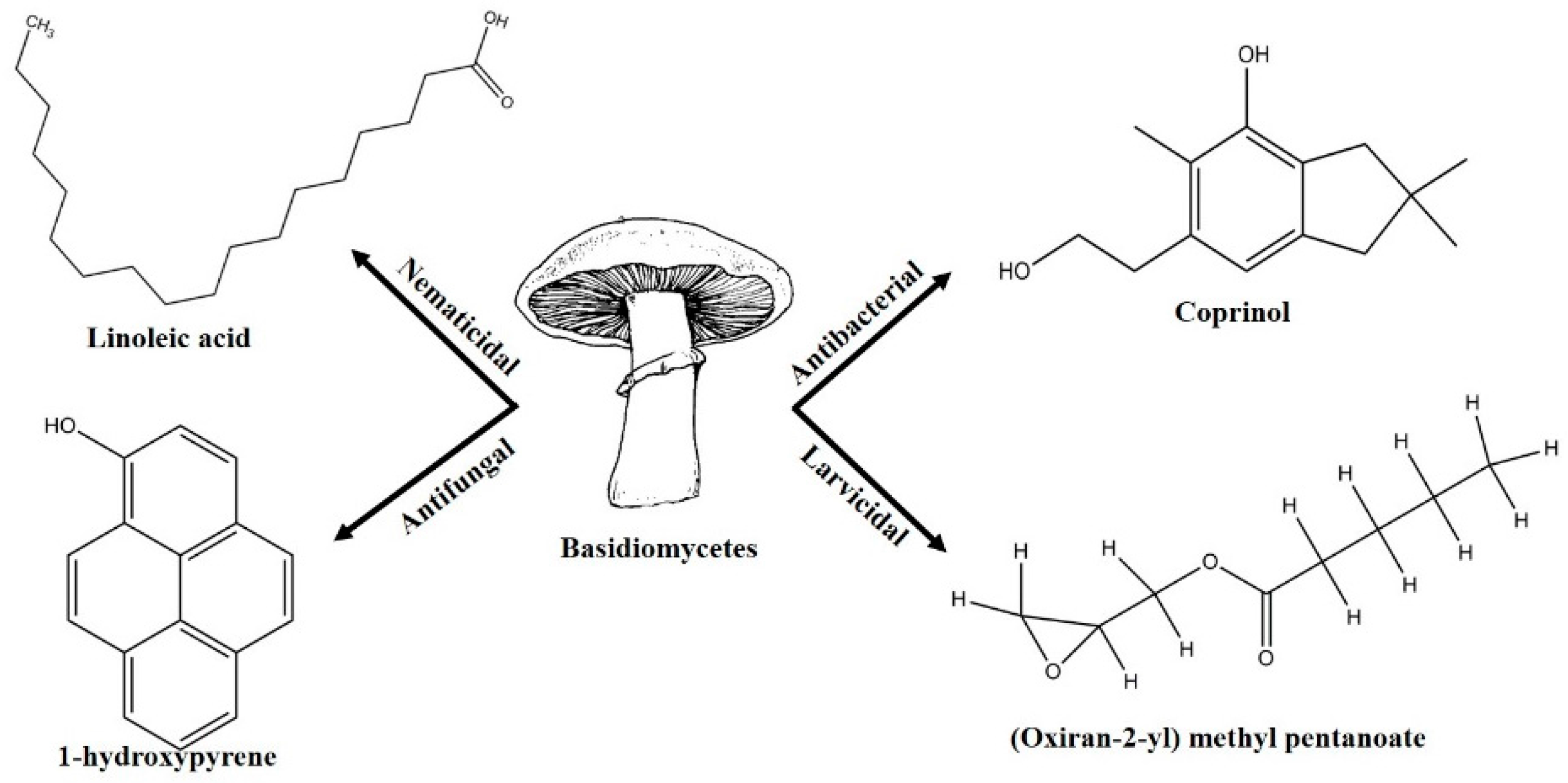
2. Biocontrol Properties of Basidiomycetes
2.1. Anti-Phytofungal Activity (Table 1)
2.2. Anti-Phytobacterial Activities
2.3. Anti-Phytoviral Activity
2.4. Phytonematicidal Activity
2.5. Mosquito Larvicidal Activity
3. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Whipps, J.M.; Lumsden, R.D. Commercial use of fungi as plant disease biological control agents: Status and prospects. In Fungal Biocontrol Agents: Progress, Problems and Potential; Butt, T.M., Jackson, C., Eds.; CABI Publishing: Oxfordshire, UK, 2001; Volume 9, pp. 9–22. [Google Scholar]
- Maxmen, A. Crop pests: Under attack. Nature 2013, 501, S15–S17. [Google Scholar] [CrossRef] [PubMed]
- Montesinos, E. Development, registration and commercialization of microbial pesticides for plant protection. Int. Microbiol. 2003, 6, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, R.; Dey, U. An overview of fungal and bacterial biopesticides to control plant pathogens/diseases. Afr. J. Microbiol. Res. 2014, 8, 1749–1762. [Google Scholar]
- Reegan, A.D.; Gandhi, M.R.; Paulraj, M.G.; Ignacimuthu, S. Ovicidal and oviposition deterrent activities of medicinal plant extracts against Aedes aegypti L. and Culex quinquefasciatus Say mosquitoes (Diptera: Culicidae). Osong Public Health Res. Perspect. 2015, 6, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Dhiman, R.C.; Pahwa, S.; Dhillon, G.P.; Dash, A.P. Climate change and threat of vector-borne diseases in India: Are we prepared? Parasitol. Res. 2010, 106, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Benelli, G. Research in mosquito control: Current challenges for a brighter future. Parasitol. Res. 2015, 114, 2801–2805. [Google Scholar] [CrossRef] [PubMed]
- Williamson, V.M.; Gleason, C.A. Plant-nematode interactions. Curr. Opin. Plant Biol. 2003, 6, 327–333. [Google Scholar] [CrossRef]
- Dropkin, V.H. Introduction to Plant Nematology, 2nd ed.; John Wiley and Sons Inc.: Columbia, SC, USA, 1989; pp. 1–304. [Google Scholar]
- Campbell, J.F.; Kaya, H.K. How and why a parasitic nematode jumps. Nature 1999, 397, 485–486. [Google Scholar] [CrossRef]
- Spread of Crop Pest Threatens Global Food Security as Earth Warms. Available online: http://www.exeter.ac.uk/research/inspiring/keythemes/science/news/archive/title_316965_en.html (accessed on 1 September 2013).
- Gillespie, A. Conservation, Biodiversity and International Law; Edward Elgar Publishing: Chelteham, New Zealand, 2013; pp. 1–579. [Google Scholar]
- Wasser, S.P.; Weis, A.L. Medicinal properties of substances occurring in higher basidiomycetes mushrooms: Current perspectives (review). Int. J. Med. Mushrooms 1999, 1, 31–62. [Google Scholar] [CrossRef]
- Anke, T. Basidiomycetes: A source for new bioactive secondary metabolites. Prog. Ind. Microbiol. 1989, 27, 51–66. [Google Scholar]
- Gowrie, S.U.; Chathurdevi, G.; Rani, K. Evaluation of bioactive potential of basidiocarp extracts of Ganoderma lucidum. Int. J. Pharm. Res. Allied Sci. 2014, 3, 36–46. [Google Scholar]
- Kolundžić, M.; Grozdanić, N.Đ.; Dodevska, M.; Milenković, M.; Sisto, F.; Miani, A.; Farronato, G.; Kundaković, T. Antibacterial and cytotoxic activities of wild mushroom Fomes fomentarius (L.) Fr., Polyporaceae. Ind. Crops. Prod. 2016, 79, 110–115. [Google Scholar]
- Ren, L.; Hemar, Y.; Perera, C.O.; Lewis, G.; Krissansen, G.W.; Buchanan, P.K. Antibacterial and antioxidant activities of aqueous extracts of eight edible mushrooms. Bioact. Carbohydr. Diet. Fibre 2014, 3, 41–51. [Google Scholar] [CrossRef]
- Santos, D.N.; de Souza, L.L.; de Oliveira, C.A.; Silva, E.R.; de Oliveira, A.L. Arginase inhibition, antibacterial and antioxidant activities of Pitanga seed (Eugenia uniflora L.) extracts from sustainable technologies of high pressure extraction. Food Biosci. 2015, 12, 93–99. [Google Scholar] [CrossRef]
- Wang, H.; Ng, T. Ganodermin, an antifungal protein from fruiting bodies of the medicinal mushroom Ganoderma lucidum. Peptides 2006, 27, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Chu, K.; Xia, L.; Ng, T. Pleurostrin, an antifungal peptide from the oyster mushroom. Peptides 2005, 26, 2098–2103. [Google Scholar] [CrossRef] [PubMed]
- Sajeena, A.; Marimuthu, T. Efficacy, stability and persistence of Ganosol, a Ganoderma based fungicide against plant pathogens. J. Plant Prot. Sci. 2013, 5, 17–25. [Google Scholar]
- Lam, S.; Ng, T. First simultaneous isolation of a ribosome inactivating protein and an antifungal protein from a mushroom (Lyophyllum shimeji) together with evidence for synergism of their antifungal effects. Arch. Biochem. Biophys. 2001, 393, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ng, T. Eryngin, a novel antifungal peptide from fruiting bodies of the edible mushroom Pleurotus eryngii. Peptides 2004, 25, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Wasser, S.P.; Weis, A.L. Therapeutic effects of substances occurring in higher Basidiomycetes mushrooms: A modern perspective. Crit. Rev. Immunol. 1999, 19, 65–96. [Google Scholar] [PubMed]
- Faccin, L.C.; Benati, F.; Rincão, V.P.; Mantovani, M.S.; Soares, S.A.; Gonzaga, M.L.; Nozawa, C.; Carvalho, L.R. Antiviral activity of aqueous and ethanol extracts and of an isolated polysaccharide from Agaricus brasiliensis against poliovirus type 1. Lett. Appl. Microbiol. 2007, 45, 24–28. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Zhao, C.G.; Tong, X.; Qi, Y.P. A lectin with mycelia differentiation and antiphytovirus activities from the edible mushroom Agrocybe aegerita. J. Biochem. Mol. Biol. 2003, 36, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Triguéros, V.; Paquereau, L.; Chavant, L.; Fournier, D. Proteins as active compounds involved in insecticidal activity of mushroom fruitbodies. J. Econ. Entomol. 2002, 95, 603–607. [Google Scholar] [CrossRef] [PubMed]
- Jo, W.S.; Hossain, M.A.; Park, S.C. Toxicological profiles of poisonous, edible, and medicinal mushrooms. Mycobiology 2014, 42, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Wasser, S.P. A Book Review: The Fungal Pharmacy: Medicinal Mushrooms of Western Canada (Robert Rogers, 2006, Prairie Deva Press, Edmonton Alberta, 234 pp., $39.95 CDN). Int. J. Med. Mushrooms. 2008, 10, 97–100. [Google Scholar] [CrossRef]
- Valverde, M.E.; Hernández-Pérez, T.; Paredes-López, O. Edible mushrooms: Improving human health and promoting quality life. Int. J. Microbiol. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Wasser, S.P. Current findings, future trends, and unsolved problems in studies of medicinal mushrooms. Antimicrob. Agents Chemother. 2011, 89, 1323–1332. [Google Scholar] [CrossRef] [PubMed]
- Richard, N.S. Introduction to Plant Pathology; John Wiley & Sons: New York, NY, USA, 2006; pp. 1–50. [Google Scholar]
- Knogge, W. Fungal infection of plants. Plant Cell 1996, 8, 1711–1722. [Google Scholar] [CrossRef] [PubMed]
- Bronson, C.R. The genetics of phytotoxin production by plant pathogenic fungi. Experientia 1991, 47, 771–776. [Google Scholar] [CrossRef]
- Than, P.P.; Prihastuti, H.; Phoulivong, S.; Taylor, P.W.; Hyde, K.D. Chilli anthracnose disease caused by Colletotrichum species. J. Zhejiang Univ. Sci. B 2008, 9, 764–778. [Google Scholar] [CrossRef] [PubMed]
- Hayden, H.P.K.; Aitken, E.; Irwin, J. Genetic-relationships as assessed by molecular markers and cross-infection among strains of Colletotrichum gloeosporioides. Aust. J. Bot. 1994, 42, 9–18. [Google Scholar] [CrossRef]
- Braithwaite, K.S.; Irwin, J.A.; Manners, J.M. Restriction fragment length polymorphisms in Colletotrichum gloeosporioides infecting Stylosanthes spp. in Australia. Mycol. Res. 1990, 94, 1129–1137. [Google Scholar] [CrossRef]
- Talbot, N.J. On the trail of a cereal killer: Exploring the biology of Magnaporthe grisea. Annu. Rev. Microbiol. 2003, 57, 177–202. [Google Scholar] [CrossRef] [PubMed]
- Hafner, S. Trends in maize, rice, and wheat yields for 188 nations over the past 40 years: A prevalence of linear growth. Agric. Ecosyst. Environ. 2003, 97, 275–283. [Google Scholar] [CrossRef]
- Thinlay, X.; Finckh, M.R.; Bordeos, A.C.; Zeigler, R.S. Effects and possible causes of an unprecedented rice blast epidemic on the traditional farming system of Bhutan. Agric. Ecosyst. Environ. 2000, 78, 237–248. [Google Scholar] [CrossRef]
- Prabhu, A.S.; Filippi, M.C.; Castro, N. Pathogenic variation among isolates of Pyricularia oryzae affecting rice, wheat, and grasses in Brazil. Int. J. Pest Manag. 1992, 38, 367–371. [Google Scholar]
- Hubert, J.; Mabagala, R.B.; Mamiro, D.P. Efficacy of selected plant extracts against Pyricularia grisea, causal agent of rice blast disease. Am. J. Plant Sci. 2015, 6, 602–611. [Google Scholar] [CrossRef]
- Gelderblom, W.C.; Jaskiewicz, K.; Marasas, W.F.; Thiel, P.G.; Horak, R.M.; Vleggaar, R.; Kriek, N.P. Fumonisins—Novel mycotoxins with cancer-promoting activity produced by Fusarium moniliforme. Appl. Environ. Microbiol. 1988, 54, 1806–1811. [Google Scholar] [PubMed]
- Madden, L.; Nutter, J.F. Modeling crop losses at the field scale. Can. J. Plant Pathol. 1995, 17, 124–137. [Google Scholar] [CrossRef]
- Desjardins, A.E.; Plattner, R.D.; Nelsen, T.C.; Leslie, J.F. Genetic analysis of fumonisin production and virulence of Gibberella fujikuroi mating population A (Fusarium moniliforme) on maize (Zea mays) seedlings. Appl. Environ. Microbiol. 1995, 61, 79–86. [Google Scholar] [PubMed]
- Luo, D.Q.; Shao, H.J.; Zhu, H.J.; Liu, J.K. Activity in vitro and in vivo against plant pathogenic fungi of grifolin isolated from the basidiomycete Albatrellus dispansus. Z. Naturforsch. C J. Biosci. 2005, 60, 50–56. [Google Scholar] [CrossRef]
- Lam, S.; Ng, T. Hypsin, a novel thermostable ribosome-inactivating protein with antifungal and antiproliferative activities from fruiting bodies of the edible mushroom Hypsizigus marmoreus. Biochem. Biophys. Res. Commun. 2001, 285, 1071–1075. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.Q.; Wang, F.; Bian, X.Y.; Liu, J.K. Rufuslactone, a new antifungal sesquiterpene from the fruiting bodies of the basidiomycete Lactarius rufus. J. Antibiot. 2005, 58, 456–459. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.H.; Ng, T.B.; Wang, H.; Sze, S.C.; Zhang, K.Y.; Li, Q.; Lu, X. Cordymin, an antifungal peptide from the medicinal fungus Cordyceps militaris. Phytomedicine 2011, 18, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Heleno, S.A.; Ferreira, I.C.; Esteves, A.P.; Ćirić, A.; Glamočlija, J.; Martins, A.; Soković, M.; Queiroz, M.J. Antimicrobial and demelanizing activity of Ganoderma lucidum extract, p-hydroxybenzoic and cinnamic acids and their synthetic acetylated glucuronide methyl esters. Food Chem. Toxicol. 2013, 58, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Ngai, P.H.; Zhao, Z.; Ng, T. Agrocybin, an antifungal peptide from the edible mushroom Agrocybe cylindracea. Peptides 2005, 26, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Ngai, P.H.; Ng, T. Lentin, a novel and potent antifungal protein from shitake mushroom with inhibitory effects on activity of human immunodeficiency virus-1 reverse transcriptase and proliferation of leukemia cells. Life Sci. 2003, 73, 3363–3374. [Google Scholar] [CrossRef] [PubMed]
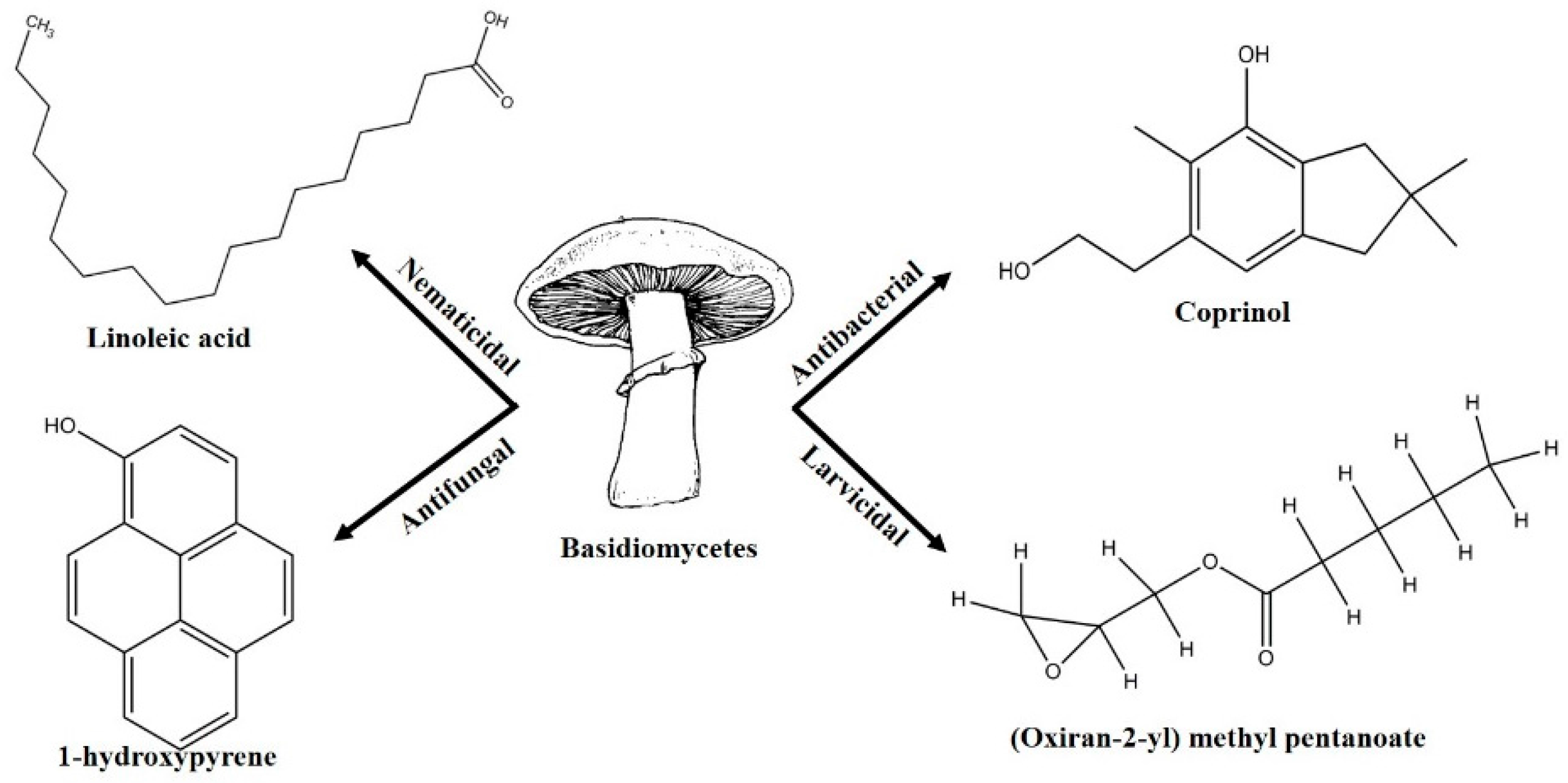
- Lambert, M.; Kremer, S.; Anke, H. Antimicrobial, phytotoxic, nematicidal, cytotoxic, and mutagenic activities of 1-hydroxypyrene, the initial metabolite in pyrene metabolism by the basidiomycete Crinipellis stipitaria. Bull. Environ. Contam. Toxicol. 1995, 55, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.I.; Yun, B.S.; Kim, Y.K.; Kwon, B.M.; Kim, H.G.; Lee, H.B.; Jeong, W.J.; Kim, S.U. Phellinsin A, a novel chitin synthases inhibitor produced by Phellinus sp. PL3. J. Antibiot. 2000, 53, 903–911. [Google Scholar] [CrossRef] [PubMed]
- Robles-Hernández, L.; Gonzalez-Franco, A.C.; Soto-Parra, J.M.; Montes-Domínguez, F. Review of agricultural and medicinal applications of basidiomycete mushrooms. Tecnociencia Chihuah. 2008, 2, 95–107. [Google Scholar]
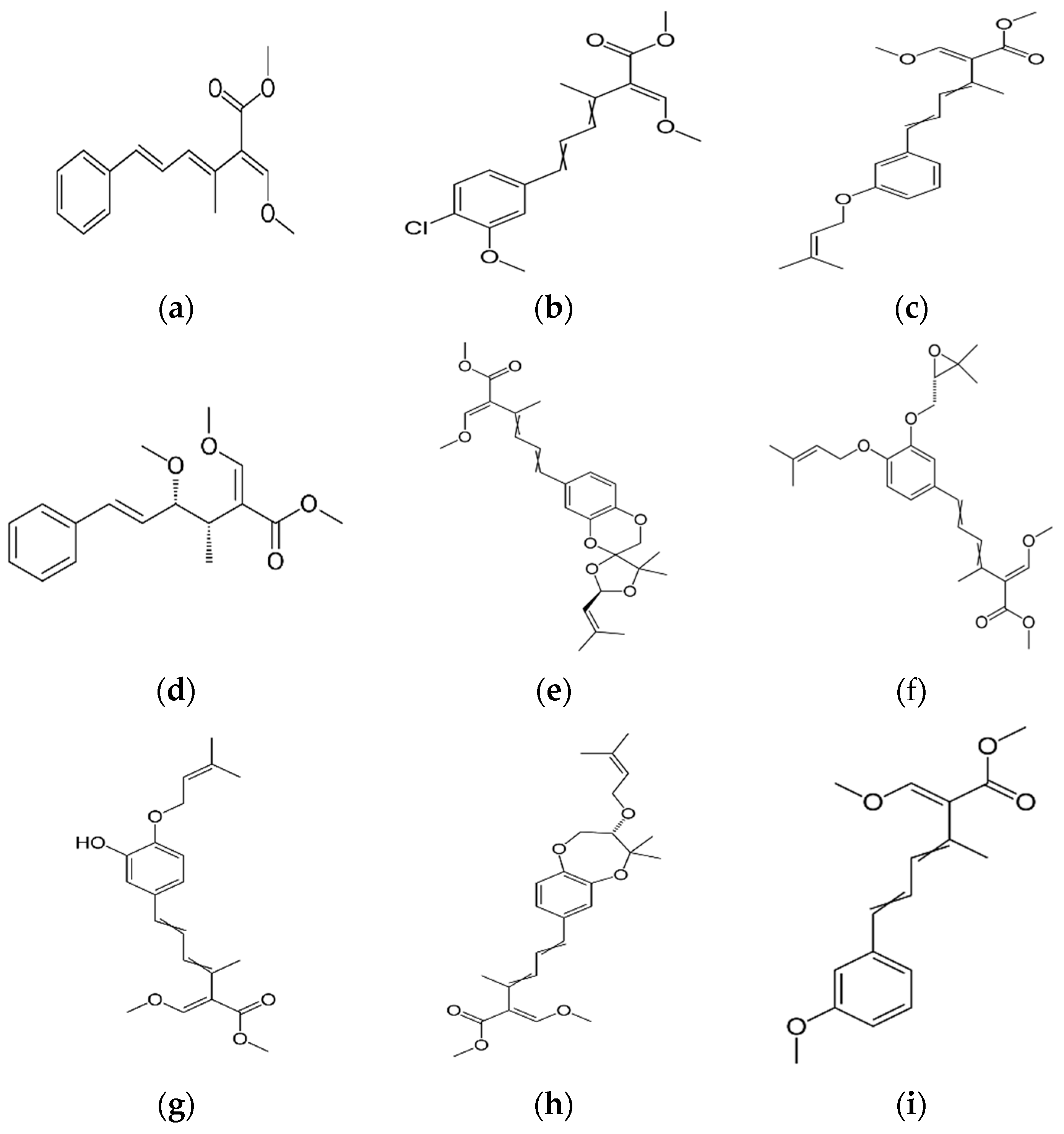
- Anke, T.; Oberwinkler, F.; Steglich, W.; Schramm, G. The strobilurins-new antifungal antibiotics from the basidiomycete Strobilurus tenacellus. J. Antibiot. 1977, 30, 806–810. [Google Scholar] [CrossRef] [PubMed]
- Weber, W.; Anke, T.; Steffan, B.; Steglich, W. Antibiotics from basidiomycetes. XXXII. Strobilurin E: A new cytostatic and antifungal (E)-BETA-methoxyacrylate antibiotic from Crepidotus fulvotomentosus Peck. J. Antibiot. 1990, 43, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Fredenhagen, A.; Kuhn, A.; Peter, H.H.; Cuomo, V.; Giuliano, U. Strobilurins F, G and H, three new antifungal metabolites from Bolinea lutea. I. Fermentation, isolation and biological activity. J. Antibiot. 1990, 43, 655–660. [Google Scholar] [CrossRef] [PubMed]
- Gullino, M.L.; Leroux, P.; Smith, C.M. Uses and challenges of novel compounds for plant disease control. Crop Prot. 2000, 19, 1–11. [Google Scholar] [CrossRef]
- Anke, T.; Besl, H.; Mocek, U.; Steglich, W. Antibiotics from basidiomycetes. XVIII. Strobilurin C and Oudemansin B, two new antifungal metabolites from Xerula species (Agaricales). J. Antibiot. 1983, 36, 661–666. [Google Scholar] [CrossRef] [PubMed]
- Imtiaj, A.; Jayasinghe, C.; Lee, G.W.; Lee, T.S. Antibacterial and antifungal activities of Stereum ostrea, an inedible wild mushroom. Mycobiology 2007, 35, 210–214. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Bao, L.; Yang, X.; Li, L.; Li, S.; Gao, H.; Yao, X.S.; Wen, H.; Liu, H.W. Bioactive sesquiterpenoids from the solid culture of the edible mushroom Flammulina velutipes growing on cooked rice. Food Chem. 2012, 132, 1346–1353. [Google Scholar] [CrossRef]
- Fu, L.Q.; Guo, X.S.; Liu, X.; He, H.L.; Wang, Y.L.; Yang, Y.S. Synthesis and antibacterial activity of C-2 (S)-substituted pleuromutilin derivatives. Chin. Chem. Lett. 2010, 21, 507–510. [Google Scholar] [CrossRef]
- Saddiqe, Z.; Naeem, I.; Maimoona, A. A review of the antibacterial activity of Hypericum perforatum L. J. Ethnopharmacol. 2010, 131, 511–521. [Google Scholar] [CrossRef] [PubMed]
- Barros, L.; Baptista, P.; Estevinho, L.M.; Ferreira, I.C. Effect of fruiting body maturity stage on chemical composition and antimicrobial activity of Lactarius sp. mushrooms. J. Agric. Food. Chem. 2007, 55, 8766–8771. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.J.; Ferreira, I.C.; Dias, J.; Teixeira, V.; Martins, A.; Pintado, M. A review on antimicrobial activity of mushroom (Basidiomycetes) extracts and isolated compounds. Planta Med. 2012, 78, 1707–1718. [Google Scholar] [CrossRef] [PubMed]
- Erjavec, J.; Ravnikar, M.; Brzin, J.; Grebenc, T.; Blejec, A.; Gosak, M.Ž.; Sabotič, J.; Kos, J.; Dreo, T. Antibacterial activity of wild mushroom extracts on bacterial wilt pathogen Ralstonia solanacearum. Plant Dis. 2016, 100, 453–464. [Google Scholar] [CrossRef]
- Dreo, T.; Zelko, M.; Skubic, J.; Brzin, J.; Ravnikar, M. Antibacterial activity of proteinaceous extracts of higher basidiomycetes mushrooms against plant pathogenic bacteria. Int. J. Med. Mushrooms 2007, 9, 226–237. [Google Scholar]
- Herms, S.; Seehaus, K.; Koehle, H.; Conrath, U. A strobilurin fungicide enhances the resistance of tobacco against tobacco mosaic virus and Pseudomonas syringae pvtabaci. Plant Physiol. 2002, 130, 120–127. [Google Scholar] [CrossRef] [PubMed]
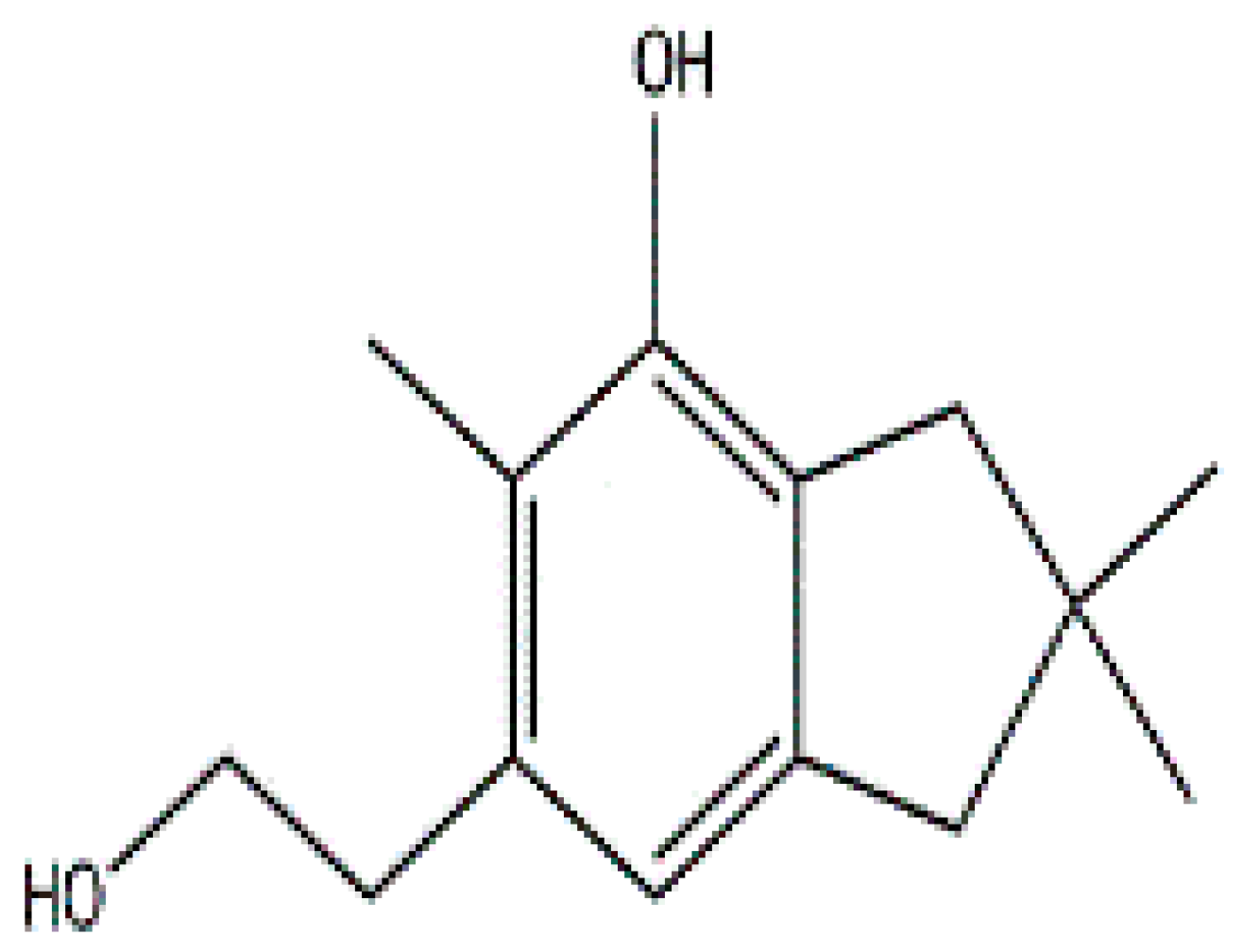
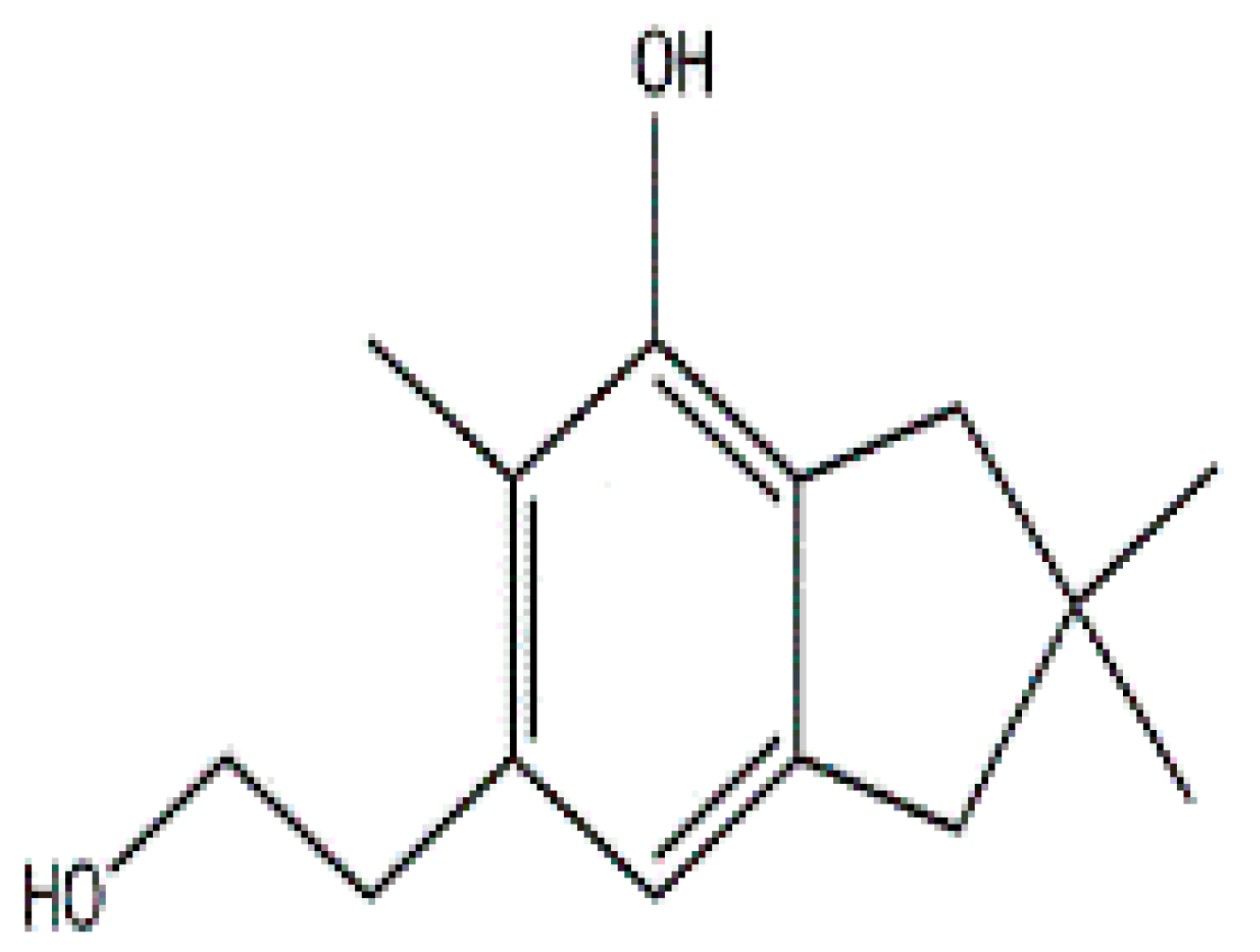
- Johansson, M.; Sterner, O.; Labischinski, H.; Anke, T. Coprinol, a new antibiotic cuparane from a Coprinus species. Z. Naturforsch. C J. Biosci. 2001, 56, 31–34. [Google Scholar] [CrossRef]
- Brandt, C.R.; Piraino, F. Mushroom antivirals. Recent Res. Dev. Antimicrob. Agents Chemother. 2000, 4, 11–26. [Google Scholar]
- Gao, Y.; Zhou, S.; Huang, M.; Xu, A. Antibacterial and antiviral value of the genus Ganoderma P. Karst. species (Aphyllophoromycetideae): A review. Int. J. Med. Mushrooms 2003, 5. [Google Scholar] [CrossRef]
- Kovalenko, O.; Polishchuk, O.; Krupodorova, T. Screening of metabolites produced by strains of Ganoderma lucidum [Curt.: Fr] P. Karst and Ganoderma applanatum [Pirs.: Waller] Pat. for their activity against tobacco mosaic virus. Bull. Tara Shevchenko Nat. Univ. Kyiv. Ser. Biol. 2008, 51, 32–34. [Google Scholar]
- Zjawiony, J.K. Biologically Active Compounds from Aphyllophorales (Polypore) Fungi. J. Nat. Prod. 2004, 67, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Di Piero, R.M.; Novaes, Q.S.; Pascholati, S.F. Effect of Agaricus brasiliensis and Lentinula edodes mushrooms on the infection of passionflower with Cowpea aphid-borne mosaic virus. Braz. Arch. Biol. Technol. 2010, 53, 269–278. [Google Scholar] [CrossRef]
- Kovalenko, O.G.; Polishchuk, O.N.; Wasser, S.P. Virus Resistance induced by glucuronoxylomannan isolated from submerged cultivated yeast-like cell biomass of medicinal yellow brain mushroom Tremella mesenterica Ritz.: Fr.(Heterobasidiomycetes) in hypersensitive host plants. Int. J. Med. Mushrooms 2009, 11, 199–205. [Google Scholar] [CrossRef]
- Tavantzis, S.; Smith, S. Isolation and evaluation of a plant-virus-inhibiting quinone from sporophores of Agaricus bisporus. Phytopathology 1982, 72, 619–621. [Google Scholar] [CrossRef]
- Kakuta, M.; Sone, Y.; Umeda, T.; Misaki, A. Comparative structural studies on acidic heteropolysaccharides isolated from “Shirokikurage” fruit body of Tremella fuciformis Berk, and the growing culture of its yeast-like cells. Agric. Biol. Chem. 1979, 43, 1659–1668. [Google Scholar] [CrossRef]
- Kovalenko, A. Protein-carbohydrate interactions in the realization of plant resistance to viruses. Mikrobiol. Zhurnal 1993, 55, 74–91. [Google Scholar]
- Vinogradov, E.; Petersen, B.O.; Duus, J.Ø.; Wasser, S. The structure of the glucuronoxylomannan produced by culinary-medicinal yellow brain mushroom (Tremella mesenterica Ritz.: Fr., Heterobasidiomycetes) grown as one cell biomass in submerged culture. Carbohydr. Res. 2004, 339, 1483–1489. [Google Scholar] [CrossRef] [PubMed]
- Haydock, P.P.; Woods, S.R.; Grove, I.G.; Hare, M.C.; Perry, R.N.; Moens, M. Chemical control of nematodes. In Plant Nematology; CABI Publishing: Oxfordshire, UK, 2006; pp. 392–410. [Google Scholar]
- Haydock, P.P.; Woods, S.R.; Grove, I.G.; Hare, M.C.; Perry, R.N.; Moens, M. Chemical control of nematodes. In Plant Nematology, 2nd ed.; CABI Publishing: Oxfordshire, UK, 2013; pp. 459–479. [Google Scholar]
- Soltani, T.; Nejad, R.F.; Ahmadi, A.R.; Fayazi, F. Chemical control of root-knot nematode (Meloidogyne javanica) on olive in the greenhouse conditions. J. Plant Pathol. Microbiol. 2013, 2013. [Google Scholar] [CrossRef]
- Comans-Pérez, R.; Aguilar-Marcelino, L.; Mendoza de Gives, P.; Sánchez, J.; López-Arellano, M.; Singh, M. In vitro lethal capability of ten strains of edible mushrooms against Haemonchus contortus (Nematoda) infective larvae [conference poster]. In Proceedings of the 8th International Conference on Mushroom Biology and Mushroom Products (ICMBMP8), New Delhi, India, 19–22 November 2014; Volume I & II, pp. 557–562.
- Luo, H.; Li, X.; Li, G.; Pan, Y.; Zhang, K. Acanthocytes of Stropharia rugosoannulata function as a nematode-attacking device. Appl. Environ. Microbiol. 2006, 72, 2982–2987. [Google Scholar] [CrossRef] [PubMed]
- Kwok, O.; Plattner, R.; Weisleder, D.; Wicklow, D. A nematicidal toxin from Pleurotus ostreatus NRRL 3526. J. Chem. Ecol. 1992, 18, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Palizi, P.; Goltapeh, E.; Pourjam, E.; Safaie, N. Potential of oyster mushrooms for the biocontrol of sugar beet nematode (Heterodera schachtii). J. Plant Prot. Res. 2009, 49, 27–34. [Google Scholar] [CrossRef]
- Mamiy, Y. Attraction of the pinewood nematode to mycelium of some wood-decay fungi. Jpn. J. Nematol. 2006, 36, 1–9. [Google Scholar] [CrossRef]
- Bua-art, S.; Saksirirat, W.; Hiransalee, A.; Kanokmedhaku, S.; Lekphrom, R. Effect of bioactive compound from luminescent mushroom (Neonothopanus nambi Speg.) on root-knot nematode (Meloidogyne incognita Chitwood) and non-target organisms. KKU Res. J. 2011, 16, 331–341. [Google Scholar]
- De Jong, E.; Cazemier, A.E.; Field, J.A.; de Bont, J.A. Physiological role of chlorinated aryl alcohols biosynthesized de novo by the white rot fungus Bjerkandera sp. strain BOS55. Appl. Environ. Microbiol. 1994, 60, 271–277. [Google Scholar] [PubMed]
- Stadler, M.; Mayer, A.; Anke, H.; Sterner, O. Fatty acids and other compounds with nematicidal activity from cultures of Basidiomycetes. Planta Med. 1994, 60, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Lorenzen, K.; Anke, T. Basidiomycetes as source for new bioactive natural products. Curr. Org. Chem. 1998, 2, 329–364. [Google Scholar]
- Hayashi, M.; Wada, K.; Munakata, K. New nematicidal metabolites from a fungus, Irpex lacteus. Agric. Biol. Chem. 1981, 45, 1527–1529. [Google Scholar]
- Mayer, A.; Anke, H.; Sterner, O. Omphalotin, a new cyclic peptide with potent nematicidal activity from Omphalotus olearius I. Fermentation and biological activity. Nat. Prod. Lett. 1997, 10, 25–32. [Google Scholar] [CrossRef]
- Dong, J.Y.; Li, X.P.; Li, L.; Li, G.H.; Liu, Y.J.; Zhang, K.Q. Preliminary results on nematicidal activity from culture filtrates of Basidiomycetes against the pine wood nematode, Bursaphelenchus xylophilus (Aphelenchoididae). Ann. Microbiol. 2006, 56, 163–166. [Google Scholar] [CrossRef]
- Njogu, E.M.; Njue, A.; Omolo, J.O.; Cheplogoi, P.K. Larvicidal activity of (oxiran-2-yl) methylpentanoate extracted from mushroom Cyptotrama asprata against mosquito Aedes aegypti. Int. J. Biol. Chem. Sci. 2009, 3. [Google Scholar] [CrossRef]
- Bucker, A.; Bucker, N.C.; Souza, A.Q.; Gama, A.M.; Rodrigues-Filho, E.; Costa, F.M.; Tadei, W.P. Larvicidal effects of endophytic and basidiomycete fungus extracts on Aedes and Anopheles larvae (Diptera, Culicidae). Rev. Soc. Bras. Med. Trop. 2013, 46, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Chelela, B.L.; Chacha, M.; Matemu, A. Larvicidal potential of wild mushroom extracts against Culex quinquefasciatus. Am. J. Res. Commun. 2014, 2, 105–114. [Google Scholar]
- Baraza, L.; Joseph, C.; Moshi, M.; Nkunya, M. Chemical constituents and biological activity of three Tanzanian wild mushroom species. Tanzan. J. Sci. 2007, 33. [Google Scholar] [CrossRef]
- Olayemi, I.K. Evaluation of Larvicidal efficacy of extract of the fungus Ganoderma lucidum, for the control of the filarial vector mosquito, Culex pipiens pipiens (Diptera: Culicidae). Am. J. Drug Discov. Dev. 2013, 3, 130–139. [Google Scholar] [CrossRef]
- Chirchir, D.K. Isolation and Purification of Mosquito Larvicidal Compounds from Extracts of a Basidiomycete jo5289. Master’s Thesis, Egerton University, Njoro, Kenya, February 2010. [Google Scholar]
- Thongwat, D.; Pimolsri, U.; Somboon, P. Screening for mosquito larvicidal activity of thai mushroom extracts with special reference to Steccherinum sp against Aedes aegypti (L.)(Diptera: Culicidae). Southeast Asian J. Trop. Med. Public Health 2015, 46, 586–595. [Google Scholar] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
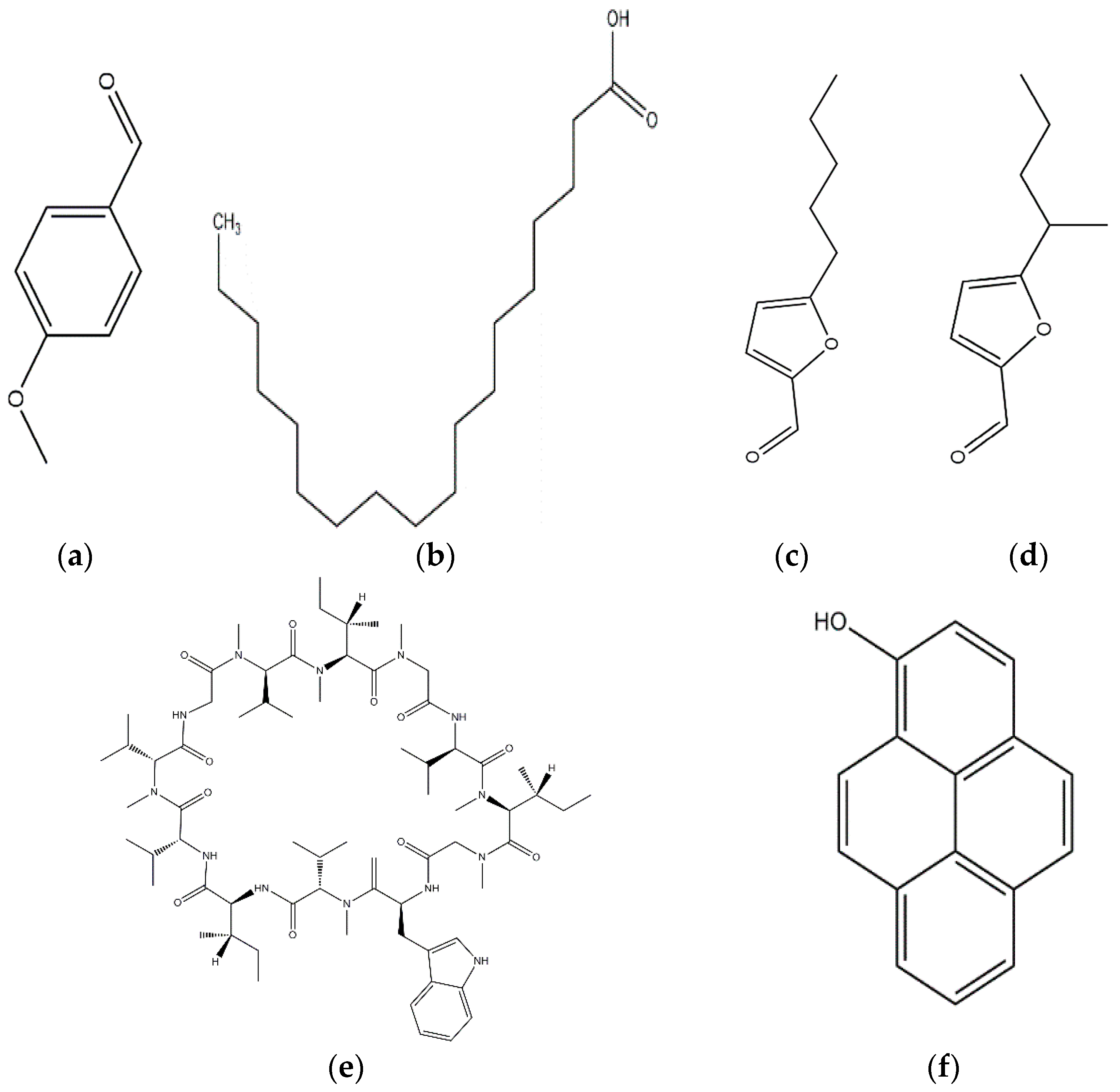
| Active Compound | Compound Structure | Target Pathogen | Reference |
|---|---|---|---|
| Ganodermin (Ganoderma lucidum) Antifungal protein | NA | Physalospora piricola; Botytis cinerea | [19] |
| Pleurostrin (Pleurotus ostreatus) Antifungal protein | NA | Botryosphaeria berengeriana | [20] |
| Eryngin (Pleurotus eryngii) Antifungal protein | NA | Mycosphaerella arachidicola | [20,23] |
| Antifungal protein mLAP (Lyophyllum shimeji) | NA | Physalospora piricola | [22] |
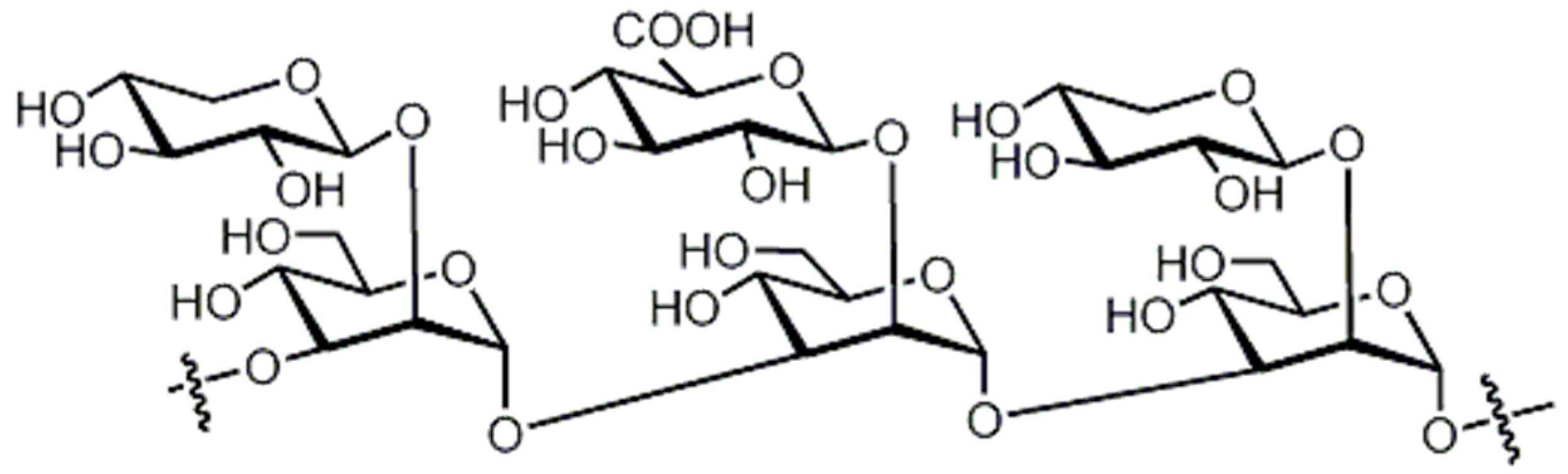
| Lyophyllin (Lyophyllus shimeji) | 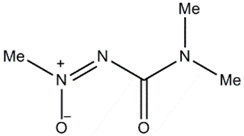 | Physalospora piricola | [22] |
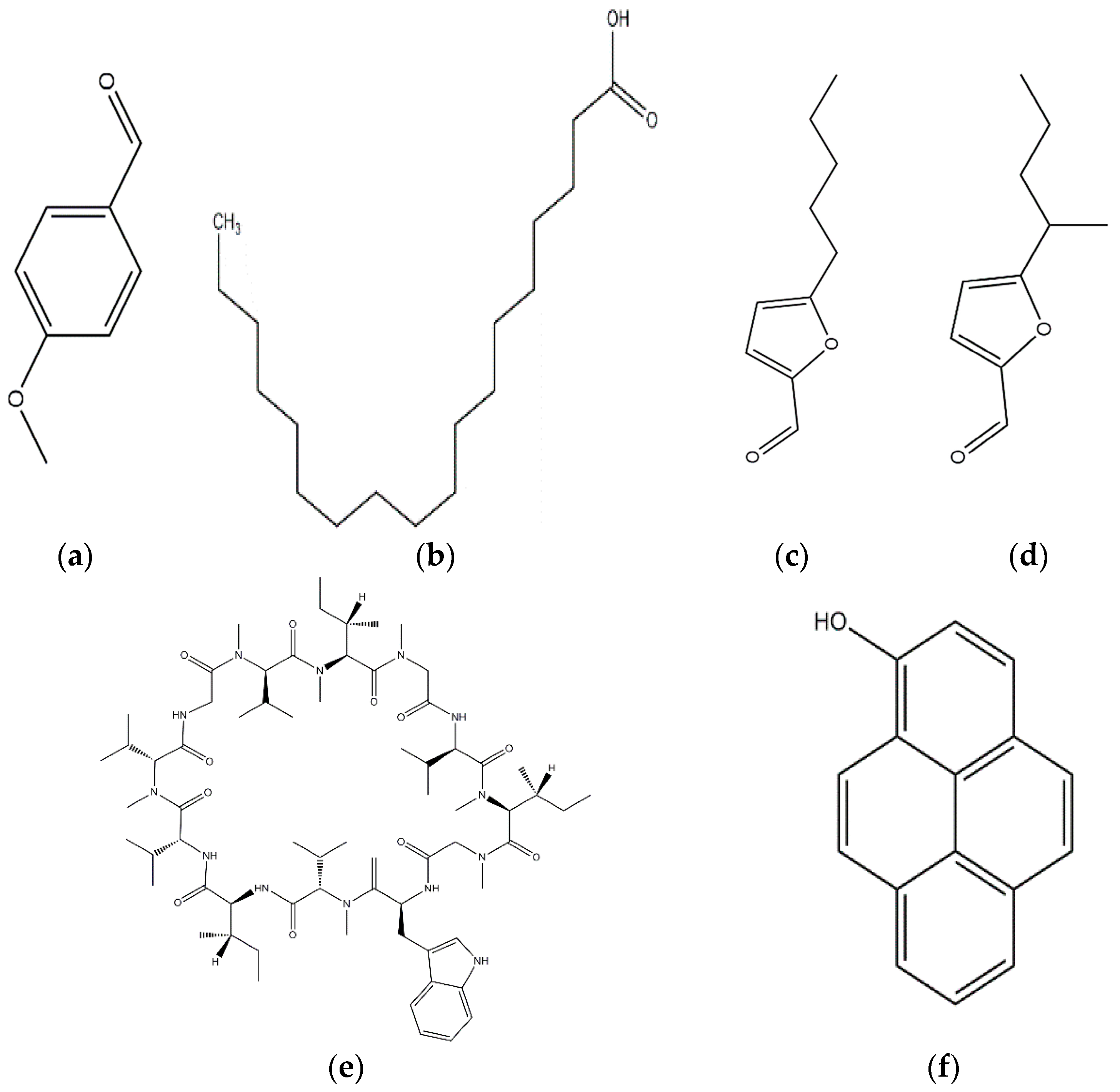
| Grifoline (Albatrellus dispansus) | 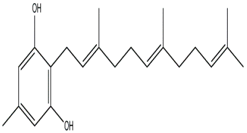 | Alternaria alternate; Pyricularia oryzae; Rhizoctonia solani; Sclerotina sclerotiorum; Fusarium graminearum; Botytis cinerea; Gaeumannomyces graminis; Gloesporium fructigenum | [46] |
| Hypsin (Hypsizigus marmoreus) Antifungal protein | NA | Botryosphaeria berengeriana; Botytis cinerea; Mycosphaerella arachidicola | [47] |
| Rufuslactone (Lactarius rufus) | 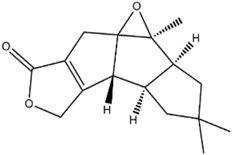 | Alternaria alternata; Fusarium graminearum; Botytis cinerea; Alternaria brassicae | [48] |
| Cordymin (Cordyceps militaris) Antifungal protein | NA | Rhizoctonia solani; Mycosphaerella arachidicola; Bipolaris maydis | [49] |
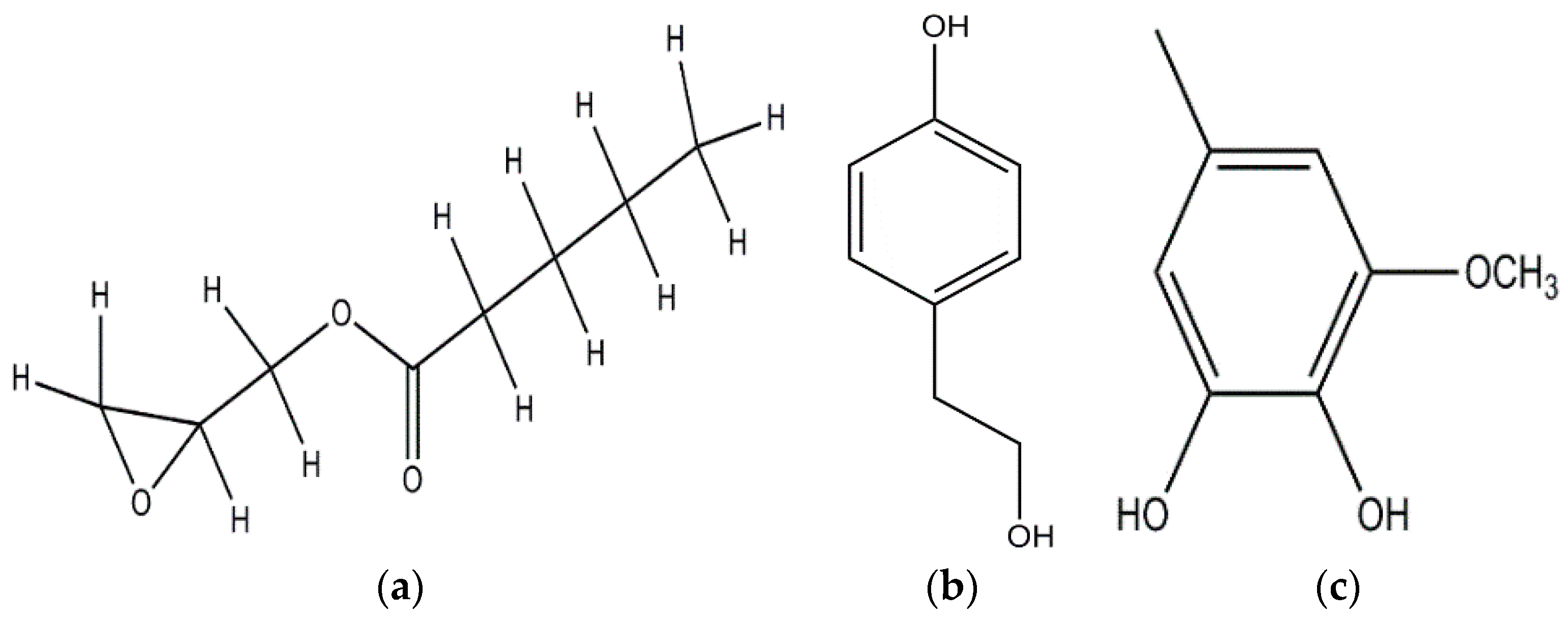
| p-Hydroxybenzoic and Cinnamic acids (Ganoderma lucidum) | 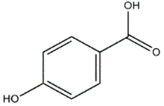 | Trichoderma viride; Penicillium ochrochloron; P. funiculosum | [50] |
| p-Hydroxybenzoic | |||
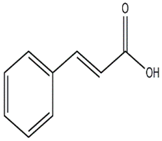 | |||
| Cinnamic acids | |||
| Chrysotrione A and B (Hygrophorus chrysodon) | 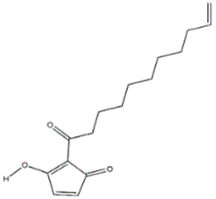 | Fusarium verticillioides | [50] |
| Chrysotrione A | |||
 | |||
| Chrysotrione B | |||
| Agrocybin (Agrocybe cylindracea) | 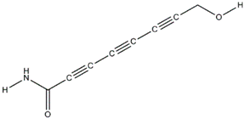 | Mycosphaerella arachidicola | [51] |
| Lentin (Lentinus edodes) Antifungal protein | NA | Mycosphaerella arachidicola | [52] |
| 1-hydroxypyrene (Cordyceps militaris) | 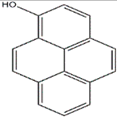 | Fusarium oxysporum | [53] |
| Phellinsin A (Phellinus sp.) | 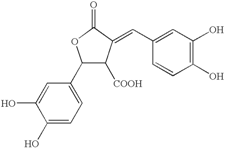 | Gloeosporium orbiculare; Pyricularia grisea | [54] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sivanandhan, S.; Khusro, A.; Paulraj, M.G.; Ignacimuthu, S.; AL-Dhabi, N.A. Biocontrol Properties of Basidiomycetes: An Overview. J. Fungi 2017, 3, 2. https://doi.org/10.3390/jof3010002
Sivanandhan S, Khusro A, Paulraj MG, Ignacimuthu S, AL-Dhabi NA. Biocontrol Properties of Basidiomycetes: An Overview. Journal of Fungi. 2017; 3(1):2. https://doi.org/10.3390/jof3010002
Chicago/Turabian StyleSivanandhan, Subramaniyan, Ameer Khusro, Michael Gabriel Paulraj, Savarimuthu Ignacimuthu, and Naif Abdullah AL-Dhabi. 2017. "Biocontrol Properties of Basidiomycetes: An Overview" Journal of Fungi 3, no. 1: 2. https://doi.org/10.3390/jof3010002
APA StyleSivanandhan, S., Khusro, A., Paulraj, M. G., Ignacimuthu, S., & AL-Dhabi, N. A. (2017). Biocontrol Properties of Basidiomycetes: An Overview. Journal of Fungi, 3(1), 2. https://doi.org/10.3390/jof3010002