Establishing the Embryonic Axes: Prime Time for Teratogenic Insults

Abstract
:Conflicts of Interest
References
- Wilson, J.G. Current status of teratology: General principles and mechanisms derived from animal studies. In Handbook of Teratology; Wilson, J.G., Fraser, F.C., Eds.; Plenum Press: New York, NY, USA, 1977; Volume 1, pp. 47–74. ISBN 0-306-36241-4. [Google Scholar]
- Takaoka, K.; Yamamoto, M.; Shiratori, H.; Meno, C.; Rossant, J.; Saijoh, Y.; Hamada, H. The mouse embryo autonomously acquires anterior-posterior polarity at implantation. Dev. Cell 2006, 10, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Rossant, J.; Tam, P.P. Blastocyst lineage formation, early embryonic asymmetries and axis patterning in the mouse. Development 2009, 136, 701–713. [Google Scholar] [CrossRef] [PubMed]
- Takaoka, K.; Hamada, H. Cell fate decisions and axis determination in the early mouse embryo. Development 2012, 139, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Takaoka, K.; Yamamoto, M.; Hamada, H. Origin of body axes in the mouse embryo. Curr. Opin. Genet. Dev. 2007, 17, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Vandeberg, L.N.; Levin, M. A unified model for left-right asymmetry? Comparison and synthesis of molecular models of embryonic laterality. Dev. Biol. 2013, 379, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Lenhart, K.F.; Holtzman, N.G.; Williams, J.R.; Burdine, R.D. Integration of nodal and BMP signals in the heart requires FoxH1 to create left-right differences in cell migration rates that direct cardiac asymmetry. PLoS Genet. 2013, 9, e1003109. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Frias, M.L. Primary midline developmental field I. Clinical and epidemiological characteristics. Am. J. Med. Genet. 1995, 56, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Frías, M.L.; Urioste, M.; Bermejo, E.; Rodríguez-Pinilla, E.; Félix, V.; Paisán, L.; Martínez, S.; Egüés, J.; Gómez, F.; Aparicio, P.; et al. Primary midline developmental field. II. Clinical/epidemiological analysis of alteration of laterality (Normal body symmetry and asymmetry. Am. J. Med. Genet. 1995, 56, 382–388. [Google Scholar]
- Kosaki, K.; Casey, B. Genetics of human left-right axis malformations. Semin. Cell Dev. Biol. 1998, 9, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Ticho, B.S.; Goldstein, A.M.; Van Praagh, R. Extracardiac anomalies in the heterotaxy syndromes with focus on anomalies of midline-associated structures. Am. J. Cardiol. 2000, 85, 729–734. [Google Scholar] [CrossRef]
- Morelli, S.H.; Young, L.; Reid, B.; Ruttenberg, H.; Bamshad, M.J. Clinical analysis of families with heart, midline, and laterality defects. Am. J. Med. Genet. 2001, 101, 388–392. [Google Scholar] [CrossRef] [PubMed]
- Ware, S.M.; Peng, J.; Zhu, L.; Fernbach, S.; Colicos, S.; Casey, B.; Towbin, J.; Belmont, J.W. Identification and functional analysis of ZIC3 mutations in heterotaxy and related congenital heart defects. Am. J. Hum. Genet. 2004, 74, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Levin, M. Left-right asymmetry in embryonic development: A comprehensive review. Mech. Dev. 2005, 122, 3–25. [Google Scholar] [CrossRef] [PubMed]
- Bisgrove, B.W.; Morelli, S.H.; Yost, H.J. Genetics of human laterality disorders: Insights from vertebrate model systems. Annu. Rev. Genom. Hum. Genet. 2003, 4, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Ramsdell, A.F. Left-right asymmetry and congenital cardiac defects: Getting to the heart of the matter in vertebrate left-right axis determination. Dev. Biol. 2005, 288, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Logan, M.; Pagan-Westphal, S.M.; Smith, D.M.; Paganessi, L.; Tabin, C.J. The transcription factor Pitx2 mediates situs specific morphogenesis in response to left-right asymmetric signals. Cell 1998, 94, 307–317. [Google Scholar] [CrossRef]
- Piedra, M.E.; Icardo, J.M.; Albajar, M.; Rodriguez-Ray, J.C.; Ros, M.A. Pitx2 participates in the late phase of the pathway controlling left-right asymmetry. Cell 1998, 94, 319–324. [Google Scholar] [CrossRef]
- Ramsdell, A.F.; Bernanke, J.M.; Trusk, T.C. Left-right lineage analysis of the embryonic Xenopus heart reveals a novel framework linking congenital cardiac defects and laterality disease. Development 2006, 133, 1399–1410. [Google Scholar] [CrossRef] [PubMed]
- Bamforth, S.D.; Bragança, J.; Farthing, C.R.; Schneider, J.E.; Broadbent, C.; Michell, A.C.; Clarke, K.; Neubauer, S.; Norris, D.; Brown, N.A.; et al. Cited2 controls left-right patterning and heart development through a Nodal-Pitx2c pathway. Nat. Gen. 2004, 36, 1189–1196. [Google Scholar] [CrossRef] [PubMed]
- Fukumoto, T.; Blakely, R.; Levin, M. Serotonin transporter function is an early step in left-right patterning in chick and frog embryos. Dev. Neurosci. 2005, 27, 349–363. [Google Scholar] [CrossRef] [PubMed]
- Fukumoto, T.; Kema, I.P.; Levin, M. Serotonin signaling is a very early step in patterning of the left-right axis in chick and frog embryos. Curr. Biol. 2005, 15, 794–803. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, K.; Donnet, C.; Rejtar, T.; Karger, B.L.; Barisone, G.A.; Diaz, E.; Kortagere, S.; Lemire, J.M.; Levin, M. Histone deacetylase activity is necessary for left-right patterning during vertebrate development. BMC Dev. Biol. 2011, 11, 29. [Google Scholar] [CrossRef] [PubMed]
- Sadler, T.W. Selective serotonin reuptake inhibitors (SSRIs) and heart defects: Potential mechanisms for the observed associations. Reprod. Toxicol. 2011, 32, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.G.; Jordan, H.C.; Brent, R.L. Effects of radiation on embryonic development. Am. J. Anat. 1953, 92, 153–177. [Google Scholar] [CrossRef] [PubMed]
- Monie, I.W.; Takacs, E.; Warkany, J. Transposition of the great vessels and other cardiovascular abnormalities in rat fetuses induced by trypan blue. Anat. Rec. 1966, 156, 175–190. [Google Scholar] [CrossRef] [PubMed]
- Abu-Issa, R.; Kirby, M.L. Patterning of the heart field in the chick. Dev. Biol. 2008, 319, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Dyer, L.A.; Kirby, M.L. The role of the secondary heart field in cardiac development. Dev. Biol. 2009, 336, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Ferencz, C.; Rubin, J.D.; McCarter, R.J.; Brenner, J.I.; Neill, C.A.; Perry, L.W.; Hepner, S.I.; Downing, J.W. Congenital heart disease: Prevalence at livebirth. The Baltimore-Washington Infant Study. Am. J. Epidemiol. 1985, 121, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Burn, J. Disturbance of morphological laterality in humans. In Ciba Found Symp; Brock, G.R., Marsh, J., Eds.; Wiley: Chinchester, UK, 1991; Volume 162, pp. 47–74. [Google Scholar]
- Gebbia, M.; Ferrero, G.B.; Pilia, G.; Bassi, M.T.; Aylsworth, A.S.; Penman-Splitt, M.; Bird, L.M.; Bamforth, J.S.; Burn, J.; Schlessinger, D.; et al. X-linked situs abnormalities result from mutations in ZIC3. Nat. Genet. 1997, 17, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Casey, B. Two rights make a wrong: Human left-right malformations. Hum. Mol. Genet. 1998, 7, 1565–1571. [Google Scholar] [CrossRef] [PubMed]
- Van Mierop, L.H.S.; Gessner, I.H.; Schiebler, G.L. Asplenia and Polysplenia syndromes. Birth Defects: Orig. Artic. Ser. VIII 1972, 5, 36–44. [Google Scholar]
- Colvin, L.; Slack-Smith, L.; Stanley, F.J.; Bower, C. Dispensing patterns and pregnancy outcomes for women dispensed selective serotonin reuptake inhibitors in pregnancy. Birth Defects Res. (Part A) 2011, 91, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Dagle, J.M.; Sabel, J.L.; Littig, J.L. Pitx2c attenuation results in cardiac defects and abnormalities of intestinal orientation in developing Xenopus laevis. Dev. Biol. 2003, 262, 268–281. [Google Scholar] [CrossRef]
- Gormley, J.P.; Nascone-Yoder, N.M. Left and right contributions to the Xenopus heart: Implications for asymmetric morphogenesis. Dev. Genes Evol. 2003, 213, 390–398. [Google Scholar] [CrossRef] [PubMed]
- Rose, V.; Izukawa, T.; Moes, C.A.F. Syndromes of asplenia and polysplenia. Br. Heart J. 1975, 37, 840–852. [Google Scholar] [CrossRef] [PubMed]
- Heinemann, M.K.; Hanley, F.L.; Van Praagh, S.; Fenton, K.N.; Jonas, R.A.; Mayer, J.E.; Castaneda, A.R. Total anomalous venous drainage in newborns with visceral heterotaxy. Ann. Thorac. Surg. 1994, 57, 88–91. [Google Scholar] [CrossRef]
- Kitamura, K.; Miura, H.; Miyagawa-Tomita, S.; Yanazawa, M.; Katoh-Fukui, Y.; Suzuki, R.; Ohuchi, H.; Suehiro, A.; Motegi, Y.; Nakahara, Y.; et al. Mouse Pitx2 deficiency leads to anomalies of the ventral body wall, heart, extra- and periocular mesoderm, and right pulmonary isomerism. Development 1999, 126, 5749–5758. [Google Scholar] [PubMed]
- Liu, C.; Liu, W.; Palie, J.; Lu, M.F.; Brown, N.A.; Martin, J.F. Pitx2c patterns anterior myocardium and aortic arch vessels and is required for local cell movement into atrioventricular cushions. Development 2002, 129, 5081–5091. [Google Scholar] [PubMed]
- Casey, B.; Hackett, B.P. Left-right axis malformations in man and mouse. Curr. Opin. Genet. Dev. 2000, 10, 257–261. [Google Scholar] [CrossRef]
- Garcia-Martinez, V.; Schoenwolf, G.C. Primitive streak origin of the cardiovascular system in avian embryos. Dev. Biol. 1993, 159, 706–719. [Google Scholar] [CrossRef] [PubMed]
- Sadler, T.W. Cardiovascular system. In Langman’s Medical Embryology; Wolters Kluwer: Philadelphia, PA, USA, 2015; pp. 175–217. ISBN 978-1-4511-9164-6. [Google Scholar]
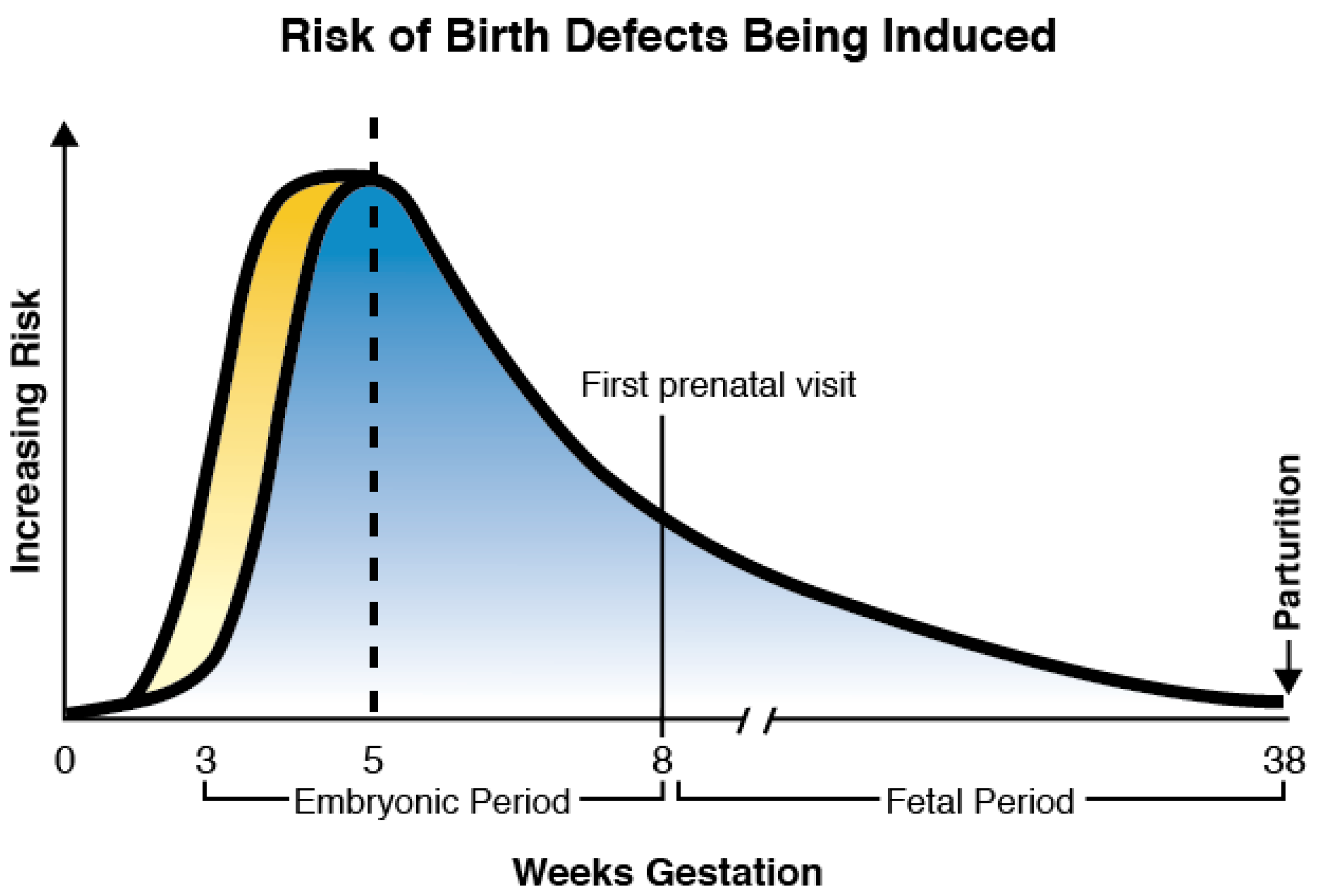

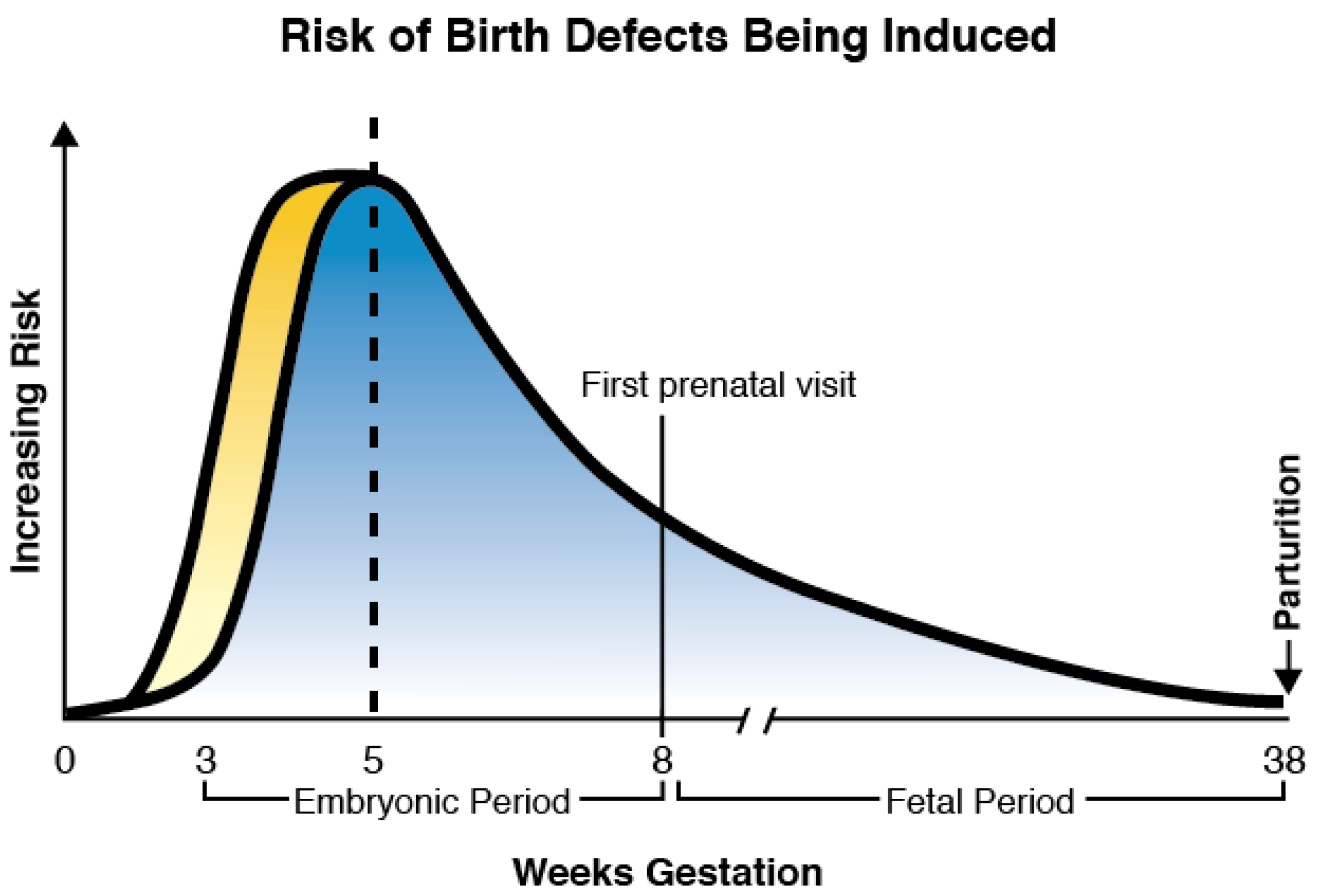{kind=link}
| Skeletal Defects | Caudal Dysgenesis |
|---|---|
| Limb defects (especially clubfoot) | Intestinal malrotation |
| Craniosynostosis | Kidney defects (agenesis, horseshoe) |
| Anal atresia | Tracheoesophageal fistula/Esophageal atresia |
| Imperforate anus | Genital defects |
| Gastroschisis | Micropthalmia |
| Omphalocele | Cleft lip and/or cleft palate |
| Neural tube defects | Vertebral anomalies |
| Dextrocardia | Pulmonary Valve Atresia and Stenosis |
|---|---|
| Single atrium | Hypoplastic left heart syndrome |
| Single ventricle | Double outlet right ventricle |
| Transposition of great arteries | Complete atrioventricular block |
| Total and partial anomulous pulomary venous return | Tetralogy of Fallot |
| Atrioventricular canal defect | Aortic Arch defects |
| Endocardial cushion defects | Ventricular inversion |
| Atrial septal defect | Atrial inversion |
| Ventricular septal defect | Atrial isomerism |
| Coarctation of the aorta | Double inlet left ventricle |
© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sadler, T.W. Establishing the Embryonic Axes: Prime Time for Teratogenic Insults. J. Cardiovasc. Dev. Dis. 2017, 4, 15. https://doi.org/10.3390/jcdd4030015
Sadler TW. Establishing the Embryonic Axes: Prime Time for Teratogenic Insults. Journal of Cardiovascular Development and Disease. 2017; 4(3):15. https://doi.org/10.3390/jcdd4030015
Chicago/Turabian StyleSadler, Thomas W. 2017. "Establishing the Embryonic Axes: Prime Time for Teratogenic Insults" Journal of Cardiovascular Development and Disease 4, no. 3: 15. https://doi.org/10.3390/jcdd4030015
APA StyleSadler, T. W. (2017). Establishing the Embryonic Axes: Prime Time for Teratogenic Insults. Journal of Cardiovascular Development and Disease, 4(3), 15. https://doi.org/10.3390/jcdd4030015