

Exercise Intervention in Autonomic Function, Immunity, and Cardiovascular Health: A Precision Medicine Approach


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
3. Autonomic–Immune Interactions: Theoretical Mechanisms and Exercise Modulation
3.1. Regulatory Pathways of the ANS in Immune Modulation
3.2. Feedback Regulation of the ANS by the Immune System
3.3. The Bidirectional Regulatory Effects of Exercise Intervention
4. Precision-Medicine-Driven Personalized Exercise Interventions
4.1. Individual Variability Analysis Based on Multi-Omics Approaches
4.2. Optimization of Exercise Prescriptions Guided by Dynamic Biomarkers
4.3. Clinical Translation: Case Studies and Challenges
5. Precision Exercise and Multimodal Data Integration in Cardiovascular–Immune Regulation
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Manolis, A.A.; Manolis, T.A.; Apostolopoulos, E.J.; Apostolaki, N.E.; Melita, H.; Manolis, A.S.L. The role of the autonomic nervous system in cardiac arrhythmias: The neuro-cardiac axis, more foe than friend? Trends Cardiovas. Med. 2021, 31, 290–302. [Google Scholar] [CrossRef]
- Lipsett, M.J.; Tsai, F.C.; Roger, L.; Woo, M.; Ostro, B.D. Coarse particles and heart rate variability among older adults with coronary artery disease in the Coachella Valley, California. Environ. Health Perspect. 2006, 114, 1215–1220. [Google Scholar] [CrossRef] [PubMed]
- Kaze, A.D.; Yuyun, M.F.; Erqou, S.; Fonarow, G.C.; Echouffo-Tcheugui, J.B. Cardiac autonomic neuropathy and risk of incident heart failure among adults with type 2 diabetes. Eur. J. Heart Fail. 2022, 24, 634–641. [Google Scholar] [CrossRef] [PubMed]
- Grässler, B.; Thielmann, B.; Böckelmann, I.; Hökelmann, A. Effects of different exercise interventions on cardiac autonomic control and secondary health factors in middle-aged adults: A systematic review. J. Cardiovasc. Dev. Dis. 2021, 8, 94. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, G.; Zhang, D.; Zhang, K.; Cao, C. Physiological mechanisms driving microcirculatory enhancement: The impact of physical activity. Rev. Cardiovasc. Med. 2025, 26, 25302. [Google Scholar] [CrossRef]
- Margonis, K.; Fatouros, I.G.; Jamurtas, A.Z.; Nikolaidis, M.G.; Douroudos, I.; Chatzinikolaou, A.; Mitrakou, A.; Mastorakos, G.; Papassotiriou, I.; Taxildaris, K.; et al. Oxidative stress biomarkers responses to physical overtraining: Implications for diagnosis. Free Radic. Biol. Med. 2007, 43, 901–910. [Google Scholar] [CrossRef]
- Solis-Urra, P.; Fernandez-Gamez, B.; Liu-Ambrose, T.; Erickson, K.I.; Ortega, F.B.; Esteban-Cornejo, I. Exercise as medicine for the brain: Moving towards precise and personalised recommendations. Brit. J. Sports Med. 2024, 58, 693–695. [Google Scholar] [CrossRef]
- Yusof, H.A.; Aziz, A.R.; Muhamed, A.M.C. The influence of angiotensin I-converting enzyme (ACE) I/D gene polymorphism on cardiovascular and muscular adaptations following 8 weeks of isometric handgrip training (IHG) in untrained normotensive males. Biol. Sport 2019, 36, 81–94. [Google Scholar] [CrossRef]
- Ross, R.; Goodpaster, B.H.; Koch, L.G.; Sarzynski, M.A.; Kohrt, W.M.; Johannsen, N.M.; Skinner, J.S.; Castro, A.; Irving, B.A.; Noland, R.C.; et al. Precision exercise medicine: Understanding exercise response variability. Brit. J. Sports Med. 2019, 53, 1141–1153. [Google Scholar] [CrossRef]
- Bıçakçı, B.; Cięszczyk, P.; Humińska-Lisowska, K. Genetic determinants of endurance: A narrative review on elite athlete status and performance. Int. J. Mol. Sci. 2024, 25, 13041. [Google Scholar] [CrossRef]
- Eynon, N.; Meckel, Y.; Sagiv, M.; Yamin, C.; Amir, R.; Sagiv, M.; Goldhammer, E.; Duarte, J.A.; Oliveira, J. Do PPARGC1A and PPARalpha polymorphisms influence sprint or endurance phenotypes? Scand. J. Med. Sci. Sports 2010, 20, e145–e150. [Google Scholar] [CrossRef] [PubMed]
- Bachem, A.; Makhlouf, C.; Binger, K.J.; de Souza, D.P.; Tull, D.; Hochheiser, K.; Whitney, P.G.; Fernandez-Ruiz, D.; Dähling, S.; Kastenmüller, W.; et al. Microbiota-derived short-chain fatty acids promote the memory potential of antigen-activated CD8+ T Cells. Immunity 2019, 51, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Maniaci, A.; Lavalle, S.; Parisi, F.M.; Barbanti, M.; Cocuzza, S.; Iannella, G.; Magliulo, G.; Pace, A.; Lentini, M.; Masiello, E.; et al. Impact of obstructive sleep apnea and sympathetic nervous system on cardiac health: A comprehensive review. J. Cardiovasc. Dev. Dis. 2024, 11, 204. [Google Scholar] [CrossRef]
- Pavlov, V.A.; Chavan, S.S.; Tracey, K.J. Molecular and functional neuroscience in immunity. Annu. Rev. Immunol. 2018, 36, 783–812. [Google Scholar] [CrossRef]
- Jacobson, A.; Yang, D.; Vella, M.; Chiu, I.M. The intestinal neuro-immune axis: Crosstalk between neurons, immune cells, and microbes. Mucosal. Immunol. 2021, 14, 555–565. [Google Scholar] [CrossRef]
- Elenkov, I.J.; Wilder, R.L.; Chrousos, G.P.; Vizi, E.S. The sympathetic nerve—An integrative interface between two supersystems: The brain and the immune system. Pharmacol. Rev. 2000, 52, 595–638. [Google Scholar] [CrossRef]
- Sleightholm, R.L.; Neilsen, B.K.; Li, J.; Steele, M.M.; Singh, R.K.; Hollingsworth, M.A.; Oupicky, D. Emerging roles of the CXCL12/CXCR4 axis in pancreatic cancer progression and therapy. Pharmacol. Therapeut. 2017, 179, 158–170. [Google Scholar] [CrossRef]
- Dutta, P.; Courties, G.; Wei, Y.; Leuschner, F.; Gorbatov, R.; Robbins, C.S.; Iwamoto, Y.; Thompson, B.; Carlson, A.L.; Heidt, T.; et al. Myocardial infarction accelerates atherosclerosis. Nature 2012, 487, 325–329. [Google Scholar] [CrossRef]
- Rice, P.A.; Boehm, G.W.; Moynihan, J.A.; Bellinger, D.L.; Stevens, S.Y. Chemical sympathectomy alters numbers of splenic and peritoneal leukocytes. Brain Behav. Immun. 2002, 16, 62–73. [Google Scholar] [CrossRef]
- Alen, N.V. The cholinergic anti-inflammatory pathway in humans: State-of-the-art review and future directions. Neurosci. Biobehav. Rev. 2022, 136, 104622. [Google Scholar] [CrossRef]
- Zhang, B.J.; Men, X.J.; Lu, Z.Q.; Li, H.Y.; Qiu, W.; Hu, X.Q. Splenectomy protects experimental rats from cerebral damage after stroke due to anti-inflammatory effects. Chin. Med. J. 2013, 126, 2354–2360. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, S.; Inoue, T.; Nakamura, Y.; Fukaya, D.; Uni, R.; Wu, C.H.; Fujii, R.; Peerapanyasut, W.; Taguchi, A.; Kohro, T.; et al. Activation of sympathetic signaling in macrophages blocks systemic inflammation and protects against renal ischemia-reperfusion injury. J. Am. Soc. Nephrol. 2012, 32, 1599–1615. [Google Scholar] [CrossRef]
- Wang, R.; Zhang, W.; Dong, Z.; Qi, Y.; Hultström, M.; Zhou, X.; Lai, E.Y. c-Jun N-terminal Kinase mediates prostaglandin-induced sympathoexcitation in rats with chronic heart failure by reducing GAD1 and GABRA1 expression. Acta Physiol. 2017, 219, 494–509. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hu, H.; Yin, J.; Shi, Y.; Tan, J.; Zheng, L.; Wang, C.; Li, X.; Xue, M.; Liu, J.; et al. TLR4 participates in sympathetic hyperactivity Post-MI in the PVN by regulating NF-κB pathway and ROS production. Redox Biol. 2019, 24, 101186. [Google Scholar] [CrossRef]
- Bassi, G.S.; Brognara, F.; Castania, J.A.; Talbot, J.; Cunha, T.M.; Cunha, F.Q.; Ulloa, L.; Kanashiro, A.; Dias, D.P.; Salgado, H.C. Baroreflex activation in conscious rats modulates the joint inflammatory response via sympathetic function. Brain Behav. Immun. 2015, 49, 140–147. [Google Scholar] [CrossRef]
- Rosas-Ballina, M.; Ochani, M.; Parrish, W.R.; Ochani, K.; Harris, Y.T.; Huston, J.M.; Chavan, S.; Tracey, K.J. Splenic nerve is required for cholinergic anti-inflammatory pathway control of TNF in endotoxemia. Proc. Natl. Acad. Sci. USA 2008, 105, 11008–11013. [Google Scholar] [CrossRef]
- Carnevale, D.; Lembo, G. Neuroimmune interactions in cardiovascular diseases. Cardiovasc. Res. 2021, 117, 402–410. [Google Scholar] [CrossRef]
- Obata, Y.; Pachnis, V. The effect of microbiota and the immune system on the development and organization of the enteric nervous system. Gastroenterology 2016, 151, 836–844. [Google Scholar] [CrossRef]
- Cutolo, M.; Straub, R.H. Circadian rhythms in arthritis: Hormonal effects on the immune/inflammatory reaction. Autoimmun. Rev. 2008, 7, 223–228. [Google Scholar] [CrossRef]
- Dantzer, R.; O’Connor, J.C.; Freund, G.G.; Johnson, R.W.; Kelley, K.W. From inflammation to sickness and depression: When the immune system subjugates the brain. Nat. Rev. Neurosci. 2008, 9, 46–56. [Google Scholar] [CrossRef]
- Fiedorowicz, J.G. Depression and cardiovascular disease: An update on how course of illness may influence risk. Curr. Psychiatry Rep. 2014, 16, 492. [Google Scholar] [CrossRef]
- Díaz, H.S.; Toledo, C.; Andrade, D.C.; Marcus, N.J.; Del Rio, R. Neuroinflammation in heart failure: New insights for an old disease. J. Physiol. Sci. 2020, 598, 33–59. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.M.; Gao, F.; Li, H.H. NF-κB in the paraventricular nucleus modulates neurotransmitters and contributes to sympathoexcitation in heart failure. Basic Res. Cardiol. 2011, 106, 1087–1097. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Wang, S.; Lin, W.; Gu, J.; Li, G.; Shao, Y. BMAL1 promotes valvular interstitial cells’ osteogenic differentiation through NF-κ B/AKT/MAPK pathway. J. Cardiovasc. Dev. Dis. 2023, 10, 110. [Google Scholar] [CrossRef]
- Halaris, A. Inflammation, heart disease, and depression. Curr. Psychiatry Rep. 2013, 15, 400. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, D.R.; Bekhbat, M.; Mehta, N.D.; Felger, J.C. Inflammation-related functional and structural dysconnectivity as a pathway to psychopathology. Biol. Psychiatry 2023, 93, 405–418. [Google Scholar] [CrossRef]
- Andrade, D.C.; Arce-Alvarez, A.; Toledo, C.; Díaz, H.S.; Lucero, C.; Schultz, H.D.; Marcus, N.J.; Del Rio, R. Exercise training improves cardiac autonomic control, cardiac function, and arrhythmogenesis in rats with preserved-ejection fraction heart failure. J. Appl. Physiol. 2017, 123, 567–577. [Google Scholar] [CrossRef]
- Tracey, K.J. The inflammatory reflex. Nature 2002, 420, 853–859. [Google Scholar] [CrossRef]
- Ribeiro, D.; Petrigna, L.; Pereira, F.C.; Muscella, A.; Bianco, A.; Tavares, P. The Impact of Physical Exercise on the Circulating Levels of BDNF and NT 4/5: A Review. Int. J. Mol. Sci. 2021, 22, 8814. [Google Scholar] [CrossRef]
- Sarmento, A.O.; Antunes-Correa, L.M.; Alves, M.J.N.N.; Bacurau, A.V.N.; Fonseca, K.C.B.; Pessoa, F.G.; Lobo, D.M.L.; Moreira, L.D.P.; Trombetta, I.C.; Rondon, M.U.P.B.; et al. Effect of exercise training on cardiovascular autonomic and muscular function in subclinical Chagas cardiomyopathy: A randomized controlled trial. Clin. Auton. Res. 2021, 31, 239–251. [Google Scholar] [CrossRef]
- Chen, H.; Shen, L.; Liu, Y.; Ma, X.; Long, L.; Ma, X.; Ma, L.; Chen, Z.; Lin, X.; Si, L.; et al. Strength exercise confers protection in central nervous system autoimmunity by altering the gut microbiota. Front. Immunol. 2021, 12, 628629. [Google Scholar] [CrossRef] [PubMed]
- Just, T.P.; Delorey, D.S. Exercise training and α1-adrenoreceptor-mediated sympathetic vasoconstriction in resting and contracting skeletal muscle. Physiol. Rep. 2016, 4, e12707. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, S.P.; Nyberg, M.; Gliemann, L.; Thaning, P.; Saltin, B.; Hellsten, Y. Exercise training modulates functional sympatholysis and α-adrenergic vasoconstrictor responsiveness in hypertensive and normotensive individuals. J. Physiol. London. 2014, 592, 3063–3073. [Google Scholar] [CrossRef]
- Jeong, J.; Sprick, J.D.; DaCosta, D.R. Exercise modulates sympathetic and vascular function in chronic kidney disease. JCI insight. 2023, 8, e164221. [Google Scholar] [CrossRef]
- Armstrong, L.E.; VanHeest, J.L. The unknown mechanism of the overtraining syndrome: Clues from depression and psychoneuroimmunology. Sports Med. 2002, 32, 185–209. [Google Scholar] [CrossRef]
- Budgett, R.; Hiscock, N.; Arida, R.M.; Castell, L.M. The effects of the 5-HT2C agonist m-chlorophenylpiperazine on elite athletes with unexplained underperformance syndrome (overtraining). Br. J. Sports Med. 2010, 44, 280–283. [Google Scholar] [CrossRef]
- Lehmann, M.; Foster, C.; Keul, J. Overtraining in endurance athletes: A brief review. Med. Sci. Sports Exerc. 1993, 25, 854–862. [Google Scholar] [CrossRef]
- Pichot, V.; Roche, F.; Gaspoz, J.M.; Enjolras, F.; Antoniadis, A.; Minini, P.; Costes, F.; Busso, T.; Lacour, J.R.; Barthélémy, J.C. Relation between heart rate variability and training load in middle-distance runners. Med. Sci. Sports Exerc. 2000, 32, 1729–1736. [Google Scholar] [CrossRef]
- Coqueiro, A.Y.; Rogero, M.M.; Tirapegui, J. Glutamine as an anti-fatigue amino acid in sports nutrition. Nutrients 2019, 11, 863. [Google Scholar] [CrossRef]
- Cuparencu, C.; Bulmuş-Tüccar, T.; Stanstrup, J. Towards nutrition with precision: Unlocking biomarkers as dietary assessment tools. Nat. Metab. 2024, 6, 1438–1453. [Google Scholar] [CrossRef]
- Agudelo, L.Z.; Femenía, T.; Orhan, F.; Porsmyr-Palmertz, M.; Goiny, M.; Martinez-Redondo, V.; Correia, J.C.; Izadi, M.; Bhat, M.; Schuppe-Koistinen, I.; et al. Skeletal muscle PGC-1α1 modulates kynurenine metabolism and mediates resilience to stress-induced depression. Cell 2014, 159, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Schwarcz, R.; Bruno, J.P.; Muchowski, P.J.; Wu, H.Q. Kynurenines in the mammalian brain: When physiology meets pathology. Nat. Rev. Neurosci. 2012, 13, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Verweij, N.; van de Vegte, Y.J.; van der Harst, P. Genetic study links components of the autonomous nervous system to heart-rate profile during exercise. Nat. Commun. 2018, 9, 898. [Google Scholar] [CrossRef]
- Saco-Ledo, G.; Valenzuela, P.L.; Almazán-Polo, J.; Plaza-Florido, A.; Alejo, L.B.; Bustos, A.; Río-García, A.; Gálvez, B.G.; Rubio-González, E.; Fiuza-Luces, C.; et al. Acute physical exercise and ambulatory blood pressure in resistant hypertension. J. Hypertens. 2025, 43, 445–455. [Google Scholar] [CrossRef]
- Timmons, J.A.; Knudsen, S.; Rankinen, T.; Koch, L.G.; Sarzynski, M.; Jensen, T.; Keller, P.; Scheele, C.; Vollaard, N.B.; Nielsen, S.; et al. Using molecular classification to predict gains in maximal aerobic capacity following endurance exercise training in humans. J. Appl. Physiol. 2010, 108, 1487–1496. [Google Scholar] [CrossRef]
- Gleeson, M.; Bishop, N.C.; Stensel, D.J.; Lindley, M.R.; Mastana, S.S.; Nimmo, M.A. The anti-inflammatory effects of exercise: Mechanisms and implications for the prevention and treatment of disease. Nat. Rev. Immunol. 2011, 11, 607–615. [Google Scholar] [CrossRef]
- Guntur, V.P.; Nemkov, T.; de Boer, E.; Mohning, M.P.; Baraghoshi, D.; Cendali, F.I.; San-Millán, I.; Petrache, I.; D’Alessandro, A. Signatures of mitochondrial dysfunction and impaired fatty acid metabolism in plasma of patients with post-acute sequelae of COVID-19 (PASC). Metabolites 2022, 12, 1026. [Google Scholar] [CrossRef]
- Bibi, A.; Bartekova, M.; Gandhi, S.; Greco, S.; Madè, A.; Sarkar, M.; Stopa, V.; Tastsoglou, S.; de Gonzalo-Calvo, D.; Devaux, Y.; et al. Circular RNA regulatory role in pathological cardiac remodelling. Brit. J. Pharmacol. 2025, 182, 316–339. [Google Scholar] [CrossRef]
- Zhou, L.; Aljiffry, A.; Lee, Y.J.; Matthews, J.; Seitter, B.; Soltis, I.; Huang, Y.; Maher, K.; Yeo, W.H. Soft imperceptible wearable electronics for at-home cardiovascular monitoring of infants with single ventricle heart disease. Biosens. Bioelectron. 2025, 278, 117372. [Google Scholar] [CrossRef]
- Li, G.; Zhou, X.; Deng, J.; Wang, J.; Ai, P.; Zeng, J.; Ma, X.; Liao, H. Digital Therapeutics-Based Cardio-Oncology Rehabilitation for Lung Cancer Survivors: Randomized Controlled Trial. JMIR mHealth uHealth 2025, 13, e60115. [Google Scholar] [CrossRef]
- Le, T.Q.; Bukkapatnam, S.T.; Komanduri, R. Real-time lumped parameter modeling of cardiovascular dynamics using electrocardiogram signals: Toward virtual cardiovascular instruments. IEEE Trans. Biomed. Eng. 2013, 60, 2350–2360. [Google Scholar] [CrossRef] [PubMed]
- Patel, V.N.; Pierce, B.R.; Bodapati, R.K.; Brown, D.L.; Ives, D.G.; Stein, P.K. Association of Holter-Derived Heart Rate Variability Parameters with the development of congestive heart failure in the cardiovascular health study. JACC Heart Fail. 2017, 5, 423–431. [Google Scholar] [CrossRef]
- Odeh, V.A.; Chen, Y.; Wang, W.; Ding, X. Recent advances in the wearable devices for monitoring and management of heart failure. Rev. Cardiovasc. Med. 2024, 25, 386. [Google Scholar] [CrossRef]
- Dorje, T.; Zhao, G.; Tso, K.; Wang, J.; Chen, Y.; Tsokey, L.; Tan, B.K.; Scheer, A.; Jacques, A.; Li, Z.; et al. Smartphone and social media-based cardiac rehabilitation and secondary prevention in China (SMART-CR/SP): A parallel-group, single-blind, randomised controlled trial. Lancet Digit. Health 2019, 1, e363–e374. [Google Scholar] [CrossRef] [PubMed]
- Güder, G.; Wilkesmann, J.; Scholz, N.; Leppich, R.; Düking, P.; Sperlich, B.; Rost, C.; Frantz, S.; Morbach, C.; Sahiti, F.; et al. Establishing a cardiac training group for patients with heart failure: The “HIP-in-Würzburg” study. Clin. Res. Cardiol. 2022, 111, 406–415. [Google Scholar] [CrossRef]
- Okwose, N.C.; Avery, L.; O’Brien, N.; Cassidy, S.; Charman, S.J.; Bailey, K.; Velicki, L.; Olivotto, I.; Brennan, P.; MacGowan, G.A.; et al. Acceptability, feasibility and preliminary evaluation of a novel, personalised, home-based physical activity intervention for chronic heart failure (Active-at-Home-HF): A pilot study. Sports Med. Open 2019, 5, 45. [Google Scholar] [CrossRef] [PubMed]
- Lyall, D.M.; Kormilitzin, A.; Lancaster, C.; Sousa, J.; Petermann-Rocha, F.; Buckley, C.; Harshfield, E.L.; Iveson, M.H.; Madan, C.R.; McArdle, R.; et al. Artificial intelligence for dementia-applied models and digital health. Alzheimer’s Dement. 2023, 19, 5872–5884. [Google Scholar] [CrossRef]
- Thangaraj, P.M.; Benson, S.H.; Oikonomou, E.K.; Asselbergs, F.W.; Khera, R. Cardiovascular care with digital twin technology in the era of generative artificial intelligence. Eur. Heart J. 2024, 45, 4808–4821. [Google Scholar] [CrossRef]
- Lee, J.W.; Su, Y.; Baloni, P.; Chen, D.; Pavlovitch-Bedzyk, A.J.; Yuan, D.; Duvvuri, V.R.; Ng, R.H.; Choi, J.; Xie, J.; et al. Integrated analysis of plasma and single immune cells uncovers metabolic changes in individuals with COVID-19. Nat. Biotechnol. 2022, 40, 110–120. [Google Scholar] [CrossRef]
- Schäfer, P.S.L.; Dimitrov, D.; Villablanca, E.J.; Saez-Rodriguez, J. Integrating single-cell multi-omics and prior biological knowledge for a functional characterization of the immune system. Nat. Immunol. 2024, 25, 405–417. [Google Scholar] [CrossRef]
- Ilonze, O.J.; Forman, D.E.; LeMond, L.; Myers, J.; Hummel, S.; Vest, A.R.; DeFilippis, E.M.; Habib, E.; Goodlin, S.J. Beyond guideline-directed medical therapy: Nonpharmacologic management for patients with heart failure. JACC Heart Fail. 2025, 13, 185–199. [Google Scholar] [CrossRef] [PubMed]
 : promote;
: promote;  : inhibit. Abbreviation: NF-κB: nuclear factor-kappa B; JAK-STAT3: janus kinase 2-signal transducer and activator of transcription 3; Ach: acetylcholine; β2-AR: β2-adrenergic receptors; NE: norepinephrine; α7-AchR: α7 nicotinic acetylcholine receptors; IL-1β: interleukin-1β; IL-6: interleukin-6; IL-10: interleukin-10; IL-18: interleukin-18; TNF-α: tumor necrosis factor-alpha; TGF-β: transforming growth factor-beta; cAMP: cyclic adenosine monophosphate; α1-AR: α1-adrenergic receptors; α2-AR: α2-adrenergic receptors; PI3K-AKT: phosphatidylinositol-3-kinase/protein kinase B; CD8 T cell: Cytotoxic T lymphocyte; CXCL12: Chemokine (C-X-C motif) ligand 12; CXCR4: Chemokine (C-X-C motif) receptor type 4.
: promote; : inhibit. Abbreviation: NF-κB: nuclear factor-kappa B; JAK-STAT3: janus kinase 2-signal transducer and activator of transcription 3; Ach: acetylcholine; β2-AR: β2-adrenergic receptors; NE: norepinephrine; α7-AchR: α7 nicotinic acetylcholine receptors; IL-1β: interleukin-1β; IL-6: interleukin-6; IL-10: interleukin-10; IL-18: interleukin-18; TNF-α: tumor necrosis factor-alpha; TGF-β: transforming growth factor-beta; cAMP: cyclic adenosine monophosphate; α1-AR: α1-adrenergic receptors; α2-AR: α2-adrenergic receptors; PI3K-AKT: phosphatidylinositol-3-kinase/protein kinase B; CD8 T cell: Cytotoxic T lymphocyte; CXCL12: Chemokine (C-X-C motif) ligand 12; CXCR4: Chemokine (C-X-C motif) receptor type 4.
: inhibit. Abbreviation: NF-κB: nuclear factor-kappa B; JAK-STAT3: janus kinase 2-signal transducer and activator of transcription 3; Ach: acetylcholine; β2-AR: β2-adrenergic receptors; NE: norepinephrine; α7-AchR: α7 nicotinic acetylcholine receptors; IL-1β: interleukin-1β; IL-6: interleukin-6; IL-10: interleukin-10; IL-18: interleukin-18; TNF-α: tumor necrosis factor-alpha; TGF-β: transforming growth factor-beta; cAMP: cyclic adenosine monophosphate; α1-AR: α1-adrenergic receptors; α2-AR: α2-adrenergic receptors; PI3K-AKT: phosphatidylinositol-3-kinase/protein kinase B; CD8 T cell: Cytotoxic T lymphocyte; CXCL12: Chemokine (C-X-C motif) ligand 12; CXCR4: Chemokine (C-X-C motif) receptor type 4.
: promote; : inhibit. Abbreviation: NF-κB: nuclear factor-kappa B; JAK-STAT3: janus kinase 2-signal transducer and activator of transcription 3; Ach: acetylcholine; β2-AR: β2-adrenergic receptors; NE: norepinephrine; α7-AchR: α7 nicotinic acetylcholine receptors; IL-1β: interleukin-1β; IL-6: interleukin-6; IL-10: interleukin-10; IL-18: interleukin-18; TNF-α: tumor necrosis factor-alpha; TGF-β: transforming growth factor-beta; cAMP: cyclic adenosine monophosphate; α1-AR: α1-adrenergic receptors; α2-AR: α2-adrenergic receptors; PI3K-AKT: phosphatidylinositol-3-kinase/protein kinase B; CD8 T cell: Cytotoxic T lymphocyte; CXCL12: Chemokine (C-X-C motif) ligand 12; CXCR4: Chemokine (C-X-C motif) receptor type 4.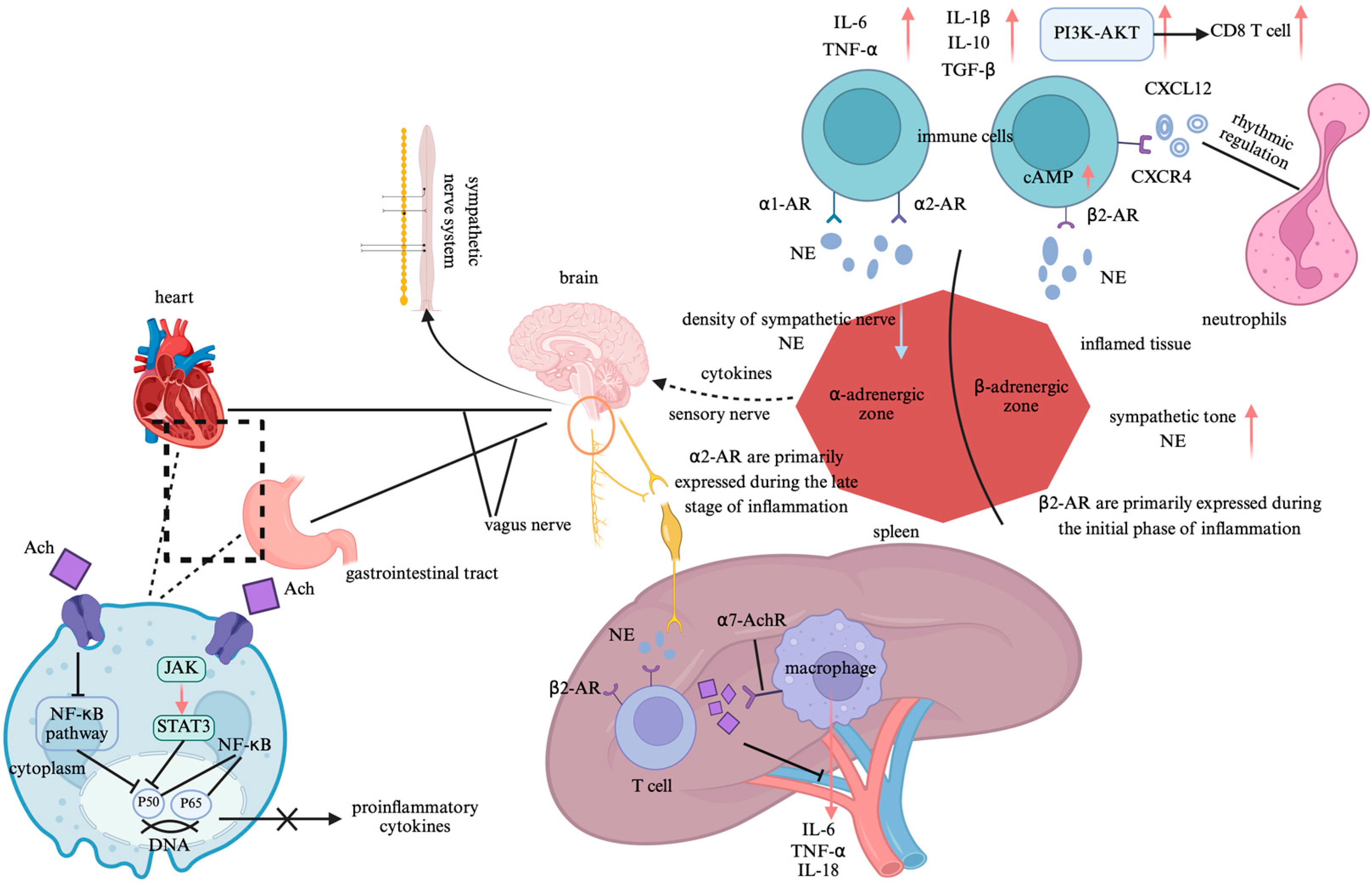
 : promote;
: promote;  : inhibit. Abbreviation: Ang II: angiotensin II; NADPH: nicotinamide adenine dinucleotide phosphate; RhoA: Ras homolog gene family member A; AT1: angiotensin type 1; CVOs: circumventricular organs; PVN: paraventricular nucleus; RVLM: rostral ventrolateral medulla; GABA: gamma-aminobutyric acid; NF-κB/AP-1: nuclear translocation of nuclear factor-kappa B/adaptor protein 1; 5-HT: 5-hydroxytryptamine; BDNF: brain-derived neurotrophic factor; ROS: reactive oxygen species; IL-6: interleukin-6; TNF-α: tumor necrosis factor-alpha; IL-1β: interleukin-1β; ACE receptor: Angiotensin-converting enzyme receptor; NE: norepinephrine; Ach: Acetylcholine; IDO: indoleamine 2,3-dioxygenase; α-7AchR: alpha-7 acetylcholine receptor.
: promote; : inhibit. Abbreviation: Ang II: angiotensin II; NADPH: nicotinamide adenine dinucleotide phosphate; RhoA: Ras homolog gene family member A; AT1: angiotensin type 1; CVOs: circumventricular organs; PVN: paraventricular nucleus; RVLM: rostral ventrolateral medulla; GABA: gamma-aminobutyric acid; NF-κB/AP-1: nuclear translocation of nuclear factor-kappa B/adaptor protein 1; 5-HT: 5-hydroxytryptamine; BDNF: brain-derived neurotrophic factor; ROS: reactive oxygen species; IL-6: interleukin-6; TNF-α: tumor necrosis factor-alpha; IL-1β: interleukin-1β; ACE receptor: Angiotensin-converting enzyme receptor; NE: norepinephrine; Ach: Acetylcholine; IDO: indoleamine 2,3-dioxygenase; α-7AchR: alpha-7 acetylcholine receptor.
: inhibit. Abbreviation: Ang II: angiotensin II; NADPH: nicotinamide adenine dinucleotide phosphate; RhoA: Ras homolog gene family member A; AT1: angiotensin type 1; CVOs: circumventricular organs; PVN: paraventricular nucleus; RVLM: rostral ventrolateral medulla; GABA: gamma-aminobutyric acid; NF-κB/AP-1: nuclear translocation of nuclear factor-kappa B/adaptor protein 1; 5-HT: 5-hydroxytryptamine; BDNF: brain-derived neurotrophic factor; ROS: reactive oxygen species; IL-6: interleukin-6; TNF-α: tumor necrosis factor-alpha; IL-1β: interleukin-1β; ACE receptor: Angiotensin-converting enzyme receptor; NE: norepinephrine; Ach: Acetylcholine; IDO: indoleamine 2,3-dioxygenase; α-7AchR: alpha-7 acetylcholine receptor.
: promote; : inhibit. Abbreviation: Ang II: angiotensin II; NADPH: nicotinamide adenine dinucleotide phosphate; RhoA: Ras homolog gene family member A; AT1: angiotensin type 1; CVOs: circumventricular organs; PVN: paraventricular nucleus; RVLM: rostral ventrolateral medulla; GABA: gamma-aminobutyric acid; NF-κB/AP-1: nuclear translocation of nuclear factor-kappa B/adaptor protein 1; 5-HT: 5-hydroxytryptamine; BDNF: brain-derived neurotrophic factor; ROS: reactive oxygen species; IL-6: interleukin-6; TNF-α: tumor necrosis factor-alpha; IL-1β: interleukin-1β; ACE receptor: Angiotensin-converting enzyme receptor; NE: norepinephrine; Ach: Acetylcholine; IDO: indoleamine 2,3-dioxygenase; α-7AchR: alpha-7 acetylcholine receptor.
 : promote;
: promote;  : inhibit. Abbreviation: nNOS: neuronal nitric oxide synthase; eNOS: endothelial nitric oxide synthase; SCFAs: short-chain fatty acids; GPCR: G-protein-coupled receptor; NO: nitric oxide; cGMP-PKG: cyclic guanosine monophosphate–protein kinase G; α1-AR: alpha-1 adrenergic receptor; α2-AR: alpha-2 adrenergic receptor; PI3K-AKT: phosphatidylinositol-3-kinase/protein kinase B; NF-κB: nuclear factor-kappa B; S-nitrosylation: S-nitrosylation of proteins.
: promote; : inhibit. Abbreviation: nNOS: neuronal nitric oxide synthase; eNOS: endothelial nitric oxide synthase; SCFAs: short-chain fatty acids; GPCR: G-protein-coupled receptor; NO: nitric oxide; cGMP-PKG: cyclic guanosine monophosphate–protein kinase G; α1-AR: alpha-1 adrenergic receptor; α2-AR: alpha-2 adrenergic receptor; PI3K-AKT: phosphatidylinositol-3-kinase/protein kinase B; NF-κB: nuclear factor-kappa B; S-nitrosylation: S-nitrosylation of proteins.
: inhibit. Abbreviation: nNOS: neuronal nitric oxide synthase; eNOS: endothelial nitric oxide synthase; SCFAs: short-chain fatty acids; GPCR: G-protein-coupled receptor; NO: nitric oxide; cGMP-PKG: cyclic guanosine monophosphate–protein kinase G; α1-AR: alpha-1 adrenergic receptor; α2-AR: alpha-2 adrenergic receptor; PI3K-AKT: phosphatidylinositol-3-kinase/protein kinase B; NF-κB: nuclear factor-kappa B; S-nitrosylation: S-nitrosylation of proteins.
: promote; : inhibit. Abbreviation: nNOS: neuronal nitric oxide synthase; eNOS: endothelial nitric oxide synthase; SCFAs: short-chain fatty acids; GPCR: G-protein-coupled receptor; NO: nitric oxide; cGMP-PKG: cyclic guanosine monophosphate–protein kinase G; α1-AR: alpha-1 adrenergic receptor; α2-AR: alpha-2 adrenergic receptor; PI3K-AKT: phosphatidylinositol-3-kinase/protein kinase B; NF-κB: nuclear factor-kappa B; S-nitrosylation: S-nitrosylation of proteins.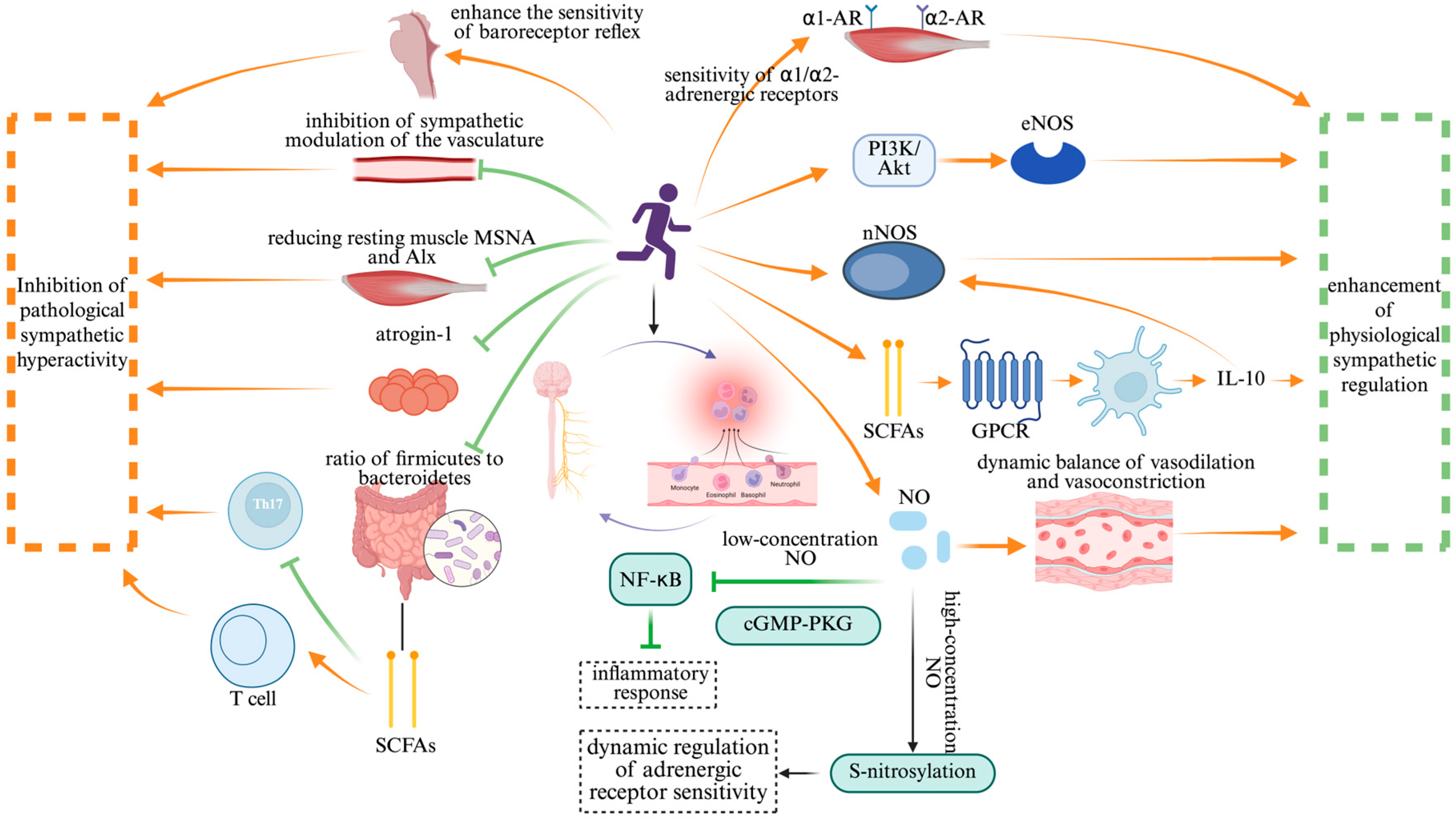
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Bai, J.; Liu, G.; Zhu, Z.; Cao, C. Exercise Intervention in Autonomic Function, Immunity, and Cardiovascular Health: A Precision Medicine Approach. J. Cardiovasc. Dev. Dis. 2025, 12, 247. https://doi.org/10.3390/jcdd12070247
Li J, Bai J, Liu G, Zhu Z, Cao C. Exercise Intervention in Autonomic Function, Immunity, and Cardiovascular Health: A Precision Medicine Approach. Journal of Cardiovascular Development and Disease. 2025; 12(7):247. https://doi.org/10.3390/jcdd12070247
Chicago/Turabian StyleLi, Jianyu, Junbei Bai, Guochun Liu, Ziyan Zhu, and Chunmei Cao. 2025. "Exercise Intervention in Autonomic Function, Immunity, and Cardiovascular Health: A Precision Medicine Approach" Journal of Cardiovascular Development and Disease 12, no. 7: 247. https://doi.org/10.3390/jcdd12070247
APA StyleLi, J., Bai, J., Liu, G., Zhu, Z., & Cao, C. (2025). Exercise Intervention in Autonomic Function, Immunity, and Cardiovascular Health: A Precision Medicine Approach. Journal of Cardiovascular Development and Disease, 12(7), 247. https://doi.org/10.3390/jcdd12070247