
Zinc Metalloprotease SlMEP1: An Essential Factor Required for Fungal Virulence in Stemphylium lycopersici

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Tomato Plants and Fungal Cultures
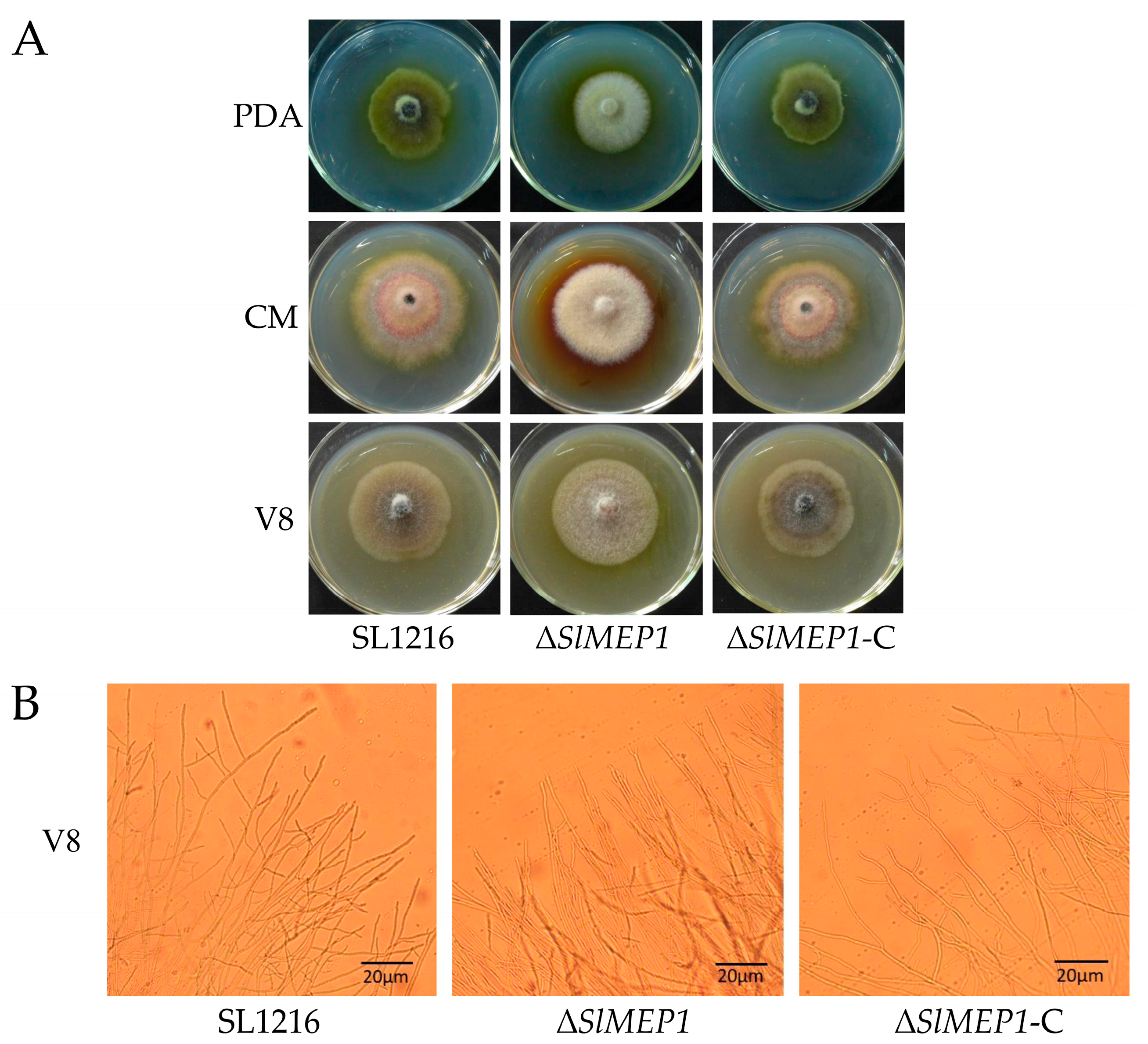
2.2. Vegetative Growth Assays and Observation of Mycelial Morphology
2.3. Phylogenetic Analysis and Sequence Alignment
2.4. Construction of Gene Deletion and Complementary Mutants
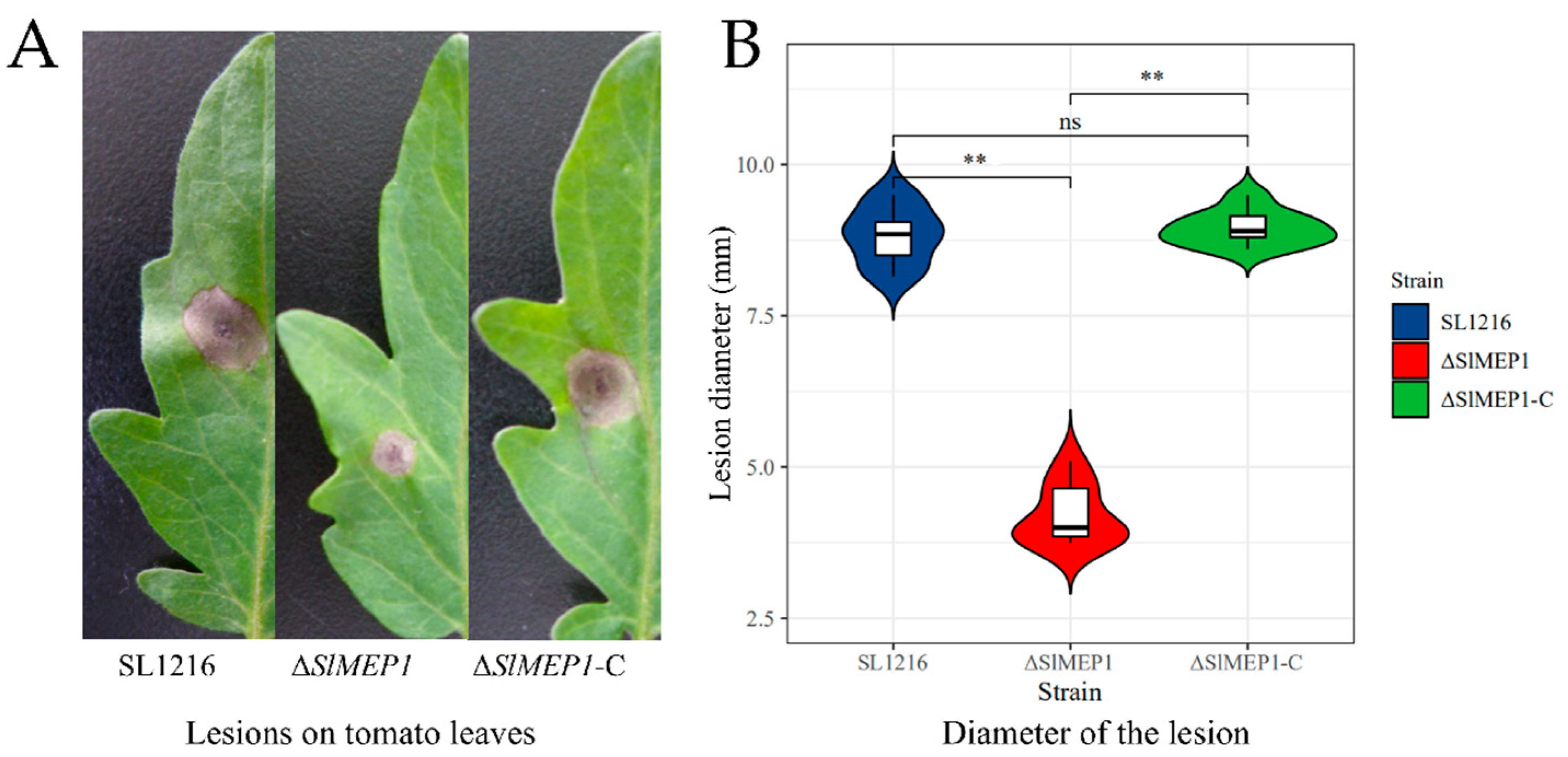
2.5. Pathogenicity Assays and Penetration Determination
2.6. Cell-Death-Inducing Activities and Disease Assays of Heterologously Expressed SlMEP1
2.7. RNA Extraction and cDNA Synthesis
2.8. Quantitative Reverse Transcription PCR Analysis of Gene Expressions
3. Results
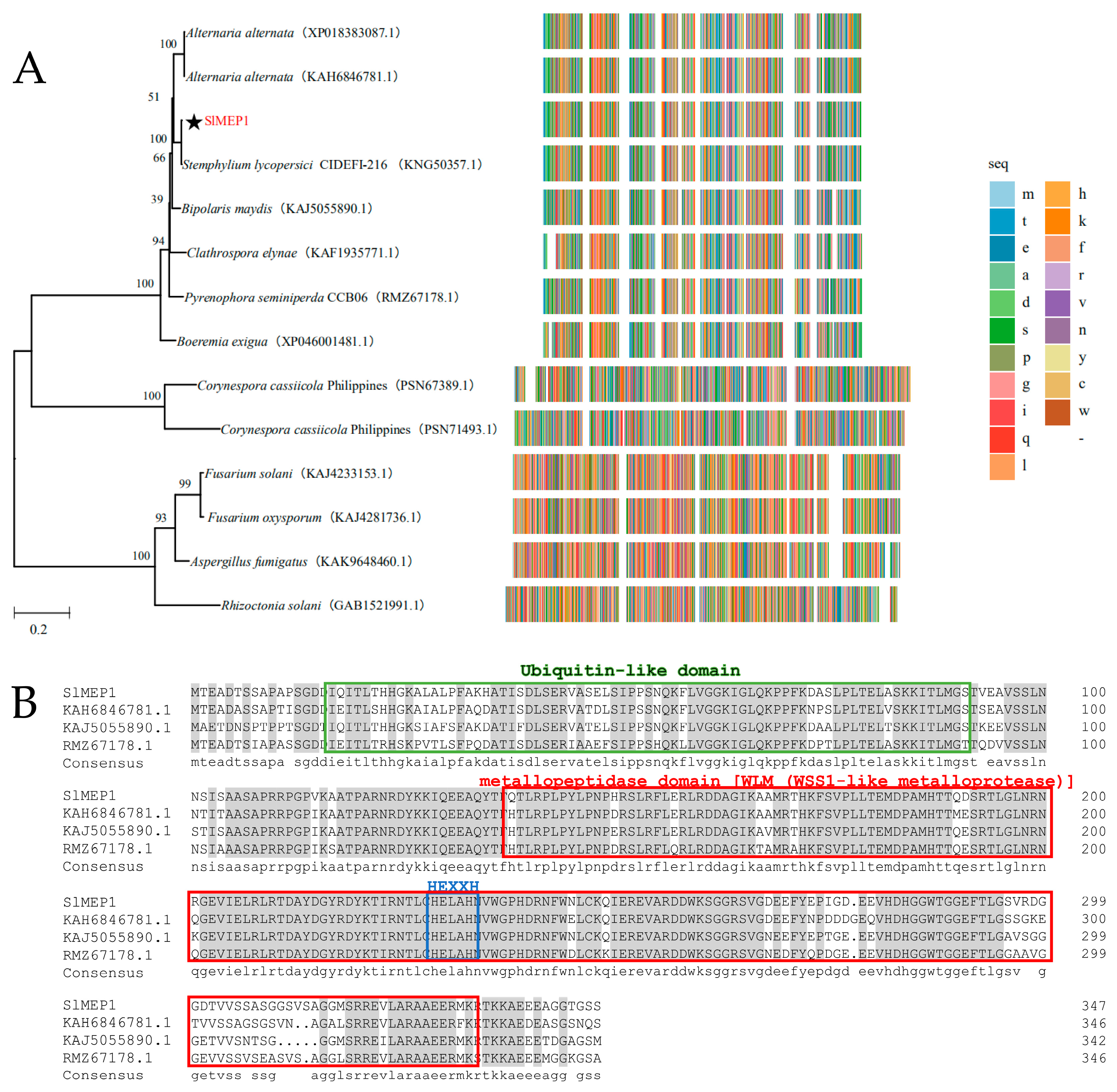
3.1. Phylogenetic Analysis and Sequence Alignment of Different Fungal Zinc Metalloproteases
3.2. Transcriptional Profiling of SlMEP1 During Fungal Pathogenesis of S. lycopersici to Tomato
3.3. Reduced Pathogenicity of S. lycopersici Due to Deletion of SlMEP1
3.4. Impaired Hyphal Penetration of S. lycopersici Due to SlMEP1 Deletion
3.5. SlMEP1 Induced Cell Collapse in Inoculated Tomato Leaves
3.6. Enhanced Pathogenicity of S. lycopersici Due to Added SlMEP1 Protein
3.7. The Chitinase Gene Expression in Tomato Affected by the SlMEP1 Protein
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simmons, E.G. Perfect states of Stemphylium—IV. Harv. Pap. Bot. 2001, 6, 199–208. [Google Scholar]
- Yang, H.H.; Zhao, T.T.; Jiang, J.B.; Wang, S.B.; Wang, A.X.; Li, J.F.; Xu, X.Y. Mapping and screening of the tomato Stemphylium lycopersici resistance gene, Sm, based on bulked segregant analysis in combination with genome resequencing. BMC Plant Biol. 2017, 17, 266. [Google Scholar] [CrossRef]
- Liu, A.M.; Sun, J.D.; Tao, X.Z.; Qu, Q.W.; Xu, D.M.; Wang, T. Occurrence and integrated control of grey leaf spot of tomato in protected areas. China Plant Prot. Guide 2004, 2004, 22–23. (In Chinease) [Google Scholar]
- Xie, X.W.; Wu, J.; Cheng, Y.C.; Shi, J.J.; Zhang, X.J.; Shi, Y.X.; Chai, A.L.; Li, B.J. First report of Stemphylium lycopersici causing leaf spot on hot pepper in China. Can. J. Plant Pathol. 2019, 41, 124–128. [Google Scholar] [CrossRef]
- Jiang, W.; Bond, J.S. Families of metalloendopeptidases and their relationships. FEBS Lett. 1992, 321, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Tallant, C.; García-Castellanos, R.; Seco, J.; Baumann, U.; Gomis-Rüth, F.X. Molecular analysis of ulilysin, the structural prototype of a new family of metzincin metalloproteases. J. Biol. Chem. 2006, 281, 17920–17928. [Google Scholar] [CrossRef]
- Zhou, R.; Zhou, X.Z.; Fan, A.L.; Wang, Z.X.; Huang, B. Differential functions of two metalloproteases, Mrmep1 and Mrmep2, in growth, sporulation, cell wall integrity, and virulence in the filamentous fungus Metarhizium robertsii. Front. Microbiol. 2018, 9, 1528. [Google Scholar] [CrossRef]
- Laursen, L.S.; Overgaard, M.T.; Nielsen, C.G.; Boldt, H.B.; Hopmann, K.H.; Conover, C.A.; Sottrup-Jensen, L.; Giudice, L.C.; Oxvig, C. Substrate specificity of the metalloproteinase pregnancy-associated plasma protein-A (PAPP-A) assessed by mutagenesis and analysis of synthetic peptides: Substrate residues distant from the scissile bond are critical for proteolysis. Biochem. J. 2002, 367, 31–40. [Google Scholar] [CrossRef]
- Glerup, S.; Boldt, H.B.; Overgaard, M.T.; Sottrup-Jensen, L.; Giudice, L.C.; Oxvig, C. Proteinase inhibition by proform of eosinophil major basic protein (pro-MBP) is a multistep process of intra- and intermolecular disulfide rearrangements. J. Biol. Chem. 2005, 280, 9823–9832. [Google Scholar] [CrossRef]
- Wang, X.T.; He, M.M.; Liu, H.H.; Ding, H.Y.; Liu, K.H.; Li, Y.; Cheng, P.; Li, Q.; Wang, B.T. Functional characterization of the M36 cetalloprotease FgFly1 in Fusarium graminearum. J. Fungi 2022, 8, 726. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Bak, D.D.; Heid, H.; Geider, K. Molecular characterization of a protease secreted by Erwinia amylovora. J. Mol. Biol. 1999, 289, 1239–1251. [Google Scholar] [CrossRef]
- Dow, M.; Newman, M.-A.; von Roepenack, E. The induction and modulation of plant defense responses by Bacterial lipopolysaccharides. Annu. Rev. Phytopathol. 2001, 38, 241–261. [Google Scholar] [CrossRef]
- Jia, Y.L.; McAdams, S.A.; Bryan, G.T.; Hershey, H.P.; Valent, B. Direct interaction of resistance gene and avirulence gene products confers rice blast resistance. EMBO J. 2000, 19, 4004–4014. [Google Scholar] [CrossRef]
- Sanz-Martín, J.M.; Pacheco-Arjona, J.R.; Bello-Rico, V.; Vargas, W.A.; Monod, M.; Díaz-Mínguez, J.M.; Thon, M.R.; Sukno, S.A. A highly conserved metalloprotease effector enhances virulence in the maize anthracnose fungus Colletotrichum graminicola. Mol. Plant Pathol. 2016, 19, 2044–2055. [Google Scholar] [CrossRef]
- Vargas, W.A.; Martín, J.M.S.; Rech, G.E.; Rivera, L.P.; Benito, E.P.; Díaz-Mínguez, J.M.; Thon, M.R.; Sukno, S.A. Plant defense mechanisms are activated during biotrophic and necrotrophic development of Colletotricum graminicola in maize. Plant Physiol. 2012, 158, 1342–1358. [Google Scholar] [CrossRef]
- Pan, L.J.; Lu, L.; Liu, Y.P.; Wen, S.X.; Zhang, Z.Y. The M43 domain-containing metalloprotease RcMEP1 in Rhizoctonia cerealis is a pathogenicity factor during the fungus infection to wheat. J. Integr. Agric. 2020, 19, 2044–2055. [Google Scholar] [CrossRef]
- Pan, L.J.; Wen, S.X.; Yu, J.F.; Lu, L.; Zhu, X.L.; Zhang, Z.Y. Genome-wide identification of M35 family metalloproteases in Rhizoctonia cerealis and functional analysis of RcMEP2 as a virulence factor during the fungal infection to wheat. Int. J. Mol. Sci. 2020, 21, 2984. [Google Scholar] [CrossRef]
- Guo, F.L.; Pan, L.J.; Liu, H.W.; Lv, L.J.; Chen, X.Y.; Liu, Y.P.; Li, H.; Ye, W.W.; Zhang, Z.Y. Whole-genome metalloproteases in the wheat sharp eyespot pathogen Rhizoctonia cerealis and a role in fungal virulence. Int. J. Mol. Sci. 2022, 23, 10691. [Google Scholar] [CrossRef]
- Wang, X.T.; Liu, K.H.; Li, Y.; Ren, Y.Y.; Li, Q.; Wang, B.T. Zinc metalloprotease FgM35, which targets the wheat zinc-binding protein TaZnBP, contributes to the virulence of Fusarium graminearum. Stress Biol. 2024, 4, 45. [Google Scholar]
- Zhang, D.Z.; Chi, W.J.; Wang, C.C.; Dai, H.J.; Li, J.T.; Li, C.L.; Li, F.J. Pathogenic process-associated transcriptome analysis of Stemphylium lycopersici from tomato. Int. J. Genom. 2022, 2022, 4522132. [Google Scholar] [CrossRef]
- Ökmen, B.; Kemmerich, B.; Hilbig, D.; Wemhöner, R.; Aschenbroich, J.; Perrar, A.; Huesgen, P.F.; Schipper, K.; Doehlemann, G. Dual function of a secreted fungalysin metalloprotease in Ustilago maydis. New Phytol. 2018, 220, 249–261. [Google Scholar] [CrossRef]
- Naumann, T.A.; Wicklow, D.T.; Price, N.P.J. Identification of a chitinase-modifying protein from Fusarium verticillioides: Truncation of a host resistance protein by a fungalysin metalloprotease. J. Biol. Chem. 2011, 286, 35358–35366. [Google Scholar] [CrossRef]
- Naumann, T.A.; Wicklow, D.T. Chitinase modifying proteins from phylogenetically distinct lineages of Brassica pathogens. Physiol. Mol. Plant Pathol. 2013, 82, 1–9. [Google Scholar] [CrossRef]
- Jashni, M.K.; Dols, I.H.M.; Iida, Y.; Boeren, S.; Beenen, H.G.; Mehrabi, R.; Collemare, J.; de Wit, P.J.G.M. Synergistic action of a metalloprotease and a serine protease from Fusarium oxysporum f. sp. lycopersici cleaves chitin-binding tomato chitinases, reduces their antifungal activity, and enhances fungal virulence. Mol. Plant Microbe Interact. 2015, 28, 996–1008. [Google Scholar]
- Cho, H.J.; Kim, B.S.; Hwang, H.S. Resistance to gray leaf spot in capsicum peppers. HortScience 2019, 36, 752–754. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Lu, L.; Liu, Y.W.; Zhang, Z.Y. Global characterization of GH10 family xylanase genes in Rhizoctonia cerealis and functional analysis of xylanase RcXYN1 during fungus infection in wheat. Int. J. Mol. Sci. 2020, 21, 1812. [Google Scholar] [CrossRef]
- Dong, N.; Liu, X.; Lu, Y.; Du, L.P.; Xu, H.J.; Liu, H.X.; Xin, Z.Y.; Zhang, Z.Y. Overexpression of TaPIEP1, a pathogen-induced ERF gene of wheat, confers host-enhanced resistance to fungal pathogen Bipolaris sorokiniana. Funct. Integr. Genom. 2010, 10, 215–226. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 10, 215–226. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Lo Presti, L.; Lanver, D.; Schweizer, G.; Tanaka, S.; Liang, L.; Tollot, M.; Zuccaro, A.; Reissmann, S.; Kahmann, R. Fungal effectors and plant susceptibility. Annu. Rev. Plant Biol. 2015, 66, 513–545. [Google Scholar] [CrossRef]
- Franco, M.E.E.; Troncozo, M.I.; López, S.M.Y.; Lucentini, G.; Medina, R.; Saparrat, M.C.N.; Ronco, L.B.; Balatti, P.A. A survey on tomato leaf grey spot in the two main production areas of Argentina led to the isolation of Stemphylium lycopersici representatives which were genetically diverse and differed in their virulence. Eur. J. Plant Pathol. 2017, 149, 983–1000. [Google Scholar] [CrossRef]
- Yang, H.H.; Xu, X.Y.; Zhao, T.T.; Jiang, J.B.; Liu, G.; Li, J.F. First report of Stemphylium lycopersici causing gray leaf spot on eggplant in China. Plant Dis. 2017, 101, 834. [Google Scholar] [CrossRef]
- Liu, H.; Wang, H.; Zhong, J.; Lu, X.; Pan, X.T.; Zhu, H.J.; Zhou, Q. First report of Stemphylium lycopersici and Stemphylium vesicarium causing leaf spot on lettuce (Lactuca sativa) in China. Plant Dis. 2019, 103, 2957. [Google Scholar] [CrossRef]
- Yang, H.H.; Li, Y.; He, Y.Q.; Sun, Y.G.; Jiang, J.B.; Zhao, T.T.; Zhang, H.; Xu, X.Y.; Li, J.F. First report of Stemphylium lycopersici causing gray leaf spot on physali (Physalis alkekengi) in China. Plant Dis. 2020, 104, 2725. [Google Scholar] [CrossRef]
- Tomioka, K.; Abe, D.; Kawaguchi, A.; Kawakami, A.; Masunaka, A.; Mori, S.; Nomiyama, K.; Sekiguchi, H. Stemphylium leaf spot of asparagus caused by Stemphylium lycopersici. J. Gen. Plant Pathol. 2020, 87, 51–54. [Google Scholar] [CrossRef]
- Ben, H.Y.; Huo, J.F.; Yao, Y.R.; Gao, W.; Wang, W.L.; Hao, Y.J.; Zhang, X.Y. Stemphylium lycopersici Causing Leaf Spot of Watermelon (Citrullus lanatus) in China. Plant Dis. 2021, 105, 4157. [Google Scholar] [CrossRef]
- Spawton, K.A.; du Toit, L.J. Characterization of Stemphylium species associated with stemphylium leaf spot of spinach (Spinacia oleracea). Plant Dis. 2024, 108, 3578–3594. [Google Scholar] [CrossRef]
- Lei, C.Y.; Gou, Y.N.; Wu, N.; Liu, F.Y.; Deng, J.X. First report of leaf blight caused by Stemphylium lycopersici on Salvia splendens in China. Plant Dis. 2024, 108, 1893. [Google Scholar] [CrossRef]
- Franco, M.E.E.; López, S.; Medina, R.; Saparrat, M.C.N.; Balatti, P. Draft genome sequence and gene annotation of Stemphylium lycopersici strain CIDEFI-216. Genome Announc. 2015, 3, e01069-15. [Google Scholar] [CrossRef]
- Medina, R.; Franco, M.E.E.; Lucentini, G.; Saparrat, M.C.N.; Balatti, P.A. Draft genome sequences of sporulating (CIDEFI-213) and nonsporulating (CIDEFI-212) strains of Stemphylium lycopersici. Microbiol. Resour. Announc. 2018, 7, e00960-18. [Google Scholar] [CrossRef]
- Zeng, R.; Gao, S.G.; Xu, L.H.; Liu, X.; Dai, F.M. Prediction of pathogenesis-related secreted proteins from Stemphylium lycopersici. BMC Microbiol. 2018, 18, 191. [Google Scholar] [CrossRef]
- Medina, R.; Franco, M.E.E.; Bartel, L.C.; Lucentini, C.G.; López Diaz, S.M.Y.; Saparrat, M.C.N.; Balatti, P.A. Genome analysis of three isolates of Stemphylium lycopersici differ in their virulence and sporulation ability: Identification of effectors, pathogenesis and virulence factors. Plant Pathol. 2024, 73, 1156–1168. [Google Scholar] [CrossRef]
- Lian, J.J.; Han, H.Y.; Chen, X.Z.; Chen, Q.; Zhao, J.H.; Li, C.Y. Stemphylium lycopersici Nep1-like protein (NLP) Is a key virulence factor in tomato gray leaf spot disease. J. Fungi 2022, 8, 518. [Google Scholar] [CrossRef]
- Lv, J.Y.; Zhou, J.L.; Chang, B.Y.; Zhang, Y.H.; Feng, Z.L.; Wei, F.; Zhao, L.H.; Zhang, Y.L.; Feng, H.J. Two metalloproteases VdM35-1 and VdASPF2 from Verticillium dahliae are required for fungal pathogenicity, stress adaptation, and activating immune response of host. Microbiol. Spectr. 2022, 10, e0247722. [Google Scholar] [CrossRef]
- El Gueddari, N.E.; Rauchhaus, U.; Moerschbacher, B.M.; Deising, H.B. Developmentally regulated conversion of surface-exposed chitin to chitosan in cell walls of plant pathogenic fungi. New Phytol. 2002, 156, 103–112. [Google Scholar] [CrossRef]
- Han, L.B.; Li, Y.B.; Wang, F.X.; Wang, W.Y.; Liu, J.; Wu, J.H.; Zhong, N.Q.; Wu, S.J.; Jiao, G.L.; Wang, H.Y.; et al. The cotton apoplastic protein CRR1 stabilizes chitinase 28 to facilitate defense against the fungal pathogen Verticillium dahliae. Plant Cell. 2019, 31, 520–536. [Google Scholar] [CrossRef]
- Zhu, X.H.; Sayari, M.; Islam, M.R.; Daayf, F. NOXA is important for Verticillium dahliae’s penetration ability and virulence. J. Fungi 2021, 7, 814. [Google Scholar] [CrossRef]
- Chen, L.H.; Ma, X.H.; Sun, T.G.; Zhu, Q.H.; Feng, H.J.; Li, Y.T.; Liu, F.; Zhang, X.Y.; Sun, J.; Li, Y.J. VdPT1 encoding a neutral trehalase of Verticillium dahliae is required for growth and virulence of the pathogen. Int. J. Mol. Sci. 2023, 25, 294. [Google Scholar] [CrossRef]
- Wang, D.h.; Wen, S.l.; Zhao, Z.b.; Long, Y.h.; Fan, R. Hypothetical protein VDAG_07742 is required for Verticillium dahliae pathogenicity in potato. Int. J. Mol. Sci. 2023, 24, 3630. [Google Scholar] [CrossRef]
- Li, Y.T.; Song, S.L.; Chen, B.; Zhang, Y.; Sun, T.G.; Ma, X.H.; Li, Y.J.; Sun, J.; Zhang, X.Y. Deleting an xylosidase-encoding gene VdxyL3 increases growth and pathogenicity of Verticillium dahlia. Front. Microbiol. 2024, 15, 1428780. [Google Scholar] [CrossRef] [PubMed]
- Azizullah; Noman, M.; Gao, Y.Z.; Wang, H.; Xiong, X.H.; Wang, J.J.; Li, D.Y.; Song, F.M. The SUMOylation pathway regulates the pathogenicity of Fusarium oxysporum f. sp. niveum in watermelon through stabilizing the pH regulator FonPalC via SUMOylation. Microbiol. Res. 2024, 281, 127632. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, D.; Chi, W.; Li, T.; Wang, C.; Zhang, J.; Pan, H.; Qiao, N.; Li, J.; Sun, X. Zinc Metalloprotease SlMEP1: An Essential Factor Required for Fungal Virulence in Stemphylium lycopersici. J. Fungi 2025, 11, 330. https://doi.org/10.3390/jof11050330
Zhang D, Chi W, Li T, Wang C, Zhang J, Pan H, Qiao N, Li J, Sun X. Zinc Metalloprotease SlMEP1: An Essential Factor Required for Fungal Virulence in Stemphylium lycopersici. Journal of Fungi. 2025; 11(5):330. https://doi.org/10.3390/jof11050330
Chicago/Turabian StyleZhang, Dezhen, Wenjuan Chi, Tingting Li, Cuicui Wang, Jing Zhang, Haoqin Pan, Ning Qiao, Jintang Li, and Xiaoan Sun. 2025. "Zinc Metalloprotease SlMEP1: An Essential Factor Required for Fungal Virulence in Stemphylium lycopersici" Journal of Fungi 11, no. 5: 330. https://doi.org/10.3390/jof11050330
APA StyleZhang, D., Chi, W., Li, T., Wang, C., Zhang, J., Pan, H., Qiao, N., Li, J., & Sun, X. (2025). Zinc Metalloprotease SlMEP1: An Essential Factor Required for Fungal Virulence in Stemphylium lycopersici. Journal of Fungi, 11(5), 330. https://doi.org/10.3390/jof11050330