Abstract
Cardiac development is a complex morphogenetic process initiated as bilateral cardiogenic mesoderm is specified at both sides of the gastrulating embryo. Soon thereafter, these cardiogenic cells fuse at the embryonic midline configuring a symmetrical linear cardiac tube. Left/right bilateral asymmetry is first detected in the forming heart as the cardiac tube bends to the right, and subsequently, atrial and ventricular chambers develop. Molecular signals emanating from the node confer distinct left/right signalling pathways that ultimately lead to activation of the homeobox transcription factor Pitx2 in the left side of distinct embryonic organ anlagen, including the developing heart. Asymmetric expression of Pitx2 has therefore been reported during different cardiac developmental stages, and genetic deletion of Pitx2 provided evidence of key regulatory roles of this transcription factor during cardiogenesis and thus congenital heart diseases. More recently, impaired Pitx2 function has also been linked to arrhythmogenic processes, providing novel roles in the adult heart. In this manuscript, we provide a state-of-the-art review of the fundamental roles of Pitx2 during cardiogenesis, arrhythmogenesis and its contribution to congenital heart diseases.
1. Introduction to Cardiac Development
Cardiac development is an intricate morphogenetic process involving distinct cell lineages. The first steps of cardiogenic formation are initiated as bilateral precardiogenic mesoderm becomes specified []. Soon thereafter, the bilateral cardiogenic subpopulations merge at the embryonic midline configuring a linear cardiac tube [,] (Figure 1). At this stage of development, the heart is symmetrical, but soon thereafter, bilateral asymmetry is established as the cardiac tube bulges on its ventral aspect and then invariably bends rightward. Prospective embryonic atrial and ventricular chambers become soon established with a common inlet, atrioventricular canal and outlet segments [] (Figure 1). These cardiac regions are progressively derived from distinct cardiogenic cell subpopulations, the first heart field (FHF) and the second heart field (SHF) (Figure 1). As development proceeds, all cardiac chambers and segments are separated into systemic and venous components by the formation of a series of cardiac septa []. Inflow tract and atrial chambers are divided into right and left components by the subsequent formation of the primary and secondary atrial septa []. Ventricular chambers are separated by the coordinated formation of interventricular muscular and membranous septa, and distinct aortic and pulmonary trunks are divided with the formation of the outflow tract septum [] (Figure 1). Defects or minimal delays occurring during these morphogenetic processes lead to severe cardiac congenital malformations such as double inlet left ventricle (DILV), double outlet right ventricle (DORV), atrial or atrioventricular septal defects (ASD, AVSD), among others.
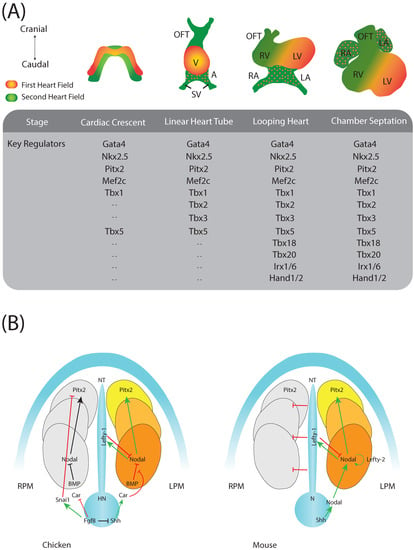
Figure 1.
(A) Schematic representation of the developing heart delineating distinct developmental stages and the most prominent transcription factors involved in these morphogenetic processes. Adapted from Paige et al. []; (B) Schematic representation of the molecular left/right signalling pathway emanating from the node and leading to the activation of the homeobox transcription factor Pitx2 in the left lateral plate mesoderm (LPM) in chicken and mouse, respectively. Images adapted from Meyers & Martin [] and Schlueter and Brand []. N, node; NT, notochord; HN, Hensen’s node; RPM, right lateral plate mesoderm; OFT, outflow tract; V, Ventricle; A, Atria; SV, sinus venosus; RV, right ventricle; RA, right atrium, LA, left atrium; LV, left ventricle.
While it is rather simple to ascribe embryonic right and left components to the initial symmetrical cardiac tube [], the contribution of the mesoderm originally patterned by the left/right axis to the developing structures is far more complicated and puzzling, a puzzle that remains to be fully elucidated. Nonetheless, a clear assumption can be drawn from this evidence, i.e., disruption of left/right signalling will have an important contribution to congenital heart diseases [,,].
2. Cardiac Regulatory Mechanisms
Over the last decade, our understanding of the molecular regulatory mechanisms underlying cardiovascular development has greatly advanced. Seminal papers reported key roles of distinct growth factors [,,] and transcription factors [] during early phases of cardiogenesis. Among those transcription factors, Nkx2.5, Mef2, Gata4 and Srf can be defined as core cardiogenic regulatory molecules [] (Figure 1). In fact, reprogramming of skin fibroblasts into cardiomyocyte underlies the functional importance of some of these transcription factors (Mef2, Gata4) [,]. In addition to these “global” cardiac-enriched transcription factors, a large array of tissue-restricted transcription factors has been identified during cardiogenesis. Hand1 and Hand2 transcription factors display ventricular chamber-specific expression in the systemic and venous components []. Tbx1 is restricted to the arterial pole with minimal contribution to the developing myocardial component, but essential for aortic arch formation and remodelling []. Tbx2 is confined to discrete areas of the developing heart, such as the outflow tract and atrioventricular canal, a pattern rather similar to Tbx3, contributing thereafter to some of the prospective cardiac conduction system components []. Tbx5 is restricted to the systemic ventricle, as well as both atrial chambers [,], whereas Tbx18 is confined to a discrete subset of ventricular cardiomyocytes, as well as to the venous pole of the heart []. Similarly, the Iroquois family members (Irx1 to Irx6) also display tissue-specific expression and thus functional roles during cardiogenesis [,,,]. Overall, these data demonstrate a complex orchestration of temporal and tissue-specific transcriptional regulation during cardiac development.
A particular case is the homeobox transcription factor Pitx2. Pitx2 expression is already present in the developing embryo soon after gastrulation [,,,]. Pitx2 subsequently acquires a tissue-restricted pattern in the early symmetrical organ anlages, including the heart, and its expression continues to be modified as organogenesis takes place, as we will see in the following chapters.
3. The Homeobox Transcription Factor Pitx2
Homeobox genes encode transcription factors that bind to cognate DNA sequences through their conserved homeodomain regions []. Pitx1, Pitx2 and Pitx3 are three vertebrate paralogues from the Pitx family []. The Pitx proteins belong to the bicoid-related subclass of homeodomain proteins with the lysine residue at position nine of the third helix, being the major determinant of DNA and RNA binding specificity [], and are highly conserved at the amino acid level at the C-terminal while significantly diverging at the N-terminal [].
The Pitx family is crucial for the formation of several tissues. Pitx1 is involved in development and specification of a number of structures including first branchial arch derivatives, body wall and hind limb musculature [,,,], whereas Pitx2 is fundamental to brain, pituitary, facial structures and heart development [,,,,,]. Pitx3 is the third gene of the family, and it is central to eye and dopaminergic neurone development [,,].
The Pitx2 transcription factor is widely distributed in different species [] and is encoded by the Pitx2 gene, which is located on chromosome 3 (3G3; 3 57.84 cM) in the mouse []. This transcription factor displays three different isoforms, which are generated by alternative splicing with a common promoter for Pitx2a and Pitx2b located upstream of exon 1, whereas Pitx2c uses an alternative promoter located upstream of exon 4 and a left side-specific enhancer (ASE) upstream of exon 6 [,,,,,,] (Figure 2). These three isoforms share exons 5 and 6, but differ in others; exon 3 is Pitx2b specific, while exon 4 is Pitx2c specific. Different laboratories assessed by genetic and/or epigenetic studies and Pitx2 knockout mice that this gene is required for the proper development of several organs and structures, such as the pituitary gland, craniofacial region, eyes, heart, abdominal viscera and limbs [,,,,,,,,,,,]. In human, PITX2 was identified by positional cloning of the 4q25 locus in patients with Axenfeld-Rieger syndrome []; a syndromic condition coursing with cardiovascular outflow tract malformations, eye dysmorphogenesis, craniofacial and pituitary abnormalities. Furthermore, Cox et al. [] reported a new PITX2 isoform (PITX2D) in humans generated by the PITX2C alternative promoter and differential splicing, acting as a dominant negative isoform, specifically against PITX2A and PITX2C (Figure 2). PITX2 mutations are associated to Axenfeld-Rieger syndrome [,,,] and several cardiac defects, as we will detail below.
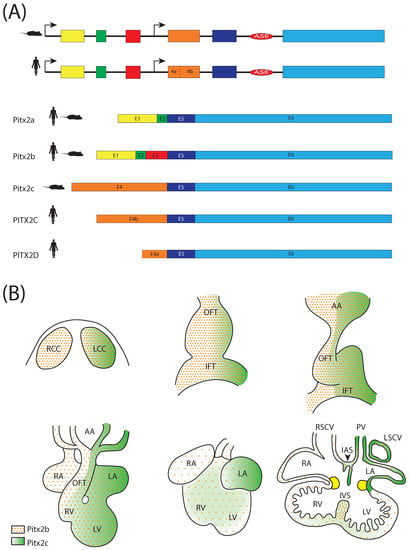
Figure 2.
(A) Schematic representation of the Pitx2 genomic structure and the corresponding Pitx2 isoforms in humans and mice; (B) Schematic representation of the Pitx2 isoform (Pitx2b and Pitx2c) expression patterns during cardiac development.
4. Pitx2 and Left-Right Signalling
During the early steps of gastrulation, the developing mouse and human embryo are symmetrical. Soon after gastrulation, the first signs of bilateral asymmetry are observed as the developing heart bulges ventrally and then bends to the right. Genetic analyses have demonstrated that morphological asymmetry is preceded by molecular asymmetry [,,], Several molecules have been identified in different species to be asymmetrically expressed during gastrulation such as lefty-2 in Xenopus and mice [,,,,,,], Nodal in Amphioxus, Xenopus, zebrafish, rat and mice [,,,,], cryptic in mice [], Charon in zebrafish [], Fgf8 and Hh in chicken and mice [,], Cerl-2 in mice [,,,] and SnR in chicken []. Importantly, several of these asymmetric regulators display left/right differential expression in the lateral plate mesoderm (LPM), while others already display asymmetric expression in the embryonic midline left/right organizer (LRO), within the developing node. In fact, differential expression in the node [] and directional flow mediated by cilia in the node [,,,,] have been extensively reported to be governing left/right symmetry break. In addition, cell movements within the node have also been implicated in left/right asymmetry []. However, two important conclusions can be drawn from all of these datasets; first, the node plays a fundamental role in establishing left/right symmetry break and particularly node cilia and node cilia-related flow (with some exceptions such as the chick and the pig); and second, the left/right symmetric break is a conserved morphogenetic process in most vertebrate phyla having a core signalling cascade represented by nodal > Pitx2 with different variable players in distinct species (Figure 1). Importantly, Pitx2 is therefore the last effector of the left/right signalling cascade transmitting positional information from the uncommitted LPM to distinct organ primordia such as the heart, lung and gut, among others [,,], leading to distinct sidedness alterations within these organs if impaired [,,,,,,].
Thus, the left LPM provides different signals controlling embryonic left-right asymmetry. Several laboratories have shown that Pitx2 expression is confined to left LPM in chick, frog and mouse [,,,,,], as well as the cardiogenic precursors of the left SHF []. During development, Pitx2 continues to be expressed asymmetrically in several organs. Over-expression studies in Xenopus and chicken demonstrate that Pitx2 is important in the determination of vertebrate heart and gut looping [,,,,,].
Pitx2 isoforms distinctly contribute to the regulation of left-right asymmetry [,,]. Pitx2a, Pitx2b and Pitx2c are symmetrically expressed in the head at the beginning of development, but Pitx2c is only expressed asymmetrically in the LPM from two to four somite stages in the mouse. During development and adulthood, Pitx2c expression still remains in the developing heart [,,] (Figure 2). This expression pattern is highly conserved among species, highlighting the role of Pitx2 during development [,,,,,]. Experimental models of gain and loss of function demonstrated that Pitx2 is dispensable in driving looping directionality in mice [,,,,,,], contrasting with those findings in other experimental models [,,,,,]. To date, such discrepancies remain to be fully elucidated.
5. Genetics of Congenital Heart Diseases
Congenital heart disease (CHD) is the most prevalent developmental abnormality in humans. CHD is indeed the most common non-infectious cause of infant morbidity and mortality. Increasing evidence has demonstrated a crucial contribution of genetic defects in the pathogenesis of CHD. Mutations in several transcription factors such as HAND1, HAND2, TBX5, GATA4, NKX2.5 and TBX20 have been identified in patients affected with distinct congenital heart diseases, such as atrial septal defects (TBX5, GATA4, NKX2.5, TBX20), Tetralogy of Fallot (TBX5, TBX20), ventricular septal defects (HAND2), double outlet right ventricle (HAND1), either syndromic (Holt-Oram; TBX5) or isolated (see for recent reviews [,]). However, CHDs are genetically heterogeneous, and at present, the genetic determinants of CHD in most patients remain yet unknown. We provide herein current state-of-the-art evidence on the contribution of the homeobox transcription factor PITX2 as a genetic determinant of congenital heart diseases.
6. Left-Right Signalling and Congenital Heart Diseases
The adult heart is an asymmetric organ. It is topologically located outside the body midline with the apex oriented to the left. Morphologically, the atrial chambers are rather similar, but conspicuous differences, such as the extent of the smooth region and the abundance of the pectinate muscles, provide hallmarks for distinguishing right and left atrial appendages []. Such left/right atrial differences are more easily assessed if their venous connections are taken into account. In humans, four pulmonary veins drain into the left atrium, whereas two caval veins (right superior and inferior caval veins) and the coronary sinus drain into the right atrium. In mice and other rodent species, pulmonary veins drain with a single orifice in the left atrium, whereas three caval veins drain into the right atrium. Atrioventricular valves are also left/right asymmetric, as the mitral valve has two leaflets and the tricuspid valve three. At the arterial pole, including the ventricles, asymmetry is also evident, as the left ventricle is larger than the right ventricle, and the aortic and pulmonary outlets are displaced from the body midline [,]; yet, these differences do not reflect in most cases embryonic left/right differences. Importantly, embryonic left/right contribution to the atria and ventricles is markedly different as reported by Pitx2 expression [,,].
Several cardiac abnormalities have been classically reported as defects of the establishment of the left/right body axis (laterality defects). Right atrial isomerism (RAI) is the most common defect in which the heart is formed by isomeric right atria and impaired venous return connections, displaying in most cases the absence of direct pulmonary vein drainage to the atria and bilateral caval veins connections. In this context, both atria display a right-specific pattern; i.e., morphologically right atrial appendages. Left atrial isomerism (LAI) is rather less frequent, presenting isomeric left atria with the absence of direct caval vein drainage to the atria and bilateral pulmonary vein connections, i.e., right pulmonary vein to the right atrium and left pulmonary veins to the left atrium. Similarly LAI also displays a left-specific pattern in both atria; i.e., with morphologically left atrial appendages. These cardiac alterations can occur with an impaired left/right topology of all other body organs, a condition dubbed situs inversus, or not, a condition dubbed situs ambiguus. Several lines of experimental evidence demonstrated that impaired left/right signalling underlies RAI and LAI.
The plausible contribution of left/right signalling to other congenital heart defects such as double outlet right ventricle (DORV), double inlet left ventricle (DILV), atrial or atrioventricular septal defects (ASD and AVSD) remains enigmatic, as well as in syndromic congenital heart defects such as Tetralogy of Fallot. Thus, an important question is when is cardiac asymmetry established during embryonic development and which is the particular contribution of left/right cardiac embryonic tissues to the adult heart. We will provide herein state-of-the-art evidence on the plausible role of left/right signaling, with particular emphasis on the homeobox transcription factor Pitx2, on the genesis of these cardiac congenital defects.
7. Pitx2 and Congenital Heart Diseases
Systemic deletion of Pitx2 in experimental models revealed complex embryonic defects. Abnormal sidedness was observed in distinct developing organs such as the lungs, gut and heart in mice. Embryonic development was compromised at Embryonic Days (ED) 13–14. Within the cardiovascular system, Pitx2 null mutants displayed right atrial isomerism with impaired venous return [,,]. Pulmonary vein development was severely impaired as was the sinoatrial node development [,]. Isoform-specific deletion of Pitx2c in mice demonstrated that it was mainly this isoform that was responsible for all of the cardiovascular embryonic defects []. Furthermore, conditional deletion in mice highlighted the importance of Pitx2 driving cardiac left-right asymmetry from the secondary heart field [] and opened up the possibility of spatio-temporally-controlled deletion of Pitx2 in distinct cardiac compartments. Tessari et al. [] conditionally ablated Pitx2 function in the early embryonic cardiomyocytes, and surprisingly, right/left cardiac asymmetry was unaltered. Atrial and ventricular-specific deletion in mouse embryos [], as well as temporal deletion in postnatal hearts [,], displayed similar findings. Only when Pitx2 deletion was carried out in the nascent cardiomyocytes, right atrial isomerism was observed []. Overall, these data demonstrate a highly controlled temporal and tissue-specific action of Pitx2 during cardiac development that, if impaired, can lead to distinct cardiac congenital heart diseases. Therefore, these observations postulate Pitx2 as a candidate gene for cardiac congenital heart diseases (Figure 3).
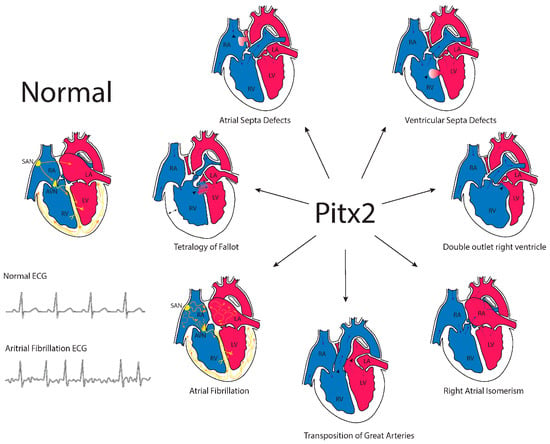
Figure 3.
Schematic representation of the contribution of the homeobox transcription factor PITX2 to distinct congenital heart diseases and cardiac arrhythmogenic defects.
CHARGE syndrome is characterized by ocular Coloboma, congenital Heart defects, Atresia of the choanae, Retarded growth, Genital hypoplasia and Ear anomalies including deafness in the context of global developmental delay. Initial screening in CHARGE syndrome patients discarded PITX2 as a causing gene []. Subsequent screening in a small cohort of transposition of great arteries (TGA) patients similarly failed to identify PITX2 causal mutations []. More recently, PITX2 point mutations have been identified in distinct isolated cardiac congenital heart diseases [], such as atrial septal defects (ASD) [], TGA [], ventricular septal defect (VSD) [,] and Tetralogy of Fallot []. A PITX2 mutation has also been identified in a patient with compound congenital heart disease (CHD), i.e., DORV and VSD. In all cases, functional analyses of their corresponding PITX2 mutation leads to decreased transcriptional activity and reduced synergistic activation with NKX2.5 [,,,] (Table 1). Furthermore, point mutations in CITED2, identified in a Tetralogy of Fallot patient with aortic stenosis, impaired PITX2 and VEGF expression [], further support a functional role for PITX2 in this type of CHD. In addition to the identification of PITX2 mutations in isolated CHD, a loss-of-function PITX2 mutation has been identified in an Axenfeld-Rieger syndrome patient carrying cardiac endocardial cushion defects. Such a PITX2 mutation provides no transcriptional and synergistic activity with Nkx2.5 []. Overall, these data demonstrate a pivotal functional role of PITX2 in cardiac congenital heart diseases (Figure 3).

Table 1.
Summary of the distinct cardiac congenital heart diseases associated to Pitx2.
8. Beyond CHD: Pitx2 and Atrial Fibrillation
Atrial fibrillation (AF) is the most common cause of arrhythmogenesis, and it has been related to several risk factors, such as advanced age, male gender, hypertension, obesity, ischaemic heart disease, myocardial infarction, valvular diseases and hyperthyroidism [,,].
Several genome-wide association studies (GWAS) have been published reporting chromosomal loci in association with atrial fibrillation (AF). Four different genetic loci on chromosomes 1q21-KCNN3, 4q25-Pitx2, 16q22-ZFHX3 and 16q13-IL6R have been associated with this arrhythmia [,,,,]. GWAS meta-analyses have provided additional risk variants associated with AF, implicating six new loci in AF; CAV1, HCN4, SYNE2, SYNPO2L, PRRX1 and WNT8A [,,], but importantly, the most relevant associated risk variants are those located in the vicinity of the PITX2 locus [,,,,,,,]. Thus, this seminal GWAS study by Gudbjartsson et al. [], identifying single nucleotide polymorphisms (SNPs) at the 4q25 locus, proposed an association between PITX2 and AF, as a causative molecular link. Thus, a novel role for Pitx2 in the adult heart has therefore emerged.
Distinct laboratories, including ours, have demonstrated that PITX2 potentially regulates AF through modulation of multiple genes implicated in AF; KCNN3 [,,]; TBX5 [,,]; HCN4 [,]; KCNJ2 [,]; CAV-1 [,]; KCNQ1 [,]; ZFHX3; SYNE2; Il6R []; ENPEP []. In addition, a transcriptional network that linked AF risk loci has been recently described. Tbx5 directly activated Pitx2, and Tbx5 and Pitx2 antagonistically regulated membrane effector genes Scn5a, Gja1, Ryr2, Dsp and Atp2a2 [] (Table 2).
Table 2.
Summary of distinct signalling pathways regulated by Pitx2 and currently associated with atrial fibrillation.
Genomic regions containing the risk variants have been reported to play a role in Pitx2 regulation in a tissue- and isoform-specific fashion [,], and a functional role for Pitx2 in atrial arrhythmias using distinct experimental models has been demonstrated [,,]. Experimental studies have demonstrated that Pitx2 loss of function predisposes to atrial arrhythmogenesis. Mice heterozygous for Pitx2 are susceptible to AF during programmed stimulation []. Pitx2 haploinsufficiency predisposes to AF in electrically-stimulated adult mice, provoked by ectopic Shox2 expression in the foetal left atrium (LA), which in turn deregulated other pivotal sino-atrial node genes such as Hcn4 and Tbx3 [].
Other substrates for AF are left-sided electrophysiological defects generated by Pitx2 impairment in atrium-specific conditional Pitx2 mouse mutants []. In this model, in foetal and adult stages, voltage-gated sodium and inward rectifying potassium channels are abnormally expressed in the atrial myocardium []. Moreover, dysregulation of ion channels resulting from impaired Pitx2 dosage has been reported in adult Pitx2 heterozygous mutant mice []. In Pitx2c heterozygous mutant mice, the expression of TWIK-related acid-sensitive K+ channel (TASK-2) and TASK-like background currents are reduced, although IK1 is not altered []. Impaired gap junction expression in adult heterozygous Pitx2 mutant mice has been reported [], being in concordance with the fact that Pitx2 controls Gja5 expression [,] and with reports of mutations in gap junctional proteins in the context of AF ([]). Surprisingly, chronic AF patients display upregulation of PITX2C, contributing to IKs increase and ICa,L reduction []. While these controversial findings remain to be elucidated, currently we can conclude that impairing Pitx2 (upregulation or downregulation) is critical for cardiac electrophysiology and, thus, tightly linked to the onset of cardiac arrhythmias.
Our laboratory recently reported that Pitx2 insufficiency also regulates WNT8 expression, which modulates a complex gene regulatory network, including multiple microRNAs, with a large impact on calcium homeostasis control and pro-arrhythmogenic events []. Multiple microRNAs are downstream from Pitx2 and involved in AF. ChiP-sequencing studies have reported that Pitx2 positively regulates miR-17-92 and its related homologue miR-106b-25. Deficient mice in these miRNAs clusters show arrhythmia susceptibility and dysregulation of Shox2 and Tbx3, similar characteristics to Pitx2-deficient mice []. IK1 channel expression is modulated by miR-1 [], which is upregulated in Pitx2 atrial-deficient mice []. A significant number of microRNAs differentially expressed in AF patients are regulated by Pitx2 and Wnt signalling, such as miR-1, miR-26b, miR-29a, miR-106a/b and miR-133 [,].
However, a question that remains unsolved is whether impaired Pitx2 function already alters cardiac function during embryogenesis, as suggested by Wang et al. [], or whether it is mainly an adulthood deficiency that is causing atrial fibrillation, as suggested by Chinchilla et al. []. We demonstrated herein that atrial-specific Pitx2-insufficient embryonic (ED18.5) mice showed arrhythmogenic substrates already at these early stages in homozygous knockout mutants. These animals showed prolonged atrial activation time (Figure 4) under the sinus rhythm. Similarly, slower impulse propagation was noted also in the electrically-stimulated beats, both in the left and right atrial appendages (Figure 4). Interestingly, there was no significant morphological substrate present at that point, as the amount of collagen, detected by Picrosirius red staining, was uniformly low and not different between controls and mutants. Furthermore, connexin43 expression in the atria did not differ between controls and mutants. These results therefore suggest that functional changes in atrial-specific Pitx2-insufficient mice pre-date structural differences. We therefore propose herein a reconciling hypothesis. Pitx2 insufficiency already predisposes from embryonic stages to atrial electrical impairment in the absence of structural alterations. Onset of spontaneous arrhythmogenic events might be triggered by additional AF risk factors such atrial dilation [] or by progressive Pitx2 downregulation that can course with age and hypertension []. Furthermore, the first arrhythmogenetic events also lead to Pitx2 downregulation, subsequently predisposing to the onset of additional arrhythmogenetic events [].
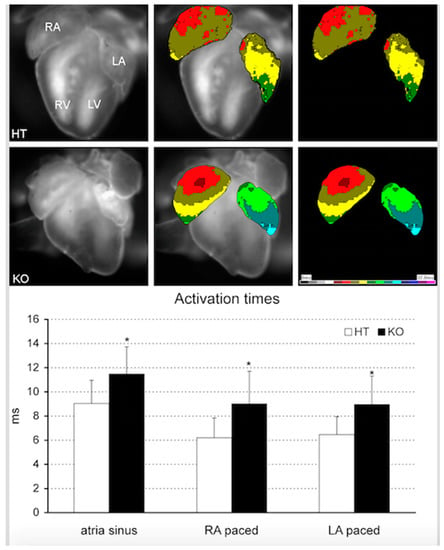
Figure 4.
Optical mapping analyses of embryonic heterozygous (HT) and homozygous (KO) atrial-specific Pitx2 mutants. Observe that atrial activation times are significantly slower in homozygous as compared to heterozygous hearts. In addition, right and left atrial pacing, respectively display similarly slower activation times, demonstrating an inherent electrical difference between heterozygous (HT) and homozygous (KO) atrial-specific Pitx2 mutants.
9. Conclusions and Future Perspectives
Congenital heart diseases are multifactorial. Over the last few decades, we have witnessed a large advance in the understanding of the genetics of congenital heart diseases. Mutations in different transcription factors have been identified in familial and sporadic cases of distinct congenital heart diseases. Genetic engineering in mice has further provided additional evidence of the functional role of these transcription factors (and their corresponding mutations) in CHDs. Herein, we have provided a state-of-the-art review of the contribution of the homeobox transcription factor Pitx2 to congenital heart diseases. Pitx2 participates in early embryonic left/right signalling and cardiac embryogenesis, and it remains to be expressed in the adult heart. Therefore, the contribution of Pitx2 to CHDs might involve distinct roles at different developmental stages. In this line of thinking, Fakhro et al. [] reported a genetic screening of rare copy number variations in heterotaxia patients and identified genes involved in cilia function. Experimental manipulation of these cilia-related genes led to impaired Pitx2 expression in Xenopus experimental models, providing functional links between cilia, left/right, Pitx2 and heterotaxia. Distinct genetic Pitx2 manipulations have also reported distinct congenital heart diseases, establishing temporal and tissue-specific contributions [,,]. More recently, a link between Pitx2 and atrial fibrillation has been reported [,,]. Our data and others demonstrated that electrical alterations underlying Pitx2 insufficiency are already present at foetal stages, suggesting therefore a congenital substrate on the future onset of atrial arrhythmogenesis. Thus, whether AF might be considered as a developmental disease should be debated.
In addition to experimental models demonstrating the functional role of Pitx2 in CHDs, emerging evidence also identified point mutations associated with distinct CHDs, ranging from isolated atrial septal defects to complex cases of Tetralogy of Fallot. At present, evidence of the functional role of these point mutations in these CHDs is rudimentary. CRISPR/Cas9 gene editing of these mutations might in future shed light on the specific contribution of Pitx2 mutations to CHDs.
Acknowledgments
This work is partially supported by a Junta de Andalucia (Spain) grant-in-aid to D.F.
Author Contributions
D.F. and D.S. conceived of and designed the experiments. D.S. performed the experiments and analysed the data. D.F. and E.L.-V. wrote the paper. E.L.-V. and D.S. carefully edited the final version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Moorman, A.F.; Christoffels, V.M. Cardiac chamber formation: Development, genes, and evolution. Physiol. Rev. 2003, 83, 1223–1267. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Martinez, V.; Schoenwolf, G.C. Positional control of mesoderm movement and fate during avian gastrulation and neurulation. Dev. Dyn. 1992, 193, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Martinez, V.; Schoenwolf, G.C. Primitive-streak origin of the cardiovascular system in avian embryos. Dev. Biol. 1993, 159, 706–719. [Google Scholar] [CrossRef] [PubMed]
- Franco, D.; Campione, M.; Kelly, R.; Zammit, P.S.; Buckingham, M.; Lamers, W.H.; Moorman, A.F. Multiple transcriptional domains, with distinct left and right components, in the atrial chambers of the developing heart. Circ. Res. 2000, 87, 984–991. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.H.; Webb, S.; Brown, N.A.; Lamers, W.; Moorman, A. Development of the heart: (3) Formation of the ventricular outflow tracts, arterial valves, and intrapericardial arterial trunks. Heart 2003, 89, 1110–1118. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.H.; Webb, S.; Brown, N.A.; Lamers, W.; Moorman, A. Development of the heart: (2) Septation of the atriums and ventricles. Heart 2003, 89, 949–958. [Google Scholar] [CrossRef] [PubMed]
- Campione, M.; Ros, M.A.; Icardo, J.M.; Piedra, E.; Christoffels, V.M.; Schweickert, A.; Blum, M.; Franco, D.; Moorman, A.F. Pitx2 expression defines a left cardiac lineage of cells: Evidence for atrial and ventricular molecular isomerisms in the iv/iv mice. Dev. Biol. 2001, 231, 252–264. [Google Scholar] [CrossRef] [PubMed]
- Kathiriya, I.S.; Srivastava, D. Left-right asymmetry and cardiac looping: Implications for cardiac development and congenital heart disease. Am. J. Med. Genet. 2000, 97, 271–279. [Google Scholar] [CrossRef]
- Franco, D.; Campione, M. The role of Pitx2 during cardiac development: Linking left-right signaling and congenital heart diseases. Trends Cardiovasc. Med. 2003, 13, 157–163. [Google Scholar] [CrossRef]
- Campione, M.; Acosta, L.; Martínez, S.; Icardo, J.M.; Aránega, A.; Franco, D. Pitx2 and cardiac development: A molecular link between left/right signaling and congenital heart disease. Cold Spring Harb. Symp. Quant. Biol. 2002, 67, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Paige, S.L.; Plonowska, K.; Xu, A.; Wu, S.M. Molecular regulation of cardiomyocyte differentiation. Circ. Res. 2015, 116, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Meyers, E.N.; Martin, G.R. Differences in left-right axis pathways in mouse and chick: Functions of FGF8 and SHH. Science 1999, 285, 403–406. [Google Scholar] [CrossRef] [PubMed]
- Schlueter, J.; Brand, T. Left/right assymetrical development of the proepicardiu. J. Dev. Biol. 2013, 1, 126–140. [Google Scholar] [CrossRef]
- Lopez-Sanchez, C.; Climent, V.; Schoenwolf, G.C.; Alvarez, I.S.; Garcia-Martinez, V. Induction of cardiogenesis by Hensen's node and fibroblast growth factors. Cell Tissue Res. 2002, 309, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Andrée, B.; Duprez, D.; Vorbusch, B.; Arnold, H.H.; Brand, T. BMP-2 induces ectopic expression of cardiac lineage markers and interferes with somite formation in chicken embryos. Mech. Dev. 1998, 70, 119–131. [Google Scholar] [CrossRef]
- Ladd, A.N.; Yatskievych, T.A.; Antin, P.B. Regulation of avian cardiac myogenesis by activin/TGFbeta and bone morphogenetic proteins. Dev. Biol. 1998, 204, 407–419. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, D. HAND proteins: Molecular mediators of cardiac development and congenital heart disease. Trends Cardiovasc. Med. 1999, 9, 11–18. [Google Scholar] [CrossRef]
- Schlesinger, J.; Schueler, M.; Grunert, M.; Fischer, J.J.; Zhang, Q.; Krueger, T.; Lange, M.; Tönjes, M.; Dunkel, I.; Sperling, S.R. The cardiac transcription network modulated by Gata4, Mef2a, Nkx2.5, Srf, histone modifications, and microRNAs. PLoS Genet. 2011, 7, e1001313. [Google Scholar] [CrossRef] [PubMed]
- Ieda, M.; Fu, J.D.; Delgado-Olguin, P.; Vedantham, V.; Hayashi, Y.; Bruneau, B.G.; Srivastava, D. Direct reprogramming of fibroblasts into functional cardiomyocytes by defined factors. Cell 2010, 142, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.D.; Stone, N.R.; Liu, L.; Spencer, C.I.; Qian, L.; Hayashi, Y.; Delgado-Olguin, P.; Ding, S.; Bruneau, B.G.; Srivastava, D. Direct reprogramming of human fibroblasts toward a cardiomyocyte-like state. Stem Cell Rep. 2013, 1, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, D. Making or breaking the heart: From lineage determination to morphogenesis. Cell 2006, 126, 1037–1048. [Google Scholar] [CrossRef] [PubMed]
- Baldini, A.; Fulcoli, F.G.; Illingworth, E. Tbx1: Transcriptional and Developmental Functions. Curr. Top. Dev. Biol. 2017, 122, 223–243. [Google Scholar] [PubMed]
- Greulich, F.; Rudat, C.; Kispert, A. Mechanisms of T-box gene function in the developing heart. Cardiovasc. Res. 2011, 91, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Mori, A.D.; Bruneau, B.G. TBX5 mutations and congenital heart disease: Holt-Oram syndrome revealed. Curr. Opin. Cardiol. 2004, 19, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Steimle, J.D.; Moskowitz, I.P. TBX5: A Key Regulator of Heart Development. Curr. Top. Dev. Biol. 2017, 122, 195–221. [Google Scholar] [PubMed]
- Christoffels, V.M.; Keijser, A.G.; Houweling, A.C.; Clout, D.E.; Moorman, A.F. Patterning the embryonic heart: Identification of five mouse Iroquois homeobox genes in the developing heart. Dev. Biol. 2000, 224, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Houweling, A.C.; Dildrop, R.; Peters, T.; Mummenhoff, J.; Moorman, A.F.; Rüther, U.; Christoffels, V.M. Gene and cluster-specific expression of the Iroquois family members during mouse development. Mech. Dev. 2001, 107, 169–174. [Google Scholar] [CrossRef]
- Bruneau, B.G.; Bao, Z.Z.; Fatkin, D.; Xavier-Neto, J.; Georgakopoulos, D.; Maguire, C.T.; Berul, C.I.; Kass, D.A.; Kuroski-de Bold, M.L.; de Bold, A.J.; et al. Cardiomyopathy in Irx4-deficient mice is preceded by abnormal ventricular gene expression. Mol. Cell. Biol. 2001, 21, 1730–1736. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Rosen, A.; Bruneau, B.G.; Hui, C.C.; Backx, P.H. Iroquois homeodomain transcription factors in heart development and function. Circ. Res. 2012, 110, 1513–1524. [Google Scholar] [CrossRef] [PubMed]
- Ryan, A.K.; Blumberg, B.; Rodríguez-Esteban, C.; Yonei-Tamura, S.; Tamura, K.; Tsukui, T.; de la Peña, J.; Sabbagh, W.; Greenwald, J.; Choe, S.; et al. Pitx2 determines left-right asymmetry of internal organs in vertebrates. Nature 1998, 394, 545–551. [Google Scholar] [PubMed]
- Logan, M.; Pagán-Westphal, S.M.; Smith, D.M.; Paganessi, L.; Tabin, C.T. The transcription factor Pitx2 mediates situs-specific morphogenesis in response to left-right asymmetric signals. Cell 1998, 94, 307–317. [Google Scholar] [CrossRef]
- Piedra, M.E.; Icardo, J.M.; Albajar, M.; Rodriguez-Rey, J.C.; Ros, M.A. Pitx2 participates in the late phase of the pathway controlling left-right asymmetry. Cell 1998, 94, 319–324. [Google Scholar] [CrossRef]
- Campione, M.; Steinbeisser, H.; Schweickert, A.; Deissler, K.; van Bebber, F.; Lowe, L.A.; Nowotschin, S.; Viebahn, C.; Haffter, P.; Kuehn, M.R.; et al. The homeobox gene Pitx2: Mediator of asymmetric left-right signaling in vertebrate heart and gut looping. Development 1999, 126, 1225–1234. [Google Scholar] [PubMed]
- Scott, M.P.; Tamkun, J.W.; Hartzell, I.G.W. The structure and function of the homeodomain. Biochem. Biophys. Acta 1989, 989, 25–48. [Google Scholar] [CrossRef]
- Gage, P.J.; Suh, H.; Camper, S.A. The bicoid-related Pitx gene family in development. Mamm. Genome 1999, 10, 197–200. [Google Scholar] [CrossRef] [PubMed]
- Gehring, W.J.; Qian, Y.Q.; Billeter, M.; Furukubo-Tokunaga, K.; Schier, A.F.; Resendez-Perez, D.; Affolter, M.; Otting, G.; Wüthrich, K. Homeodo- main-DNA recognition. Cell 1994, 78, 211–223. [Google Scholar] [CrossRef]
- Gage, P.J.; Suh, H.; Camper, S.A. Dosage requirement of Pitx2 for development of multiple organs. Development 1999, 126, 4643–4651. [Google Scholar] [PubMed]
- Lanctot, C.; Lamolet, B.; Drouin, J. The bicoid-related homeoprotein Ptx1 defines the most anterior domain of the embryo and differentiates posterior from anterior lateral mesoderm. Development 1997, 124, 2807–2817. [Google Scholar] [PubMed]
- Logan, M.; Tabin, C.J. Role of Pitx1 upstream of Tbx4 in specification of hindlimb identity. Science 1999, 283, 1736–1739. [Google Scholar] [CrossRef] [PubMed]
- Szeto, D.P.; Rodriguez-Esteban, C.; Ryan, A.K.; O’Connell, S.M.; Liu, F.; Kioussi, C.; Gleiberman, A.S.; Izpisua-Belmonte, J.C.; Rosenfeld, M.G. Role of the Bicoid-related homeodomain factor Pitx1 in specifying hindlimb morphogenesis and pituitary development. Genes Dev. 1999, 13, 484–494. [Google Scholar] [CrossRef] [PubMed]
- Kioussi, C.; Briata, P.; Baek, S.H.; Rose, D.W.; Hamblet, N.S.; Herman, T.; Ohgi, K.A.; Lin, C.; Gleiberman, A.; Wang, J.; et al. Identification of a Wnt/Dvl/beta-Catenin → Pitx2 pathway mediating cell-type-specific proliferation during development. Cell 2002, 111, 673–985. [Google Scholar] [CrossRef]
- Kioussi, C.; Briata, P.; Baek, S.H.; Wynshaw-Boris, A.; Rose, D.W.; Rosenfeld, M.G. Pitx genes during cardiovascular development. Cold Spring Harb. Symp. Quant. Biol. 2002, 67, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, K.; Miura, H.; Miyagawa-Tomita, S.; Yanazawa, M.; Katoh-Fukui, Y.; Suzuki, R.; Ohuchi, H.; Suehiro, A.; Motegi, Y.; Nakahara, Y.; et al. Mouse Pitx2 deficiency leads to anomalies of the ventral body wall, heart, extra- and periocular mesoderm and right pulmonary isomerism. Development 1999, 126, 5749–5758. [Google Scholar] [PubMed]
- Kitamura, K.; Miura, H.; Yanazawa, M.; Miyashita, T.; Kato, K. Expression patterns of Brx1 (Rieg gene), Sonic hedgehog, Nkx2.2, Dlx1 and Arx during zona limitans intrathalamica and embryonic ventral lateral geniculate nuclear formation. Mech. Dev. 1997, 67, 83–96. [Google Scholar] [CrossRef]
- Lu, M.F.; Pressman, C.; Dyer, R.; Johnson, R.L.; Martin, J.F. Function of Rieger syndrome gene in left-right asymmetry and craniofacial development. Nature 1999, 401, 276–278. [Google Scholar] [PubMed]
- Muccielli, M.L.; Martinez, S.; Pattyn, A.; Goridis, C.; Brunet, J.F. Otlx2, an Otx-related homeobox gene expressed in the pituitary gland and in a restricted pattern in the forebrain. Mol. Cell. Neurosci. 1996, 8, 258–271. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.Y.; Guo, C.; Cheng, X.W.; Huang, Y.; Xiong, Z.Q.; Ding, Y.Q. Pitx3-CreER mice showing restricted Cre expression in developing ocular lens and skeletal muscles. Genesis 2008, 46, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Smidt, M.P.; Smits, S.M.; Bouwmeester, H.; Hamers, F.P.; van der Linden, A.J.; Hellemons, A.J.; Graw, J.; Burbach, J.P. Early developmental failure of substantia nigra dopamine neurons in mice lacking the homeodomain gene Pitx3. Development 2004, 131, 1145–1155. [Google Scholar] [CrossRef] [PubMed]
- Smidt, M.P.; van Schaick, H.S.; Lanctot, C.; Tremblay, J.J.; Cox, J.J.; van der Kleij, A.A.; Wolterink, G.; Drouin, J.; Burbach, J.P. A homeodomain gene Ptx3 has highly restricted brain expression in mesencephalic dopaminergic neurons. Proc. Natl. Acad. Sci. USA 1997, 94, 13305–13310. [Google Scholar] [CrossRef] [PubMed]
- Gage, P.J.; Camper, S.A. Pituitary homeobox 2, a novel member of the bicoid-related family of homeobox genes, is a potential regulator of anterior structure formation. Hum. Mol. Genet. 1997, 6, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Arakawa, H.; Nakamura, T.; Zhadanov, A.B.; Fidanza, V.; Yano, T.; Bullrich, F.; Shimizu, M.; Blechman, J.; Mazo, A.; Canaani, E.; et al. Identification and characterization of the ARP1 gene, a target for the human acute leukemia ALL1 gene. Proc. Natl. Acad. Sci. USA 1998, 95, 4573–4578. [Google Scholar] [CrossRef] [PubMed]
- Cox, C.J.; Espinoza, H.M.; McWilliams, B.; Chappell, K.; Morton, L.; Hjalt, T.A.; Semina, E.V.; Amendt, B.A. Differential regulation of gene expression by PITX2 isoforms. J. Biol. Chem. 2002, 277, 25001–25010. [Google Scholar] [CrossRef] [PubMed]
- Semina, E.V.; Reiter, R.; Leysens, N.J.; Alward, W.L.; Small, K.W.; Datson, N.A.; Siegel-Bartelt, J.; Bierke-Nelson, D.; Bitoun, P.; Zabel, B.U.; et al. Cloning and characterization of a novel bicoid-related homeobox transcription factor gene, RIEG, involved in Rieger syndrome. Nat. Genet. 1996, 14, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Shiratori, H.; Sakuma, R.; Watanabe, M.; Hashiguchi, H.; Mochida, K.; Sakai, Y.; Nishino, J.; Saijoh, Y.; Whitman, M.; Hamada, H. Two-step regulation of left-right asymmetric expression of Pitx2: Initiation by nodal signaling and maintenance by Nkx2. Mol. Cell 2001, 7, 137–149. [Google Scholar] [CrossRef]
- Yoshioka, H.; Meno, C.; Koshiba, K.; Sugihara, M.; Itoh, H.; Ishimaru, Y.; Inoue, T.; Ohuchi, H.; Semina, E.V. Pitx2, a bicoid-type homeobox gene, is involved in a lefty-signaling pathway in determination of left-right asymmetry. Cell 1998, 94, 299–305. [Google Scholar] [CrossRef]
- Semina, E.V.; Reiter, R.S.; Murray, J.C. Isolation of a new homeobox gene belonging to the Pitx/Rieg family: Expression during lens development and mapping to the aphakia region on mouse chromosome 19. Hum. Mol. Genet. 1997, 6, 2109–2116. [Google Scholar] [CrossRef] [PubMed]
- Amand, T.R.S.; Ra, J.; Zhang, Y.; Hu, Y.; Baber, S.; Qiu, M.; Chen, Y. Cloning and expression pattern of chicken Pitx2: A new component in the SHH signalling pathway controlling embryonic heart looping. Biochem. Biophys. Res. Commun. 1998, 247, 100–105. [Google Scholar] [CrossRef]
- Lin, C.R.; Kioussi, C.; O’Connell, S.; Briata, P.; Szeto, D.; Liu, F.; Izpisúa-Belmonte, J.C.; Resenfield, M.G. Pitx2 regulates lung asymmetry, cardiac positioning and pituitary and tooth morphogenesis. Nature 1999, 401, 279–282. [Google Scholar] [PubMed]
- Cunningham, E.T., Jr.; Eliott, D.; Miller, N.R.; Maumenee, I.H.; Green, W.R. Familial Axenfeld-Rieger anormaly, atrial septal defect and sensorineural hearing loss: A possible new genetic syndrome. Arch. Ophthalmol. 1998, 1666, 78–82. [Google Scholar]
- Mammi, I.; De Giorgio, P.; Clementi, M.; Tenconi, R. Cardiovascular anomaly in Rieger syndrome: Heterogeneity or contiguity? Acta Ophthalmol. Scand. 1998, 76, 509–512. [Google Scholar] [CrossRef] [PubMed]
- Tsai, J.C.; Grajewski, A.L. Cardiac valvular disease and Axenfeld-Rieger syndrome. Am. J. Ophthalmol. 1994, 118, 255–256. [Google Scholar] [CrossRef]
- Spéder, P.; Petzoldt, A.; Suzanne, M.; Noselli, S. Strategies to establish left/right asymmetry in vertebrates and invertebrates. Curr. Opin. Genet. Dev. 2007, 17, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Bakkers, J.; Verhoeven, M.C.; Abdelilah-Seyfried, S. Shaping the zebrafish heart: From left-right axis specification to epithelial tissue morphogenesis. Dev. Biol. 2009, 330, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Namigai, E.K.; Kenny, N.J.; Shimeld, S.M. Right across the tree of life: The evolution of left-right asymmetry in the Bilateria. Genesis 2014, 52, 458–470. [Google Scholar] [CrossRef] [PubMed]
- Meno, C.; Ito, Y.; Saijoh, Y.; Matsuda, Y.; Tashiro, K.; Kuhara, S.; Hamada, H. Two closely-related left-right asymmetrically expressed genes, lefty-1 and lefty-2: Their distinct expression domains, chromosomal linkage and direct neutralizing activity in Xenopus embryos. Genes Cells 1997, 2, 513–524. [Google Scholar] [CrossRef] [PubMed]
- Meno, C.; Shimono, A.; Saijoh, Y.; Yashiro, K.; Mochida, K.; Ohishi, S.; Noji, S.; Kondoh, H.; Hamada, H. lefty-1 is required for left-right determination as a regulator of lefty-2 and nodal. Cell 1998, 94, 287–297. [Google Scholar] [CrossRef]
- Oki, S.; Kitajima, K.; Marques, S.; Belo, J.A.; Yokoyama, T.; Hamada, H.; Meno, C. Reversal of left-right asymmetry induced by aberrant Nodal signaling in the node of mouse embryos. Development 2009, 136, 3917–3925. [Google Scholar] [CrossRef] [PubMed]
- Adachi, H.; Saijoh, Y.; Mochida, K.; Ohishi, S.; Hashiguchi, H.; Hirao, A.; Hamada, H. Determination of left/right asymmetric expression of nodal by a left side-specific enhancer with sequence similarity to a lefty-2 enhancer. Genes Dev. 1999, 13, 1589–1600. [Google Scholar] [CrossRef] [PubMed]
- Soukup, V.; Yong, L.W.; Lu, T.M.; Huang, S.W.; Kozmik, Z.; Yu, J.K. The Nodal signaling pathway controls left-right asymmetric development in amphioxus. EvoDevo 2015, 6, 5. [Google Scholar] [CrossRef] [PubMed]
- Fujinaga, M.; Lowe, L.A.; Kuehn, M.R. Alpha (1)-Adrenergic stimulation perturbs the left-right asymmetric expression pattern of nodal during rat embryogenesis. Teratology 2000, 62, 317–324. [Google Scholar] [CrossRef]
- Long, S.; Ahmad, N.; Rebagliati, M. The zebrafish nodal-related gene southpaw is required for visceral and diencephalic left-right asymmetry. Development 2003, 130, 2303–2316. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Lualdi, M.; Lewandoski, M.; Kuehn, M.R. Broad mesodermal and endodermal deletion of Nodal at postgastrulation stages results solely in left/right axial defects. Dev. Dyn. 2008, 237, 3591–3601. [Google Scholar] [CrossRef] [PubMed]
- Heymer, J.; Kuehn, M.; Rüther, U. The expression pattern of nodal and lefty in the mouse mutant Ft suggests a function in the establishment of handedness. Mech. Dev. 1997, 66, 5–11. [Google Scholar] [CrossRef]
- Kawasumi, A.; Nakamura, T.; Iwai, N.; Yashiro, K.; Saijoh, Y.; Belo, J.A.; Shiratori, H.; Hamada, H. Left-right asymmetry in the level of active Nodal protein produced in the node is translated into left-right asymmetry in the lateral plate of mouse embryos. Dev. Biol. 2011, 353, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Tadjuidje, E.; Kofron, M.; Mir, A.; Wylie, C.; Heasman, J.; Cha, S.W. Nodal signalling in Xenopus: The role of Xnr5 in left/right asymmetry and heart development. Open Biol. 2016, 6, 150187. [Google Scholar] [CrossRef] [PubMed]
- Vincent, S.D.; Norris, D.P.; Le Good, J.A.; Constam, D.B.; Robertson, E.J. Asymmetric Nodal expression in the mouse is governed by the combinatorial activities of two distinct regulatory elements. Mech. Dev. 2004, 121, 1403–1415. [Google Scholar] [CrossRef] [PubMed]
- Gaio, U.; Schweickert, A.; Fischer, A.; Garratt, A.N.; Müller, T.; Ozcelik, C.; Lankes, W.; Strehle, M.; Britsch, S.; Blum, M.; et al. A role of the cryptic gene in the correct establishment of the left-right axis. Curr. Biol. 1999, 9, 1339–1342. [Google Scholar] [CrossRef]
- Hashimoto, H.; Rebagliati, M.; Ahmad, N.; Muraoka, O.; Kurokawa, T.; Hibi, M.; Suzuki, T. The Cerberus/Dan-family protein Charon is a negative regulator of Nodal signaling during left-right patterning in zebrafish. Development 2004, 131, 1741–1753. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez Esteban, C.; Capdevila, J.; Economides, A.N.; Pascual, J.; Ortiz, A.; Izpisúa Belmonte, J.C. The novel Cer-like protein Caronte mediates the establishment of embryonic left-right asymmetry. Nature 1999, 401, 243–251. [Google Scholar]
- Tsiairis, C.D.; McMahon, A.P. An Hh-dependent pathway in lateral plate mesoderm enables the generation of left/right asymmetry. Curr. Biol. 2009, 19, 1912–1917. [Google Scholar] [CrossRef] [PubMed]
- Inácio, J.M.; Marques, S.; Nakamura, T.; Shinohara, K.; Meno, C.; Hamada, H.; Belo, J.A. The dynamic right-to-left translocation of Cerl2 is involved in the regulation and termination of Nodal activity in the mouse node. PLoS ONE 2013, 8, e60406. [Google Scholar] [CrossRef] [PubMed]
- Larkins, C.E.; Long, A.B.; Caspary, T. Defective Nodal and Cerl2 expression in the Arl13b(hnn) mutant node underlie its heterotaxia. Dev. Biol. 2012, 367, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Marques, S.; Borges, A.C.; Silva, A.C.; Freitas, S.; Cordenonsi, M.; Belo, J.A. The activity of the Nodal antagonist Cerl-2 in the mouse node is required for correct L/R body axis. Genes Dev. 2004, 18, 2342–2347. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Saito, D.; Kawasumi, A.; Shinohara, K.; Asai, Y.; Takaoka, K.; Dong, F.; Takamatsu, A.; Belo, J.A.; Mochizuki, A.; et al. Fluid flow and interlinked feedback loops establish left-right asymmetric decay of Cerl2 mRNA. Nat. Commun. 2012, 3, 1322. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.; Isaac, A.; Cooke, J. Nodal signalling and the roles of the transcription factors SnR and Pitx2 in vertebrate left-right asymmetry. Curr. Biol. 1999, 9, 609–612. [Google Scholar] [CrossRef]
- Kitajima, K.; Oki, S.; Ohkawa, Y.; Sumi, T.; Meno, C. Wnt signaling regulates left-right axis formation in the node of mouse embryos. Dev. Biol. 2013, 380, 222–232. [Google Scholar] [CrossRef] [PubMed]
- Supp, D.M.; Witte, D.P.; Potter, S.S.; Brueckner, M. Mutation of an axonemal dynein affects left-right asymmetry in inversus viscerum mice. Nature 1997, 389, 963–966. [Google Scholar] [CrossRef] [PubMed]
- Buceta, J.; Ibañes, M.; Rasskin-Gutman, D.; Okada, Y.; Hirokawa, N.; Izpisúa-Belmonte, J.C. Nodal cilia dynamics and the specification of the left/right axis in early vertebrate embryo development. Biophys. J. 2005, 89, 2199–2209. [Google Scholar] [CrossRef] [PubMed]
- Hjeij, R.; Lindstrand, A.; Francis, R.; Zariwala, M.A.; Liu, X.; Li, Y.; Damerla, R.; Dougherty, G.W.; Abouhamed, M.; Olbrich, H.; et al. ARMC4 mutations cause primary ciliary dyskinesia with randomization of left/right body asymmetry. Am. J. Hum. Genet. 2013, 93, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Burnicka-Turek, O.; Steimle, J.D.; Huang, W.; Felker, L.; Kamp, A.; Kweon, J.; Peterson, M.; Reeves, R.H.; Maslen, C.L.; Gruber, P.J.; et al. Cilia gene mutations cause atrioventricular septal defects by multiple mechanisms. Hum. Mol. Genet. 2016, 25, 3011–3028. [Google Scholar] [CrossRef] [PubMed]
- Hadjantonakis, A.K.; Pisano, E.; Papaioannou, V.E. Tbx6 regulates left/right patterning in mouse embryos through effects on nodal cilia and perinodal signaling. PLoS ONE 2008, 3, e2511. [Google Scholar] [CrossRef] [PubMed]
- Gros, J.; Feistel, K.; Viebahn, C.; Blum, M.; Tabin, C.J. Cell movements at Hensen’s node establish left/right asymmetric gene expression in the chick. Science 2009, 324, 941–944. [Google Scholar] [CrossRef] [PubMed]
- Bisgrove, B.W.; Essner, J.J.; Yost, H.J. Multiple pathways in the midline regulate concordant brain, heart and gut left-right asymmetry. Development 2000, 127, 3567–3579. [Google Scholar] [PubMed]
- Bisgrove, B.W.; Yost, H.J. Classification of left-right patterning defects in zebrafish, mice, and humans. Am. J. Med. Genet. 2001, 101, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Branford, W.W.; Essner, J.J.; Yost, H.J. Regulation of gut and heart left-right asymmetry by context-dependent interactions between xenopus lefty and BMP4 signaling. Dev. Biol. 2000, 223, 291–306. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Liu, W.; Lu, M.F.; Brown, N.A.; Martin, J.F. Regulation of left-right asymmetry by thresholds of Pitx2c activity. Development 2001, 128, 2039–2048. [Google Scholar] [PubMed]
- Dagle, J.M.; Sabel, J.L.; Littig, J.L.; Sutherland, L.B.; Kolker, S.J.; Weeks, D.L. Pitx2c attenuation results in cardiac defects and abnormalities of intestinal orientation in developing Xenopus laevis. Dev. Biol. 2003, 262, 268–281. [Google Scholar] [CrossRef]
- Davis, N.M.; Kurpios, N.A.; Sun, X.; Gros, J.; Martin, J.F.; Tabin, C.J. The chirality of gut rotation derives from left-right asymmetric changes in the architecture of the dorsal mesentery. Dev. Cell 2008, 15, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Essner, J.J.; Branford, W.W.; Zhang, J.; Yost, H.J. Mesendoderm and left-right brain, heart and gut development are differentially regulated by pitx2 isoforms. Development 2000, 127, 1081–1093. [Google Scholar] [PubMed]
- Guioli, S.; Lovell-Badge, R. PITX2 controls asymmetric gonadal development in both sexes of the chick and can rescue the degeneration of the right ovary. Development 2007, 134, 4199–4208. [Google Scholar] [CrossRef] [PubMed]
- Nowotschin, S.; Liao, J.; Gage, P.J.; Epstein, J.A.; Campione, M.; Morrow, B.E. Tbx1 affects asymmetric cardiac morphogenesis by regulating Pitx2 in the secondary heart field. Development 2006, 133, 1565–1573. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Selever, J.; Lu, M.F.; Martin, J.F. Genetic dissection of Pitx2 in craniofacial development uncovers new functions in branchial arch morphogenesis, late aspects of tooth morphogenesis and cell migration. Development 2003, 130, 6375–6385. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Amand, T.R.S.; Wang, S.; Li, G.; Zhang, Y.; Hu, Y.; Nguyen, L.; Qiu, M.; Chen, Y. Differential expression and functional analysis of Pitx2 isoforms in regulation of heart looping in the chick. Development 2001, 128, 1005–1101. [Google Scholar] [PubMed]
- Chinchilla, A.; Daimi, H.; Lozano-Velasco, E.; Domínguez, J.N.; Caballero, R.; Delpón, E.; Tamargo, J.; Cinca, J.; Hove-Madsen, L.; Aránega, A.E.; et al. Pitx2 insufficiency leads to electrical and structural remodelling linked to arrhythmogenesis. Circ. Cardiovasc. Genet. 2011, 4, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Klysik, E.; Sood, S.; Johnson, R.L.; Wehrens, X.H.; Martin, J.F. Pitx2 prevents susceptibility to atrial arrhythmias by inhibiting left-sided pacemarker specification. Proc. Natl. Acad. Sci. USA 2010, 107, 9753–9758. [Google Scholar] [CrossRef] [PubMed]
- Kirchhof, P.; Kahr, P.C.; Kaese, S.; Piccini, I.; Vokshi, I.; Scheld, H.H.; Rotering, H.; Fortmueller, L.; Laakmann, S.; Verheule, S.; et al. PITX2c is expressed in the adult left atrium, and reducing Pitx2c expression promotes atrial fibrillation inducibility and complex changes in gene expression. Circ. Cardiovasc. Genet. 2011, 4, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Boorman, C.J.; Shimeld, S.M. The evolution of left-right asymmetry in chordates. Bioessays 2002, 24, 1004–1011. [Google Scholar] [CrossRef] [PubMed]
- Schweickert, A.; Campione, M.; Steinbeisser, H.; Blum, M. Pitx2 isoforms: Involvement of Pitx2c but not Pitx2a or Pitx2b in vertebrate left-right asymmetry. Mech. Dev. 2000, 90, 41–51. [Google Scholar] [CrossRef]
- Linask, K.K.; Yu, X.; Chen, Y.; Han, M.D. Directionality of heart looping: Effects of Pitx2c misexpression on flectin asymmetry and midline structures. Dev. Biol. 2002, 246, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Blue, G.M.; Kirk, E.P.; Giannoulatou, E.; Sholler, G.F.; Dunwoodie, S.L.; Harvey, R.P.; Winlaw, D.S. Advances in the Genetics of Congenital Heart Disease: A Clinician’s Guide. J. Am. Coll. Cardiol. 2017, 69, 859–870. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, S.; Brueckner, M. Genetics and Genomics of Congenital Heart Disease. Circ. Res. 2017, 120, 923–940. [Google Scholar] [CrossRef] [PubMed]
- Moorman, A.; Webb, S.; Brown, N.A.; Lamers, W.; Anderson, R.H. Development of the heart: (1) Formation of the cardiac chambers and arterial trunks. Heart 2003, 89, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Furtado, M.B.; Biben, C.; Shiratori, H.; Hamada, H.; Harvey, R.P. Characterization of Pitx2c expression in the mouse heart using a reporter transgene. Dev. Dyn. 2011, 240, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Mommersteeg, M.T.; Brown, N.A.; Prall, O.W.; de Gier-de Vries, C.; Harvey, R.P.; Moorman, A.F.; Christoffels, V.M. Pitx2c and Nkx2–5 are required for the formation and identity of the pulmonary myocardium. Circ. Res. 2007, 101, 902–909. [Google Scholar] [CrossRef] [PubMed]
- Mommersteeg, M.T.; Hoogaars, W.M.; Prall, O.W.; de Gier-de Vries, C.; Wiese, C.; Clout, D.E.; Papaioannou, V.E.; Brown, N.A.; Harvey, R.P.; Moorman, A.F.; et al. Molecular pathway for the localized formation of the sinoatrial node. Circ. Res. 2007, 100, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Liu, W.; Palie, J.; Lu, M.F.; Brown, N.A.; Martin, J.F. Pitx2c patterns anterior myocardium and aortic arch vessels and is required for local cell movement into atrioventricular cushions. Development 2002, 129, 5081–5091. [Google Scholar] [PubMed]
- Ai, D.; Liu, W.; Ma, L.; Dong, F.; Lu, M.F.; Wang, D.; Verzi, M.P.; Cai, C.; Gage, P.J.; Evans, S.; et al. Pitx2 regulates cardiac left-right asymmetry by patterning second cardiac lineage-derived myocardium. Dev. Biol. 2006, 296, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Tessari, A.; Pietrobon, M.; Notte, M.; Cifelli, G.; Gage, P.J.; Schneider, M.D.; Lembi, G.; Campione, M. Myocardial Pitx2 Differentially Regulates the Left Atrial Identity and Ventricular Asymmetric Remodeling Programs. Circ. Res. 2008, 102, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Zhang, M.; Li, L.; Bai, Y.; Zhou, Y.; Moon, A.M.; Kaminski, H.J.; Martin, J.F. Pitx2, an atrial fibrillation predisposition gene, directly regulates ion transport and intercalated disc genes. Circ. Cardiovasc. Genet. 2014, 7, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Ammirabile, G.; Tessari, A.; Pignataro, V.; Szumska, D.; Sardo, F.S.; Benes, J., Jr.; Balistreri, M.; Bhattacharya, S.; Sedmera, D.; Campione, M. Pitx2 confers left morphological, molecular, and functional identity to the sinus venosus myocardium. Cardiovasc. Res. 2012, 93, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.M.; Probst, F.J.; Fox, S.E.; Schimmenti, L.A.; Semina, E.V.; Hefner, M.A.; Belmont, J.W.; Camper, S.A. Exclusion of PITX2 mutations as a major cause of CHARGE association. Am. J. Med. Genet. 2002, 111, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Muncke, N.; Niesler, B.; Roeth, R.; Schön, K.; Rüdiger, H.J.; Goldmuntz, E.; Goodship, J.; Rappold, G. Mutational analysis of the PITX2 coding region revealed no common cause for transposition of the great arteries (dTGA). BMC Med. Genet. 2005, 6, 20. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, S.; Choi, M.; Wakimoto, H.; Ma, L.; Jiang, J.; Overton, J.D.; Romano-Adesman, A.; Bjornson, R.D.; Breitbart, R.E.; Brown, K.K.; et al. De novo mutations in histone-modifying genes in congenital heart disease. Nature 2013, 498, 220–223. [Google Scholar] [CrossRef] [PubMed]
- Yuan, F.; Zhao, L.; Wang, J.; Zhang, W.; Li, X.; Qiu, X.B.; Li, R.G.; Xu, Y.J.; Xu, L.; Qu, X.K.; et al. PITX2c loss-of-function mutations responsible for congenital atrial septal defects. Int. J. Med. Sci. 2013, 10, 1422–1429. [Google Scholar] [CrossRef] [PubMed]
- Wei, D.; Gong, X.H.; Qiu, G.; Wang, J.; Yang, Y.Q. Novel PITX2c loss-of-function mutations associated with complex congenital heart disease. Int. J. Mol. Med. 2014, 33, 1201–1208. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xin, Y.F.; Xu, W.J.; Liu, Z.M.; Qiu, X.B.; Qu, X.K.; Xu, L.; Li, X.; Yang, Y.Q. Prevalence and spectrum of PITX2c mutations associated with congenital heart disease. DNA Cell Biol. 2013, 32, 708–716. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.M.; Wang, J.; Qiu, X.B.; Yuan, F.; Xu, Y.J.; Li, R.G.; Qu, X.K.; Huang, R.T.; Xue, S.; Yang, Y.Q. PITX2 loss-of-function mutation contributes to tetralogy of Fallot. Gene 2016, 577, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Pan, H.; Guan, L.; Su, D.; Ma, X. CITED2 mutation links congenital heart defects to dysregulation of the cardiac gene VEGF and PITX2C expression. Biochem. Biophys. Res. Commun. 2012, 423, 895–899. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.M.; Peng, L.Y.; Li, L.; Liu, X.Y.; Wang, J.; Zhang, X.L.; Yuan, F.; Li, R.G.; Qiu, X.B.; Yang, Y.Q. PITX2 Loss-of-Function Mutation Contributes to Congenital Endocardial Cushion Defect and Axenfeld-Rieger Syndrome. PLoS ONE 2015, 10, e0124409. [Google Scholar] [CrossRef] [PubMed]
- Hirayama-Yamada, K.; Kamisago, M.; Akimoto, K.; Aotsuka, H.; Nakamura, Y.; Tomita, H.; Furutani, M.; Imamura, S.; Takao, A.; Nakazawa, M.; et al. Phenotypes with GATA4 or NKX2.5 mutations in familial atrial septal defect. Am. J. Med. Genet. Part A 2005, 135, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Kirk, E.P.; Hyun, C.; Thomson, P.C.; Lai, D.; Castro, M.L.; Biben, C.; Buckley, M.F.; Martin, I.C.; Moran, C.; Harvey, R.P. Quantitative trait loci modifying cardiac atrial septal morphology and risk of patent foramen ovale in the mouse. Circ. Res. 2006, 98, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Posch, M.G.; Perrot, A.; Berger, F.; Ozcelik, C. Molecular genetics of congenital atrial septal defects. Clin. Res. Cardiol. 2010, 99, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Gebbia, M.; Ferrero, G.B.; Pilia, G.; Bassi, M.T.; Aylsworth, A.; Penman-Splitt, M.; Bird, L.M.; Bamforth, J.S.; Burn, J.; Schlessinger, D.; et al. X-linked situs abnormalities result from mutations in ZIC3. Nat. Genet. 1997, 17, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, E.J.; Levy, D.; Vaziri, S.M.; D’Agostino, R.B.; Belanger, A.J.; Wolf, P.A. Independent risk factors for atrial fibrillation in a population-based cohort: The Framingham Heart Study. JAMA 1994, 271, 840–844. [Google Scholar] [CrossRef] [PubMed]
- Go, A.S.; Hylek, E.M.; Philips, K.A.; Chang, Y.; Henault, L.E.; Selby, J.V. Prevalence of diagnosed atrial fibrillation in adults: National implications for rhythm management and stroke prevention: The AnTicoagulation and Risk Factors in Atrial Fibrillation (ATRIA) Study. JAMA 2001, 285, 2370–2375. [Google Scholar] [CrossRef] [PubMed]
- Krahn, A.D.; Manfreda, J.; Tate, R.B.; Mathewson, F.A.; Cuddy, T.E. The natural history of atrial fibrillation: Incidence, risk factors and prognosis in the Manitoba Follow-Up Study. Am. J. Med. 1995, 98, 476–484. [Google Scholar] [CrossRef]
- Gudbjartsson, D.F.; Arnar, D.O.; Helgadottir, A.; Gretarsdottir, S.; Holm, H.; Sigurdsson, A.; Jonasdottir, A.; Baker, A.; Thorleifsson, G.; Kristjansson, K.; et al. Var-iants conferring risk of atrial fibrillation on chromosome 4q25. Nature 2007, 448, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Gudbjartsson, D.F.; Holm, H.; Gretarsdottir, S.; Thorleifsson, G.; Walters, G.B.; Thorgeirsson, G.; Gulcher, J.; Mathiesen, E.B.; Njølstad, I.; Nyrnes, A.; et al. A sequence variant in ZFHX3 on 16q22 associates with atrial fibrillation and ischemic stroke. Nat. Genet. 2009, 41, 876–878. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, E.J.; Rice, K.M.; Arking, D.E.; Pfeufer, A.; van Noord, C.; Smith, A.V.; Schnabel, R.B.; Bis, J.C.; Boerwinkle, E.; Sinner, M.F.; et al. Variants in ZFHX3 are associated with atrial fibrilla- tion in individuals of European ancestry. Nat. Genet. 2009, 41, 879–881. [Google Scholar] [CrossRef] [PubMed]
- Ellinor, P.T.; Lunetta, K.L.; Glazer, N.L.; Pfeufer, A.; Alonso, A.; Chung, M.K.; Sinner, M.F.; de Bakker, P.I.; Mueller, M.; Lubitz, S.A.; et al. Common variants in KCNN3 are associated with lone atrial fibrillation. Nat. Genet. 2010, 42, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Schnabel, R.B.; Kerr, K.F.; Lubitz, S.A.; Alkylbekova, E.L.; Marcus, G.M.; Sinner, M.F.; Magnani, J.W.; Wolf, P.A.; Deo, R.; Lloyd-Jones, D.M.; et al. Large-scale candidate gene analysis in whites and African Americans identifies IL6R polymorphism in relation to atrial fibrillation: The National Heart, Lung, and Blood Institute’s Candidate Gene Association Resource (CARe) project. Circ. Cardiovasc. Genet. 2011, 4, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Kääb, S.; Darbar, D.; van Noord, C.; Dupuis, J.; Pfeufer, A.; Newton-Cheh, C.; Schnabel, R.; Makino, S.; Sinner, M.F.; Kannankeril, P.J.; et al. Large scale replication and meta-analysis of variants on chromosome 4q25 associated with atrial fibrillation. Eur. Heart J. 2009, 30, 813–819. [Google Scholar] [CrossRef] [PubMed]
- Lubitz, S.A.; Lunetta, K.L.; Lin, H.; Arking, D.E.; Trompet, S.; Li, G.; Krijthe, B.P.; Chasman, D.I.; Barnard, J.; Kleber, M.E.; et al. Novel genetic markers associate with atrial fibrillation risk in Europeans and Japanese. J. Am. Coll. Cardiol. 2014, 63, 1200–1210. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sun, L.; Zhang, Y.; Liang, H.; Li, X.; Cai, R.; Wang, L.; Du, W.; Zhang, R.; Li, J.; et al. Overexpression of microRNA-1 causes atrioventricular block in rodents. Int. J. Biol. Sci. 2013, 9, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Mahida, S.; Ellinor, P.T. New advances in the genetic basis of atrial fibrillation. J. Cardiovasc. Electrophysiol. 2012, 23, 1400–1406. [Google Scholar] [CrossRef] [PubMed]
- Mahida, S.; Mills, R.W.; Tucker, N.R.; Simonson, B.; Macri, V.; Lemoine, M.D.; Das, S.; Milan, D.J.; Ellinor, P.T. Overexpression of KCNN3 results in sudden cardiac death. Cardiovasc. Res. 2014, 101, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Velasco, E.; Hernández-Torres, F.; Daimi, H.; Serra, S.A.; Herraiz, A.; Hove-Madsen, L.; Aránega, A.; Franco, D. Pitx2 impairs calcium handling in a dose-dependent manner by modulating Wnt signalling. Cardiovasc. Res. 2016, 109, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, C.; Yao, Y.; Zuo, X.; Chen, S.; Xu, C.; Zhang, H.; Lu, Q.; Chang, L.; Wang, F.; et al. Molecular Basis of Gene-Gene Interaction: Cyclic Cross-Regulation of Gene Expression and Post-GWAS Gene-Gene Interaction Involved in Atrial Fibrillation. PLoS Genet. 2015, 11, e1005393. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.M.; Yang, F.; Mahida, S.; Zhao, L.; Chen, X.; Zhang, M.L.; Sun, Z.; Yao, Y.; Zhang, Y.X.; Zheng, G.Y.; et al. TBX5 mutations contribute to early-onset atrial fibrillation in Chinese and Caucasians. Cardiovasc. Res. 2016, 109, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Nadadur, R.D.; Broman, M.T.; Boukens, B.; Mazurek, S.R.; Yang, X.; van den Boogaard, M.; Bekeny, J.; Gadek, M.; Ward, T.; Zhang, M.; et al. Pitx2 modulates a Tbx5-dependent gene regulatory network to maintain atrial rhythm. Sci. Transl. Med. 2016, 8, 354ra115. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Bai, Y.; Li, N.; Ye, W.; Zhang, M.; Greene, S.B.; Tao, Y.; Chen, Y.; Wehrens, X.H.; Martin, J.F. Pitx2-microRNA pathway that delimits sinoatrial node development and inhibits predisposition to atrial fibrillation. Proc. Natl. Acad. Sci. USA 2014, 111, 9181–9186. [Google Scholar] [CrossRef] [PubMed]
- Xia, M.; Jin, Q.; Bendahhou, S.; He, Y.; Larroque, M.M.; Chen, Y.; Zhou, Q.; Yang, Y.; Liu, Y.; Liu, B.; et al. A Kir2.1 gain-of-function mutation underlies familial atrial fibrillation. Biochem. Biophys. Res. Commun. 2005, 332, 1012–1019. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Xu, S.J.; Bendahhou, S.; Wang, X.L.; Wang, Y.; Xu, W.Y.; Jin, H.W.; Sun, H.; Su, X.Y.; Zhuang, Q.N.; et al. KCNQ1 gain-of-function mutation in familial atrial fibrillation. Science 2003, 299, 251–254. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, L.A.; Alonso, M.E.; Badía-Careaga, C.; Rollán, I.; Arias, C.; Fernández-Miñán, A.; López-Jiménez, E.; Aránega, A.; Gómez-Skarmeta, J.L.; Franco, D.; et al. Long-range regulatory interactions at the 4q25 atrial fibrillation risk locus involve PITX2c and ENPEP. BMC Biol. 2015, 13, 26. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Tucker, N.R.; Weng, L.C.; Clauss, S.; Lubitz, S.A.; Ellinor, P.T. A Functional Variant Associated with Atrial Fibrillation Regulates PITX2c Expression through TFAP2a. Am. J. Hum. Genet. 2016, 99, 1281–1291. [Google Scholar] [CrossRef] [PubMed]
- Torrado, M.; Franco, D.; Lozano-Velasco, E.; Hernández-Torres, F.; Calviño, R.; Aldama, G.; Centeno, A.; Castro-Beiras, A.; Mikhailov, A. A MicroRNA-Transcription Factor Blueprint for Early Atrial Arrhythmogenic Remodeling. BioMed Res. Int. 2015, 2015, 263151. [Google Scholar] [CrossRef] [PubMed]
- Syeda, F.; Holmes, A.P.; Yu, T.Y.; Tull, S.; Kuhlmann, S.M.; Pavlovic, D.; Betney, D.; Riley, G.; Kucera, J.; Jousset, F.; et al. Pitx2 modulates atrial membrane potential and reduced PITX2 potentiates the anti-arrhythmic effects of socium-channel blockers. J. Am. Coll. Cardiol. 2016, 68, 1881–1894. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Yang, Y.-Q.; Gong, X.Q.; Wang, X.-H.; Li, R.-G.; Tan, H.-W.; Liu, X.; Fang, W.-Y.; Bai, D. Novel germline GJA5/connexin40 mutations associated with the lone atrial fibrillation impair gap junctional intercellular communication. Hum. Mutat. 2013, 34, 603–609. [Google Scholar] [PubMed]
- Pérez-Hernández, M.; Matamoros, M.; Barana, A.; Amorós, I.; Gómez, R.; Núñez, M.; Sacristán, S.; Pinto, A.; Fernández-Avilés, F.; Tamargo, J.; et al. Pitx2c increases in atrial myocites from chronic atrial fibrillation patients enhancing IKs and devreasin ICa, L. Cardiovasc. Res. 2016, 109, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Girmatsion, Z.; Biliczki, P.; Bonauer, A.; Wimmer-Greinecker, G.; Scherer, M.; Moritz, A.; Bukowska, A.; Goette, A.; Nattel, S.; Hohnloser, S.H.; et al. Changes in microRNA-1 expression and IK1 up-regulation in human atrial fibrillation. Heart Rhythm 2009, 6, 1802–1809. [Google Scholar] [CrossRef] [PubMed]
- Cooley, N.; Cowley, M.J.; Lin, R.C.; Marasco, S.; Wong, C.; Kaye, D.M.; Dart, A.M.; Woodcock, E.A. Influence of atrial fibrillation on microRNA expression profiles in left and right atria from patients with valvular heart disease. Physiol. Genom. 2012, 44, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Scridon, A.; Fouilloux-Meugnier, E.; Loizon, E.; Rome, S.; Julien, C.; Barrès, C.; Chevalier, P. Long-standing arterial hypertension is associated with Pitx2 down-regulation in a rat model of spontaneous atrial tachyarrhythmias. Europace 2015, 17, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Fakhro, K.A.; Choi, M.; Ware, S.M.; Belmont, J.W.; Towbin, J.A.; Lifton, R.P.; Khokha, M.K.; Brueckner, M. Rare copy number variations in congenital heart disease patients identify unique genes in left-right patterning. Proc. Natl. Acad. Sci. USA 2011, 108, 2915–2920. [Google Scholar] [CrossRef] [PubMed]
- Franco, D.; Chinchilla, A.; Daimi, H.; Dominguez, J.N.; Aránega, A. Modulation of conductive elements by Pitx2 and their impact on atrial arrhythmogenesis. Cardiovasc. Res. 2011, 91, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Franco, D.; Chinchilla, A.; Aránega, A.E. Transgenic insights linking pitx2 and atrial arrhythmias. Front. Physiol. 2012, 3, 206. [Google Scholar] [CrossRef] [PubMed]
- Franco, D.; Christoffels, V.M.; Campione, M. Homeobox transcription factor Pitx2: The rise of an asymmetry gene in cardiogenesis and arrhythmogenesis. Trends Cardiovasc. Med. 2014, 24, 23–31. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).