
Multiple Analyses Reveal Evidence for Three New Species of Collybia (Clitocybaceae, Basidiomycete) from China

Abstract
1. Introduction
2. Materials and Methods
2.1. Specimens and Morphological Studies
2.2. DNA Extraction, PCR, and Sequencing
2.3. Phylogenetic Analyses
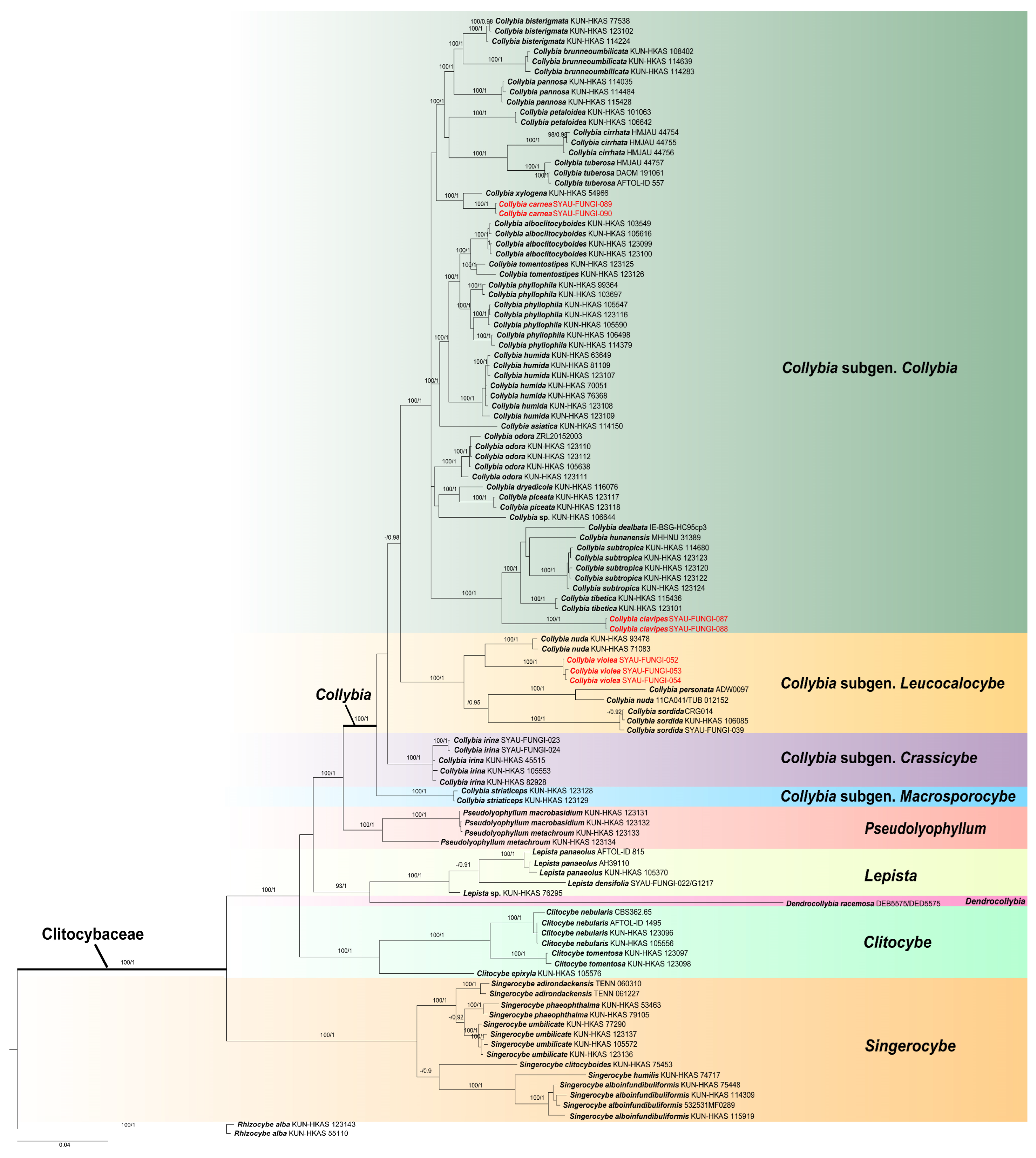
3. Results
3.1. Phylogenetic Analyses
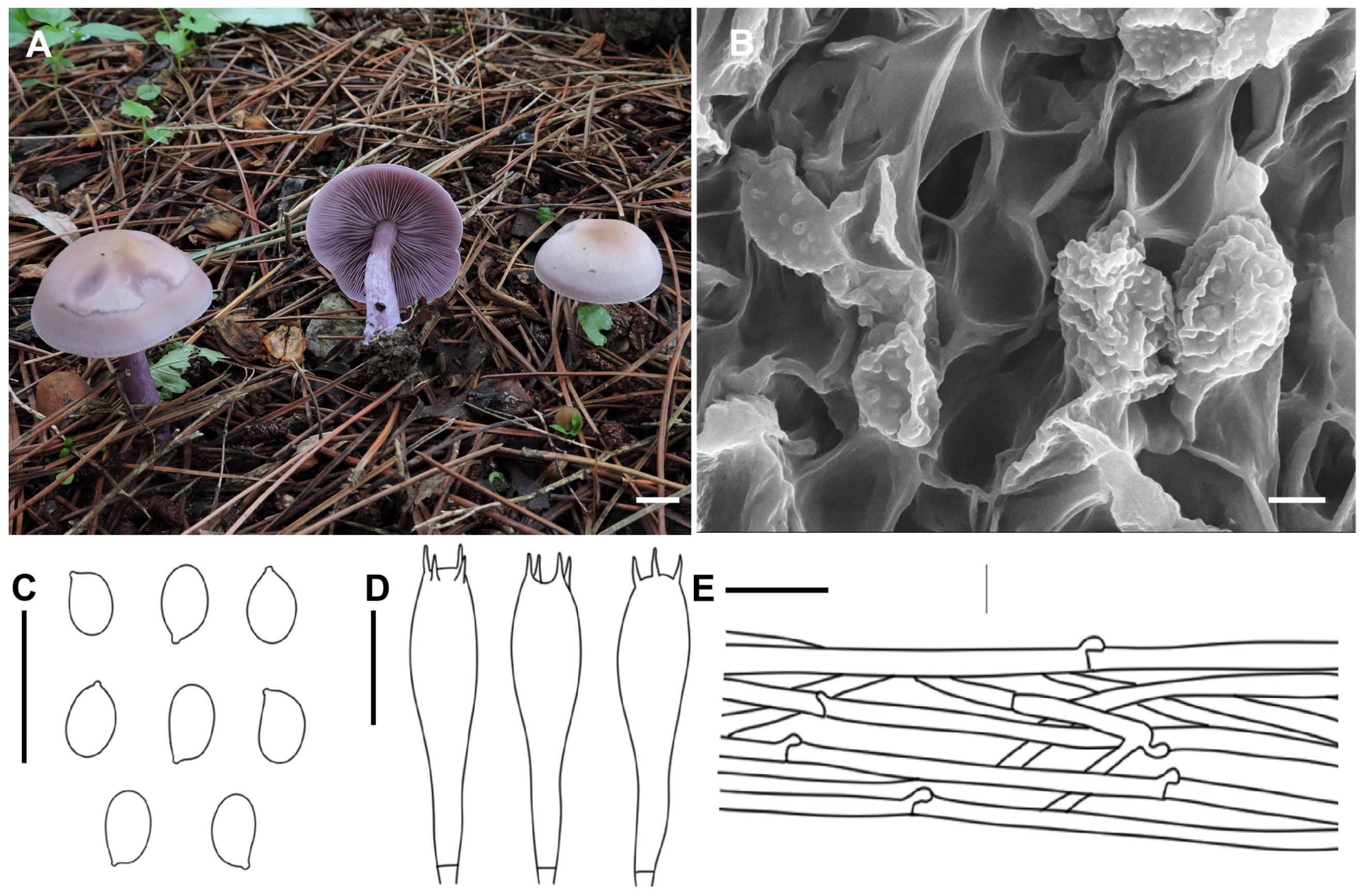
3.2. Taxonomy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Matheny, P.B.; Hofstetter, V.; Aime, M.C.; Moncalvo, J.M.; Ge, Z.M.; Yang, Z.L.; Slot, J.C.; Ammirati, J.F.; Baroni, T.J.; Bougher, N.L.; et al. Major clades of Agaricales: A multilocus phylogenetic overview. Mycologia 2006, 98, 982–995. [Google Scholar] [CrossRef] [PubMed]
- Ammirati, J.F.; Parker, A.D.; Matheny, P.B. Cleistocybe, a new genus of Agaricales. Mycoscience 2007, 48, 282–289. [Google Scholar] [CrossRef]
- Vizzini, A.; Musumeci, E.; Murat, C. Trichocybe, a new genus for Clitocybe puberula (Agaricomycetes, Agaricales). Fungal Divers. 2010, 42, 97–105. [Google Scholar] [CrossRef]
- Alvarado, P.; Moreno, G.; Vizzini, A.; Consiglio, G.; Manjón, J.L.; Setti, L. Atractosporocybe, Leucocybe and Rhizocybe: Three new clitocyboid genera in the Tricholomatoid clade (Agaricales) with notes on Clitocybe and Lepista. Mycologia 2015, 107, 123–136. [Google Scholar] [CrossRef]
- Matheny, P.B.; Hughes, K.W.; Kalichman, J.; Lebeuf, R. Pulverulina, a new genus of Agaricales for Clitocybe ulmicola. Southeast. Nat. 2020, 19, 447–459. [Google Scholar] [CrossRef]
- He, Z.M.; Yang, Z.L. A new clitocyboid genus Spodocybe and a new subfamily Cuphophylloideae in the family Hygrophoraceae (Agaricales). MycoKeys 2021, 79, 129–148. [Google Scholar] [CrossRef] [PubMed]
- Moncalvo, J.M.; Vilgalys, R.; Redhead, S.A.; Johnson, J.E.; James, T.Y.; Catherine Aime, M.; Hofstetter, V.; Verduin, S.J.; Larsson, E.; Baroni, T.J.; et al. One hundred and seventeen clades of euagarics. Mol. Phylogenetics Evol. 2002, 23, 357–400. [Google Scholar] [CrossRef]
- Walther, G.; Garnica, S.; Weiß, M. The systematic relevance of conidiogenesis modes in the gilled Agaricales. Mycol. Res. 2005, 109, 525–544. [Google Scholar] [CrossRef]
- Binder, M.; Larsson, K.H.; Matheny, P.B.; Hibbett, D.S. Amylocorticiales ord. nov. and Jaapiales ord. nov.: Early diverging clades of agaricomycetidae dominated by corticioid forms. Mycologia 2010, 102, 865–880. [Google Scholar] [CrossRef]
- Sánchez-García, M.; Matheny, P.B.; Palfner, G.; Lodge, D.J. Deconstructing the Tricholomataceae (Agaricales) and introduction of the new genera Albomagister, Corneriella, Pogonoloma and Pseudotricholoma. Taxonomy 2014, 63, 993–1007. [Google Scholar] [CrossRef]
- Alvarado, P.; Moreau, P.A.; Sesli, E.; Khodja, L.Y.; Contu, M.; Vizzini, A. Phylogenetic studies on Bonomyces (Tricholomatineae, Agaricales) and two new combinations from Clitocybe. Cryptogam. Mycol. 2018, 39, 149–168. [Google Scholar] [CrossRef]
- Alvarado, P.; Moreau, P.A.; Dima, B.; Vizzini, A.; Consiglio, G.; Moreno, G.; Setti, L.; Kekki, T.; Huhtinen, S.; Liimatainen, K.; et al. Pseudoclitocybaceae fam. nov. (Agaricales, Tricholomatineae), a new arrangement at family, genus and species level. Fungal Divers. 2018, 90, 109–133. [Google Scholar] [CrossRef]
- He, Z.M.; Yang, Z.L. The genera Bonomyces, Harmajaea and Notholepista from Northwestern China: Two new species and a new record. Mycol. Prog. 2022, 21, 26. [Google Scholar] [CrossRef]
- He, Z.M.; Chen, Z.H.; Bau, T.; Wang, G.S.; Yang, Z.L. Systematic arrangement within the family Clitocybaceae (Tricholomatineae, Agaricales): Phylogenetic and phylogenomic evidence, morphological data and muscarine-producing innovation. Fungal Divers. 2023, 123, 1–47. [Google Scholar] [CrossRef]
- Qi, Y.; Yu, X.D.; Hou, J.X.; Guo, H.B.; Yang, R.H.; Xu, A.G. Addition to the genus Harmajaea (Agaricales, Pseudoclitocybaceae): A new and a known species from China. Phytotaxa 2024, 665, 157–166. [Google Scholar] [CrossRef]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour; Methuen and Co., Ltd.: London, UK, 1963; 242p. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes from phylogenetics. In PCR Protocols: Methods and Applications; Innes, M.A., Gelfand, D.H., Sninsky, J.S., White, T.J., Eds.; Academic Press: London, UK, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hestsr, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- Matheny, P.B. Improving phylogenetic inference of mushrooms with RPB1 and RPB2 nucleotide sequences (Inocybe; Agaricales). Mol. Phylogenetics Evol. 2005, 35, 1–20. [Google Scholar] [CrossRef]
- Matheny, P.B.; Wang, Z.; Binder, M.; Curtis, J.M.; Lim, Y.W.; Nilsson, H.; Hughes, K.W.; Hofstetter, V.; Ammirati, J.F.; Schoch, C.L.; et al. Contributions of rpb2 and tef1 to the phylogeny of mushrooms and allies (Basidiomycota, Fungi). Mol. Phylogenetics Evol. 2007, 43, 430–451. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Haeseler, A.V.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree v1.4.4: Tree Figure Drawing Tool. Available online: https://github.com/rambaut/figtree/releases (accessed on 18 September 2024).
- Harmaja, H. The genus Clitocybe (Agaricales) in Fennoscandia. Karstenia 1969, 10, 5–121. [Google Scholar] [CrossRef]
- Bigelow, H.E. North American species of Clitocybe; Part I; Strauss & Cramer GmbH: Vaduz, Liechtenstein, 1982. [Google Scholar]
- Bon, M. The Mushrooms and Toadstools of Britain and North-Western Europe; Hodder & Stoughton: London, UK, 1987. [Google Scholar]
- Breitenbach, J.; Kränzlin, F. Fungi of Switzerland, Vol. 3-Boletes and Agarics, 1st ed.; Mykologia Lucerne: Lucerne, Switzerland, 1991; 361p. [Google Scholar]
- Buczacki, S. Collins Fungi Guide: The Most Complete Field Guide to the Mushrooms and Toadstools of Britain & Europe; Harpercollins Pub Ltd.: New Delhi, India, 2014. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Voucher Numbers | GenBank Accession Numbers | Location | |||
|---|---|---|---|---|---|---|
| ITS | nrLSU | rpb2 | tef1-α | |||
| Collybia clavipes | SYAU-FUNGI-087 | PQ594916 | PQ594905 | PQ801137 | PQ801139 | Xizang, China |
| SYAU-FUNGI-088 | PQ594917 | PQ594906 | PQ801138 | PQ801140 | Xizang, China | |
| C. carnea | SYAU-FUNGI-089 | PQ594919 | PQ594908 | PQ609669 | PQ609675 | Xizang, China |
| SYAU-FUNGI-090 | PQ594920 | PQ594909 | PQ609670 | PQ609676 | Xizang, China | |
| C. violea | SYAU-FUNGI-052 | MT482103 | MT482097 | PQ817427 | MT510970 | Liaoning, China |
| SYAU-FUNGI-053 | MT482104 | MT482098 | PQ817428 | MT510971 | Liaoning, China | |
| SYAU-FUNGI-054 | MT482105 | MT482099 | PQ817429 | MT510972 | Liaoning, China | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qi, Y.; Xu, A.; Yang, L.; Guo, H.; Guo, Y.; Wan, F.; Yang, R.; Pei, Y.; Yu, X. Multiple Analyses Reveal Evidence for Three New Species of Collybia (Clitocybaceae, Basidiomycete) from China. J. Fungi 2025, 11, 371. https://doi.org/10.3390/jof11050371
Qi Y, Xu A, Yang L, Guo H, Guo Y, Wan F, Yang R, Pei Y, Yu X. Multiple Analyses Reveal Evidence for Three New Species of Collybia (Clitocybaceae, Basidiomycete) from China. Journal of Fungi. 2025; 11(5):371. https://doi.org/10.3390/jof11050371
Chicago/Turabian StyleQi, Yue, Aiguo Xu, Liu Yang, Hongbo Guo, Yaobin Guo, Fashuang Wan, Ruiheng Yang, Ying Pei, and Xiaodan Yu. 2025. "Multiple Analyses Reveal Evidence for Three New Species of Collybia (Clitocybaceae, Basidiomycete) from China" Journal of Fungi 11, no. 5: 371. https://doi.org/10.3390/jof11050371
APA StyleQi, Y., Xu, A., Yang, L., Guo, H., Guo, Y., Wan, F., Yang, R., Pei, Y., & Yu, X. (2025). Multiple Analyses Reveal Evidence for Three New Species of Collybia (Clitocybaceae, Basidiomycete) from China. Journal of Fungi, 11(5), 371. https://doi.org/10.3390/jof11050371