
The Characterization of the Purine Nucleoside Phosphorylase from Agaricus bisporus and Its Potential Application in Reducing Purine Content in Beer


Abstract
1. Introduction
2. Materials and Methods
2.1. Plasmids and Strains
2.2. Structural Bioinformatics Analysis
2.3. Expression and Purification of PNPs in P. pastoris
2.4. Enzyme Activity Assay
2.5. Biochemical Characterization of AbPNP
2.6. Wort Preparation
2.7. Wort Fermentation
2.8. Determination of Total Purine Content
2.9. Analytical Methods
2.10. Statistical Analysis
3. Results and Discussion
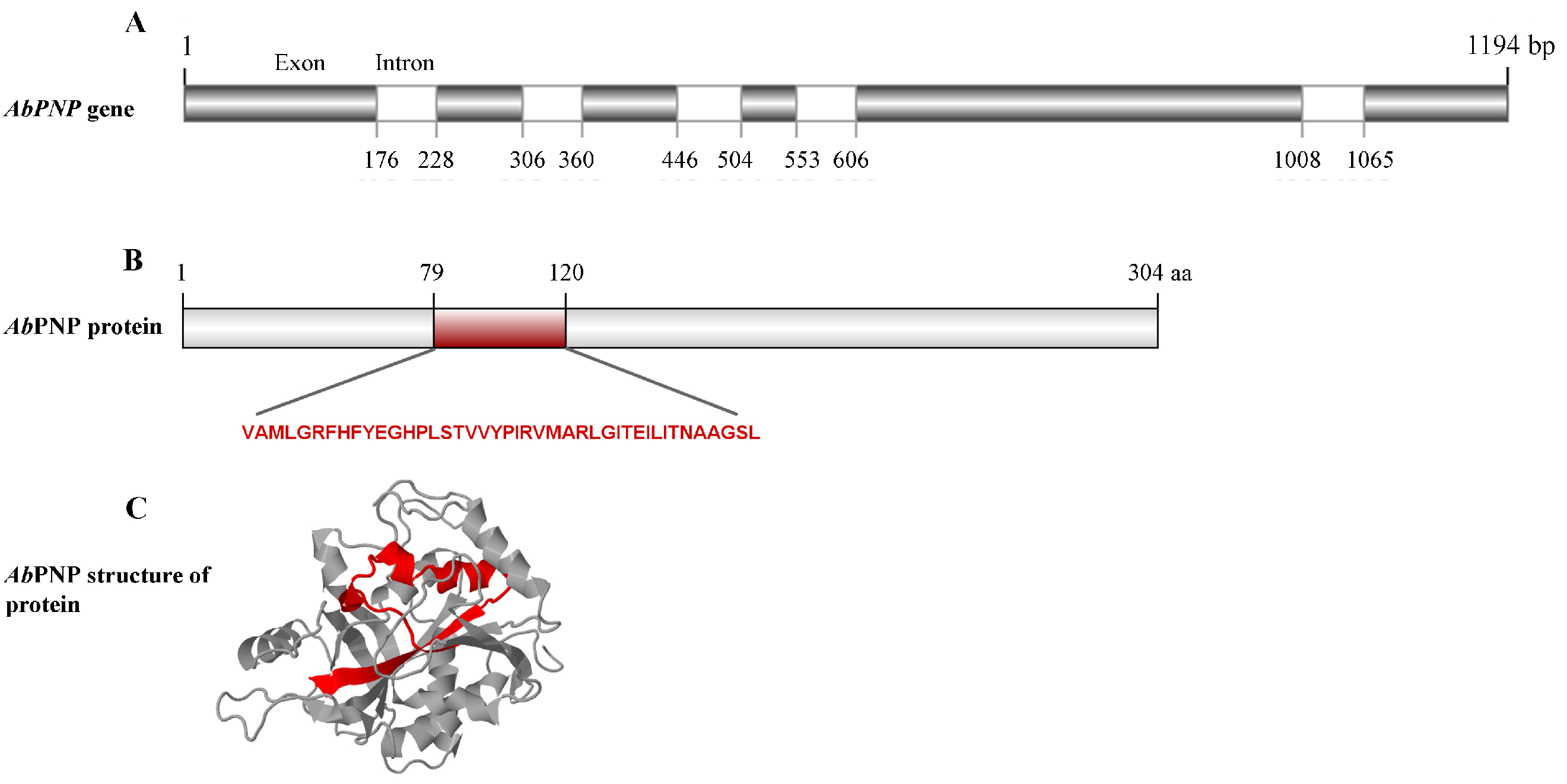
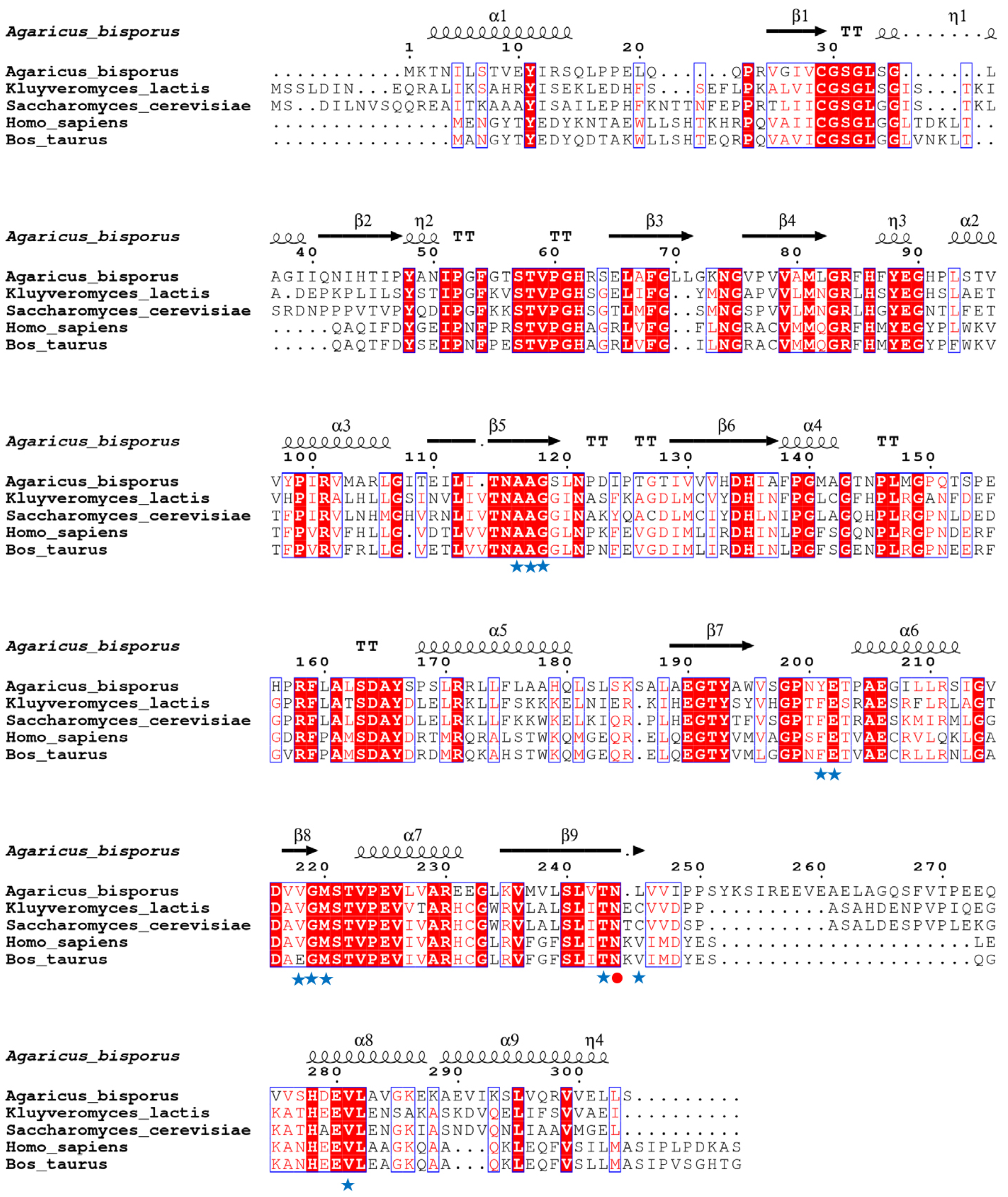
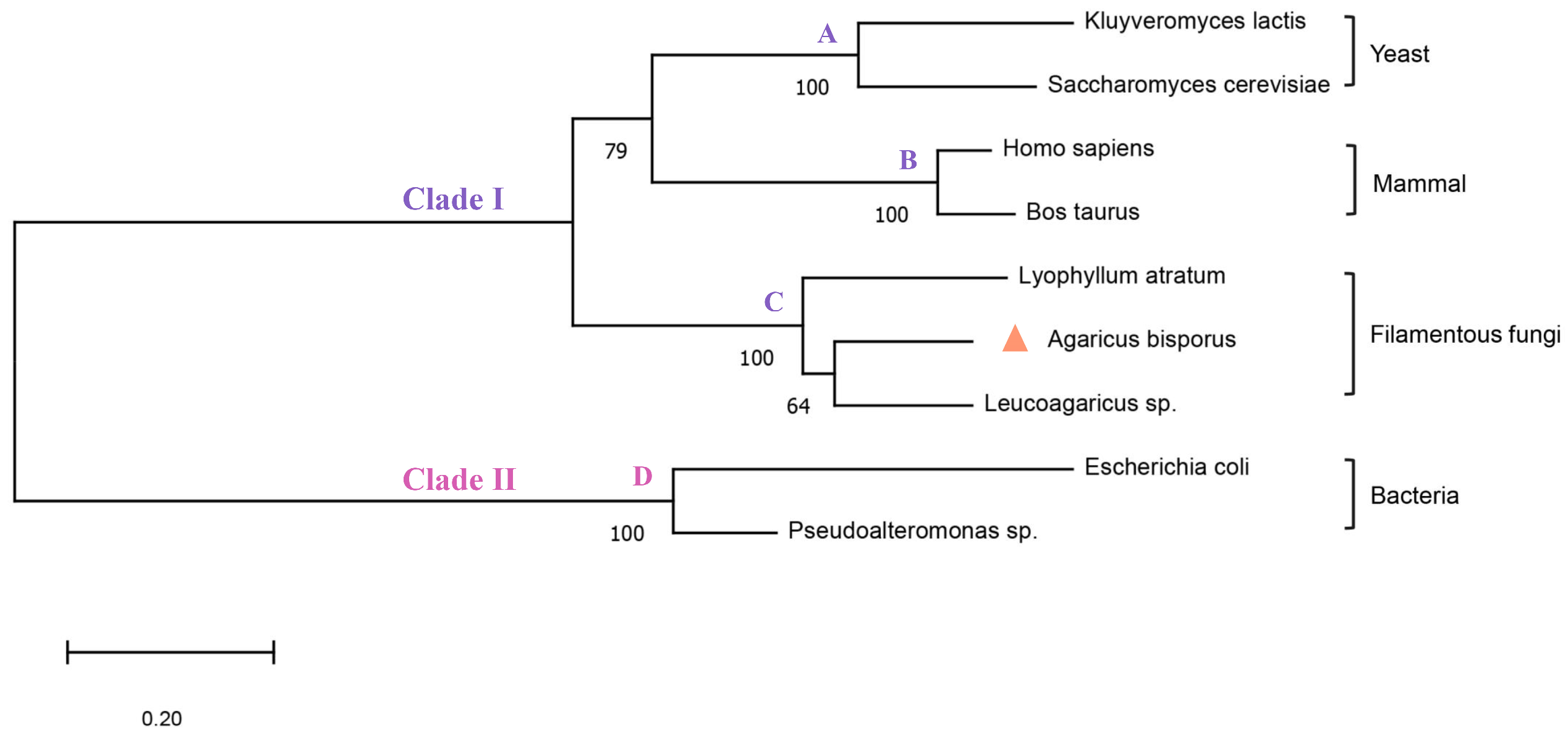
3.1. The Analysis of Amino Acid Structure and Sequence
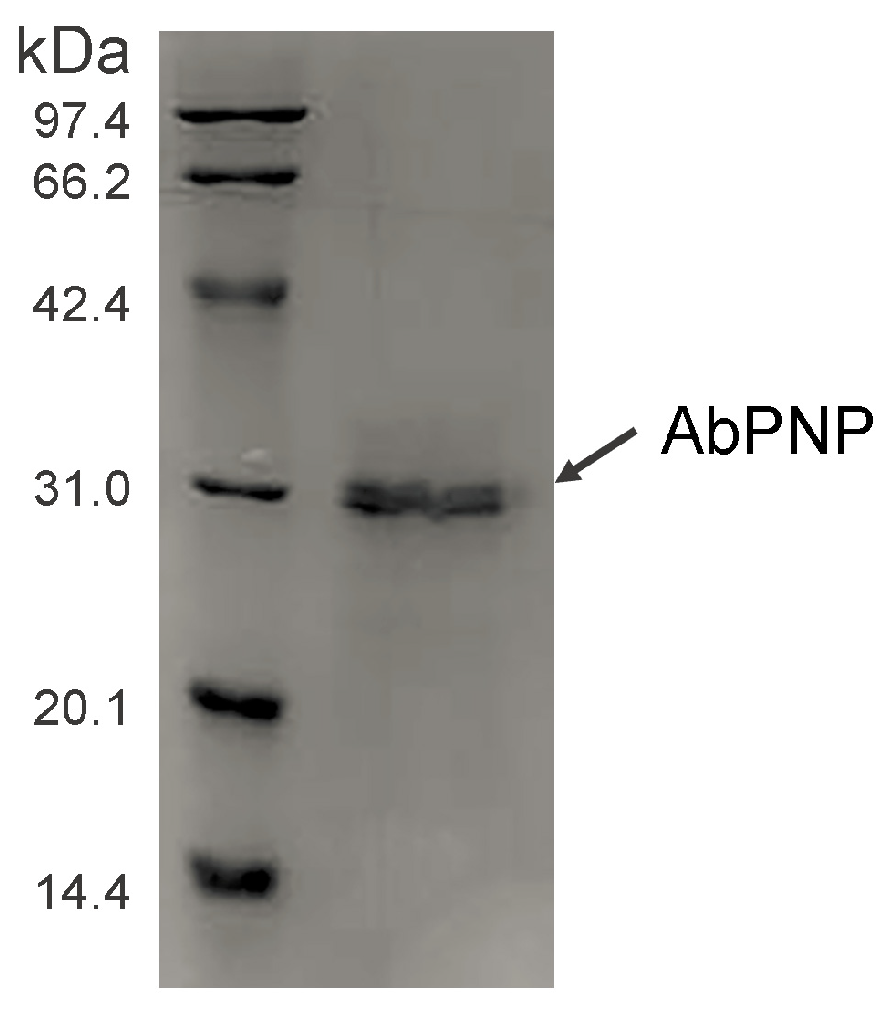
3.2. Expression of Recombinant AbPNP in P. pastoris
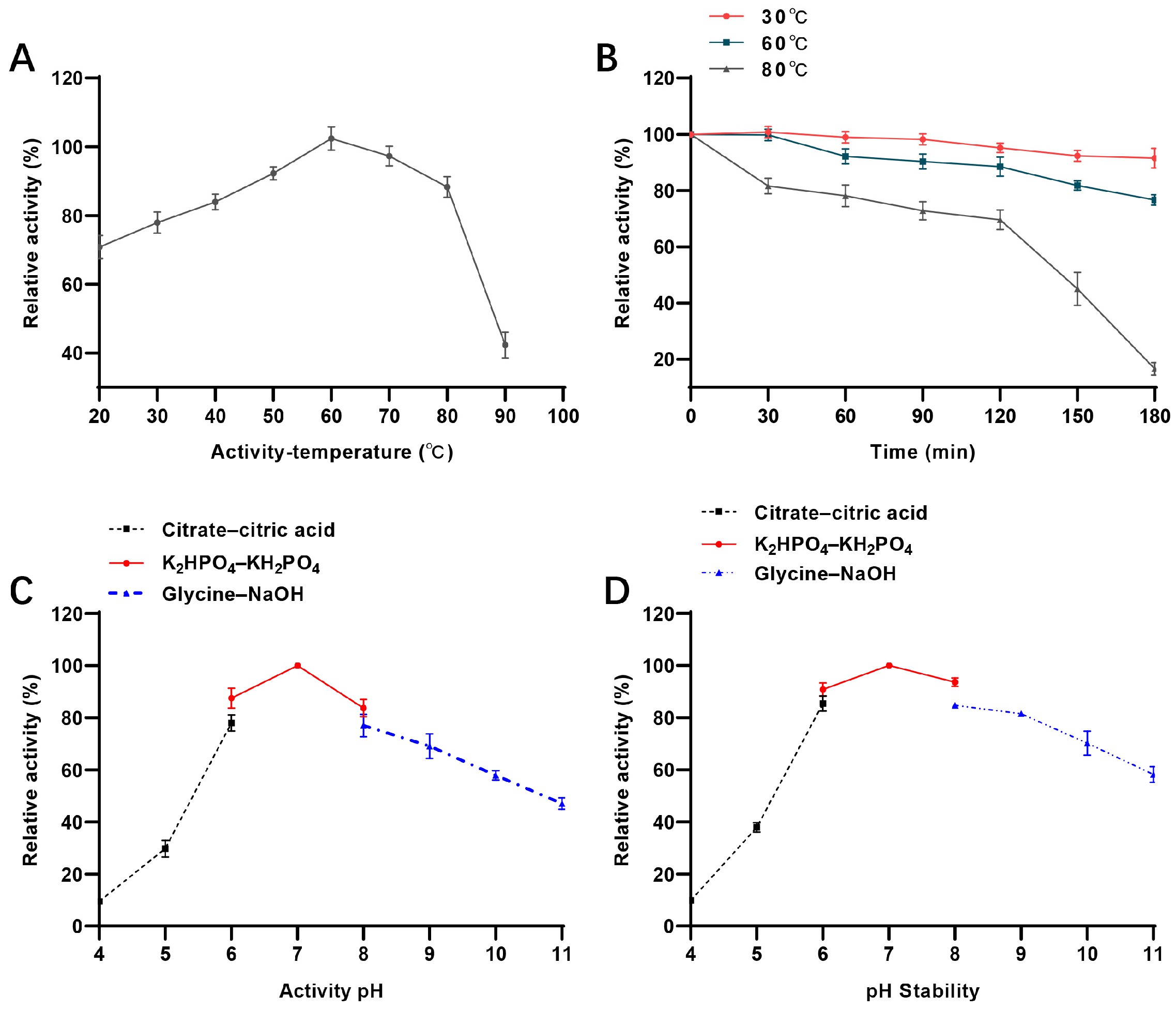
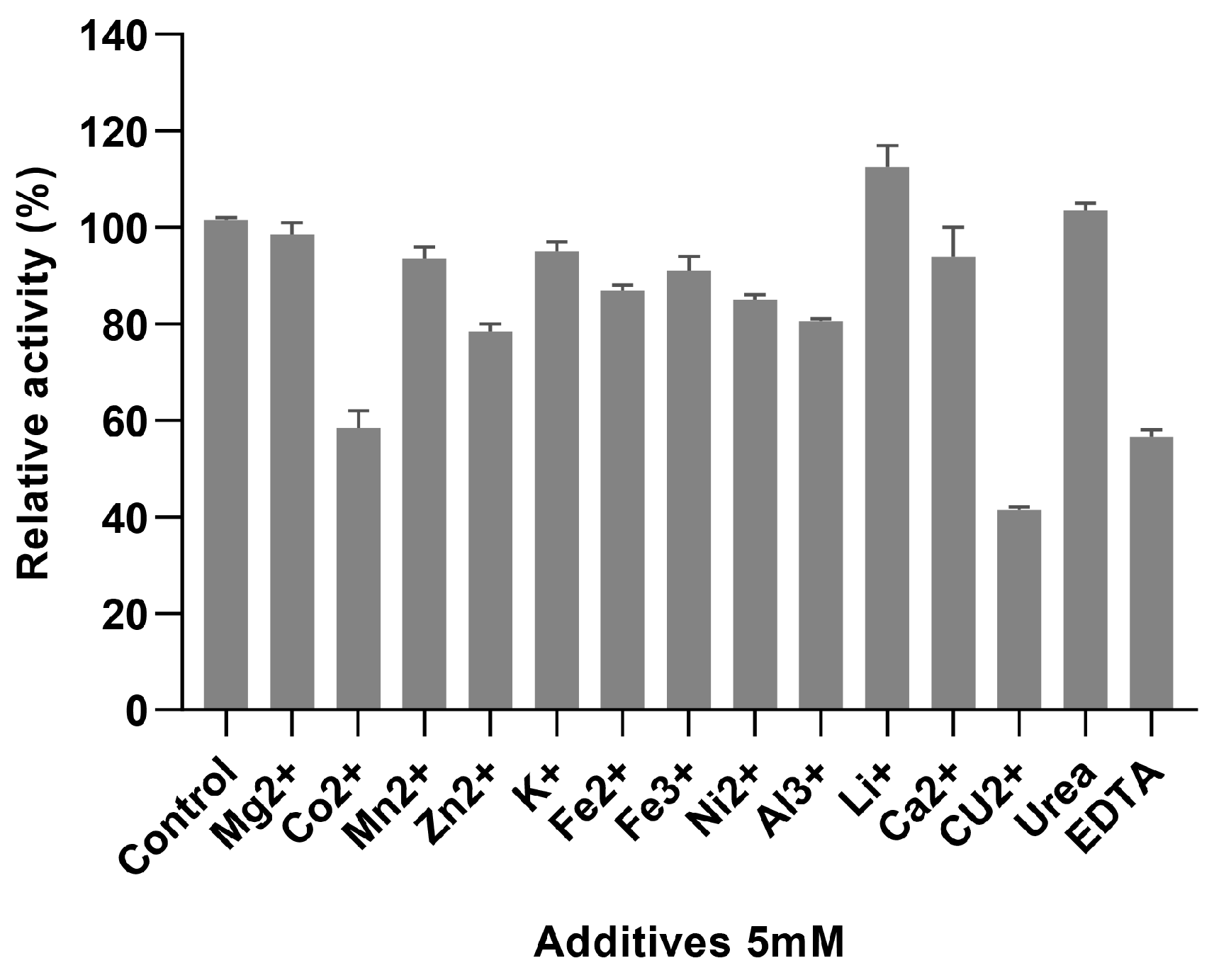
3.3. Enzymatic Properties of AbPNP
3.4. Substrate Specificity and Enzyme Kinetics
3.5. Nucleoside Degradation Analysis in the Mashing Process
3.6. The Effect of AbPNP on Physicochemical Parameters and Purines of Beer
3.7. The Effect of AbPNP on Beer Aroma Compounds
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aihemaitijiang, S.; Zhang, Y.; Zhang, L.; Yang, J.; Ye, C.; Halimulati, M.; Zhang, W.; Zhang, Z. The association between purine-rich food intake and hyperuricemia: A cross-sectional study in Chinese adult residents. Nutrients 2020, 12, 3835. [Google Scholar] [CrossRef]
- Feng, S.M.; Wu, S.J.; Xie, F.; Yang, C.S.; Shao, P. Natural compounds lower uric acid levels and hyperuricemia: Molecular mechanisms and prospective. Trends Food Sci. Technol. 2022, 123, 87–102. [Google Scholar]
- Major, T.J.; Topless, R.K.; Dalbeth, N.; Merriman, T.R. Evaluation of the diet wide contribution to serum urate levels: Meta-analysis of population based cohorts. BMJ 2018, 363, k3951. [Google Scholar]
- Kaneko, K.; Yamanobe, T.; Fujimori, S. Determination of purine contents of alcoholic beverages using high performance liquid chromatography. Biomed. Chromatogr. 2009, 23, 858–864. [Google Scholar] [CrossRef]
- Li, H.; Liu, F.; Hao, J.; Liu, C. Determination of purines in beer by HPLC using a simple and rapid sample pretreatment. J. Am. Soc. Brew. Chem. 2015, 73, 137–142. [Google Scholar]
- Nicholls, A.; Scott, J.T. Effect of weight-loss on plasma and urinary levels of uric acid. Lancet 1972, 300, 1223–1224. [Google Scholar]
- Levinson, W.; Cohen, M.S.; Brady, D.; Duffy, F.D. To change or not to change: “Sounds like you have a dilemma”. Ann. Intern. Med. 2001, 135, 386–391. [Google Scholar] [CrossRef]
- Shibano, Y.; Yomo, H.; Matsumoto, T.; Koda, H.; Suwa, Y.; Amachi, T.; Hatanaka, H.; Shimizu, S. Process for Manufacturing Beer. Patent EP0753572A1, 16 February 1996. [Google Scholar]
- Pugmire, M.J.; Ealick, S.E. Structural analyses reveal two distinct families of nucleoside phosphorylases. Biochem. J. 2002, 361, 1–25. [Google Scholar]
- Breer, K.; Girstun, A.; Wielgus-Kutrowska, B.; Staroń, K.; Bzowska, A. Overexpression, purification and characterization of functional calf purine nucleoside phosphorylase (PNP). Protein Expr. Purif. 2008, 61, 122–130. [Google Scholar]
- Silva, R.G.; Carvalho, L.P.S.; Oliveira, J.S.; Pinto, C.A.; Mendes, M.A.; Palma, M.S.; Basso, L.A.; Santos, D.S. Cloning, overexpression, and purification of functional human purine nucleoside phosphorylase. Protein Expr. Purif. 2003, 27, 158–164. [Google Scholar] [CrossRef]
- Spohner, S.C.; Schaum, V.; Quitmann, H.; Czermak, P. Kluyveromyces lactis: An emerging tool in biotechnology. J. Biotechnol. 2016, 222, 104–116. [Google Scholar] [PubMed]
- Mahor, D.; Priyanka, A.; Prasad, G.S.; Thakur, K.G. Functional and structural characterization of purine nucleoside phosphorylase from Kluyveromyces lactis and its potential applications in reducing purine content in food. PLoS ONE 2016, 11, e0164279. [Google Scholar]
- De Windt, F.E.; Wagemaker, M.J.M.; Op den Camp, H.J.M.; van der Drift, C. Purine degradation in the edible mushroom Agaricus bisporus. Folia Microbiol. 2002, 47, 672–676. [Google Scholar]
- Duan, H.; Wang, H.; Ma, B.; Jiang, P.; Tu, P.; Ni, Z.; Li, X.; Li, M.; Ma, X.; Wang, B.; et al. Codon optimization and expression of irisin in Pichia pastoris GS115. Int. J. Biol. Macromol. 2015, 79, 21–26. [Google Scholar]
- Bao, M.; Niu, C.; Xu, X.; Zheng, F.; Liu, C.; Wang, J.; Li, Q. Identification, soluble expression, and characterization of a novel endo-inulinase from Lipomyces starkeyi NRRL Y-11557. Int. J. Biol. Macromol. 2019, 137, 537–544. [Google Scholar] [PubMed]
- Luo, W.; Liu, Y.P.; Zhu, X.C.; Zhao, W.J.; Huang, L.; Cai, J.; Xu, Z.N.; Cen, P.L. Cloning and characterization of purine nucleoside phosphorylase in Escherichia coli and subsequent ribavirin biosynthesis using immobilized recombinant cells. Enzyme Microb. Technol. 2011, 48, 438–444. [Google Scholar] [PubMed]
- Yin, H.; He, Y.; Deng, Y.; Dong, J.; Lu, J.; Chen, L. Application of Plackett–Burman experimental design for investigating the effect of wort amino acids on flavour-active compounds production during lager yeast fermentation. J. Inst. Brew. 2017, 123, 300–311. [Google Scholar]
- Wielgus, K.B.; Modrak, W.A.; Dyzma, A.; Breer, K.; Zolkiewski, M.; Bzowska, A. Purine nucleoside phosphorylase activity decline is linked to the decay of the trimeric form of the enzyme. Arch. Biochem. Biophys. 2014, 549, 40–48. [Google Scholar]
- Li, X.; Jiang, X.; Li, H.; Ren, D. Purine nucleoside phosphorylase from Pseudoalteromonas sp. Bsi590: Molecular cloning, gene expression and characterization of the recombinant protein. Extremophiles 2008, 12, 325–333. [Google Scholar]
- Bzowska, A.; Kulikowska, E.; Shugar, D. Properties of purine nucleoside phosphorylase (PNP) of mammalian and bacterial origin. Z. Naturforschung C 1990, 45, 59–70. [Google Scholar]
- Koellner, G.; Luić, M.; Shugar, D.; Saenger, W.; Bzowska, A. Crystal structure of calf spleen purine nucleoside phosphorylase in a complex with hypoxanthine at 2.15 A resolution. J. Mol. Biol. 1997, 265, 202–216. [Google Scholar] [CrossRef] [PubMed]
- Lecoq, K.; Belloc, I.; Desgranges, C.; Konrad, M.; Daignan-Fornier, B. YLR209c encodes Saccharomyces cerevisiae purine nucleoside phosphorylase. J. Bacteriol. 2001, 183, 4910–4913. [Google Scholar] [PubMed]
- Li, Y.; Li, N. The main inorganic ions in the wort and their originates. Beer Technol. 2013, 18, 44–46. [Google Scholar]
- Ghanem, M.; Li, L.; Wing, C.; Schramm, V.L. Altered thermodynamics from remote mutations altering human toward bovine purine nucleoside phosphorylase. Biochemistry 2008, 47, 2559–2564. [Google Scholar]
- Xie, X.X.; Huo, W.T.; Xia, J.G.; Xu, Q.Y.; Chen, N. Structure-activity relationship of a cold-adapted purine nucleoside phosphorylase by site-directed mutagenesis. Enzyme Microb. Technol. 2012, 51, 59–65. [Google Scholar]
- Li, Y.M. Expression of Purine Nucleoside Phosphorylase in Escherichia coli and Its Application. Master’s Thesis, Jiangnan University, Wuxi, China, 2016. [Google Scholar]
- Wang, M.; Sun, Z.; Wang, Y.; Wei, Z.; Chen, B.; Zhang, H.; Guo, X.; Xiao, D. The effect of pitching rate on the production of higher alcohols by top-fermenting yeast in wheat beer fermentation. Ann. Microbiol. 2019, 69, 713–726. [Google Scholar]
- Hong, K.-Q.; Fu, X.-M.; Dong, S.-S.; Xiao, D.-g.; Dong, J. Modulating acetate ester and higher alcohol production in Saccharomyces cerevisiae through the cofactor engineering. J. Ind. Microbiol. Biotechnol. 2019, 46, 1003–1011. [Google Scholar] [CrossRef]
- Yang, M.; Hao, J.; Zhang, R.; He, R.; Ma, H. Breeding High-Yield Ethyl Caproate-Producing Saccharomyces cerevisiae in Sake: Flux Regulation from Glycolytic Fermentation to the FAS Pathway and Alcohol Acyltransferase Overexpression. J. Agric. Food Chem. 2025. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | CK | AbPNP Addition |
|---|---|---|
| Alcohol %/vol | 4.48 ± 0.05 | 4.64 ± 0.03 * |
| RDF/% | 71.79 ± 0.42 | 73.68 ± 0.51 |
| Original extract/% w/w | 12.20 ± 0.15 | 12.21 ± 0.09 |
| Adenine/mg/L | 7.23 ± 0.13 | 6.59 ± 0.24 |
| Guanine/mg/L | 5.04 ± 0.06 | 10.34 ± 0.19 * |
| Hypoxanthine/mg/L | 5.81 ± 0.12 | 6.71 ± 0.08 * |
| Xanthine/mg/L | 8.79 ± 0.57 | 9.90 ± 0.14 * |
| Total purine bases/mg/L | 26.87 | 33.54 |
| Inosine/mg/L | 13.35 ± 0.11 | 4.12 ± 0.04 * |
| Guanosine/mg/L | 87.88 ± 3.76 | 46.38 ± 2.14 * |
| Adenosine/mg/L | 50.30 ± 2.16 | 50.15 ± 1.47 |
| Total purine nucleoside/mg/L | 151.53 | 100.65 |
| Aroma Compounds | CK | AbPNP Addition |
|---|---|---|
| Propanol | 13.08 ± 0.49 | 13.20 ± 0.64 |
| Isobutanol | 26.38 ± 0.77 | 26.51 ± 2.11 |
| Isoamyl alcohol | 51.04 ± 3.41 | 60.73 ± 3.32 * |
| Phenethyl alcohol | 29.17 ± 2.86 | 35.68 ± 1.78 * |
| Total higher alcohols | 119.67 | 136.12 |
| Ethyl acetate | 25.86 ± 2.39 | 27.01 ± 1.54 |
| Isoamyl acetate | 2.97 ± 0.11 | 3.47 ± 0.09 * |
| Ethyl hexanoate | 0.22 ± 0.01 | 0.23 ± 0.01 |
| Ethyl caprylate | 0.19 ± 0.01 | 0.21 ± 0.01 |
| Total esters | 29.24 | 30.92 |
| Ratio of total higher alcohols to esters | 4.09 | 4.40 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Lu, J. The Characterization of the Purine Nucleoside Phosphorylase from Agaricus bisporus and Its Potential Application in Reducing Purine Content in Beer. J. Fungi 2025, 11, 268. https://doi.org/10.3390/jof11040268
Liu J, Lu J. The Characterization of the Purine Nucleoside Phosphorylase from Agaricus bisporus and Its Potential Application in Reducing Purine Content in Beer. Journal of Fungi. 2025; 11(4):268. https://doi.org/10.3390/jof11040268
Chicago/Turabian StyleLiu, Jun, and Jian Lu. 2025. "The Characterization of the Purine Nucleoside Phosphorylase from Agaricus bisporus and Its Potential Application in Reducing Purine Content in Beer" Journal of Fungi 11, no. 4: 268. https://doi.org/10.3390/jof11040268
APA StyleLiu, J., & Lu, J. (2025). The Characterization of the Purine Nucleoside Phosphorylase from Agaricus bisporus and Its Potential Application in Reducing Purine Content in Beer. Journal of Fungi, 11(4), 268. https://doi.org/10.3390/jof11040268