
The Effect of Supplementation with Weizmannia coagulans Strain SANK70258 to Coccidia-Infected Broilers Is Similar to That of a Coccidiostat Administration


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experiment 1
2.1.1. Preparation of Probiotic
2.1.2. Animals and Diets
2.1.3. Coccidial Challenge
2.1.4. Dissection and Sampling Procedure
2.1.5. Coccidial Count in Intestinal Digesta
2.1.6. Statistical Analysis
2.2. Experiment 2
2.2.1. Preparation of the Probiotic
2.2.2. Birds and Diets
2.2.3. Coccidial Challenge
2.2.4. Dissection and Sampling Procedure
2.2.5. Coccidial Count in Intestinal Digesta
2.2.6. Bacteriological Analysis
2.2.7. Histopathologic Observation
2.2.8. Statistical Analysis
3. Results
3.1. Experiment 1
3.2. Experiment 2
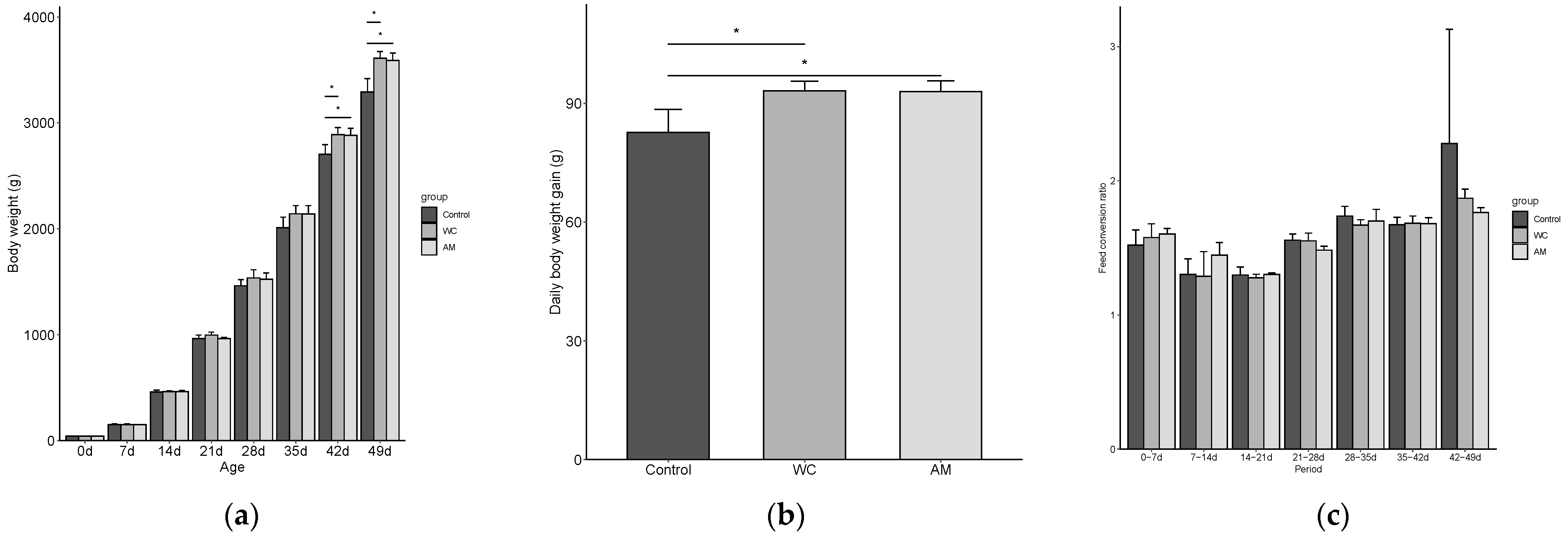
3.2.1. Effect of WC Supplementation on Production Performance of Coccidiosis Broilers
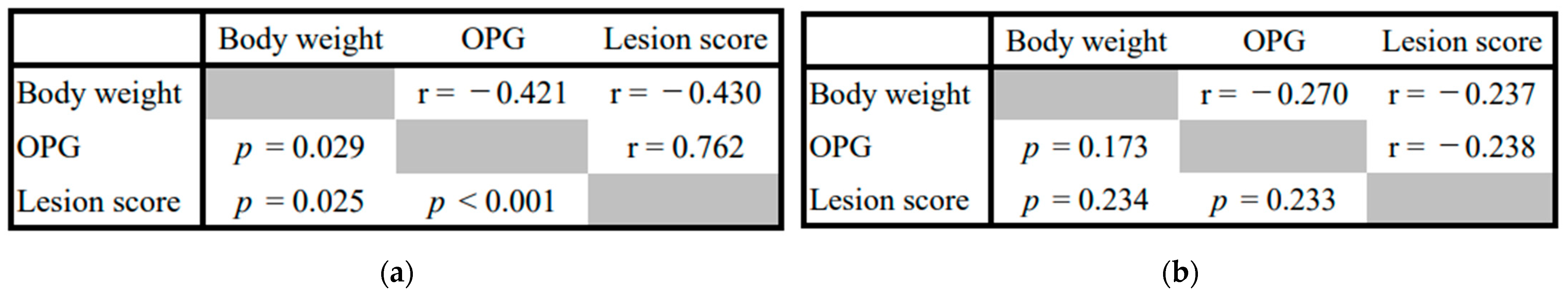
3.2.2. Effect of WC Supplementation on Intestinal Lesion Scores and Number of Oocysts of Coccidiosis Broilers
3.2.3. Effect of WC Supplementation on Total Bacteria and Escherichia coli Counts in Digesta Samples of Coccidiosis Broilers
3.2.4. The Effect of WC Supplementation on the Intestinal Morphology of Coccidiosis Broilers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chapman, H.D.; Roberts, B.; Shirley, M.W.; Williams, R.B. Guidelines for evaluating the efficacy and safety of live anticoccidial vaccines, and obtaining approval for their use in chickens and turkeys. Avian Pathol. 2005, 34, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Mohsin, M.; Zhang, Z.; Yin, G. Effect of probiotics on the performance and intestinal health of broiler chickens infected with Eimeria tenella. Vaccines 2022, 10, 97. [Google Scholar] [CrossRef] [PubMed]
- Ritzi, M.M.; Abdelrahman, W.; van-Heerden, K.; Mohnl, M.; Barrett, N.W.; Dalloul, R.A. Combination of probiotics and coccidiosis vaccine enhances protection against an Eimeria challenge. Vet. Res. 2016, 47, 111. [Google Scholar] [CrossRef]
- Blake, D.P.; Knox, J.; Dehaeck, B.; Huntington, B.; Rathinam, T.; Ravipati, V.; Ayoade, S.; Gilbert, W.; Adebambo, A.O.; Danladi Jatau, I.; et al. Re-calculating the cost of coccidiosis in chickens. Vet. Res. 2020, 51, 115. [Google Scholar] [CrossRef] [PubMed]
- Peek, H.W.; Landman, W.J. Coccidiosis in poultry: Anticoccidial products, vaccines and other prevention strategies. Vet. Q. 2011, 31, 143–161. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.; Walk, C.; Misiura, M.; Sorbara, J.-O.B.; Giannenas, I.; Kyriazakis, I. Quantifying the effect of coccidiosis on broiler performance and infection outcomes in the presence and absence of control methods. Poult. Sci. 2022, 101, 101746. [Google Scholar] [CrossRef] [PubMed]
- Boros, Z.; Băieș, M.H.; Vodnar, D.C.; Gherman, C.M.; Borșan, S.-D.; Cozma-Petruț, A.; Lefkaditis, M.; Györke, A.; Cozma, V. Antiparasitic action of Lactobacillus casei ATCC 393 and Lactobacillus paracasei CNCM strains in CD-1 mice experimentally infected with Trichinella britovi. Pathogens 2022, 11, 296. [Google Scholar] [CrossRef] [PubMed]
- Bucková, B.; Hurníková, Z.; Lauková, A.; Revajová, V.; Dvorožňáková, E. The Anti-parasitic effect of probiotic bacteria via limiting the fecundity of Trichinella spiralis female adults. Helminthologia 2018, 55, 102–111. [Google Scholar] [CrossRef]
- Myhill, L.J.; Stolzenbach, S.; Mejer, H.; Krych, L.; Jakobsen, S.R.; Kot, W.; Skovgaard, K.; Canibe, N.; Nejsum, P.; Nielsen, D.S.; et al. Parasite-probiotic interactions in the gut: Bacillus sp. and Enterococcus faecium regulate type-2 inflammatory responses and modify the gut microbiota of pigs during helminth infection. Front. Immunol. 2021, 12, 793260. [Google Scholar] [CrossRef]
- Park, I.; Lee, Y.; Goo, D.; Zimmerman, N.P.; Smith, A.H.; Rehberger, T.; Lillehoj, H.S. The effects of dietary Bacillus subtilis supplementation, as an alternative to antibiotics, on growth performance, intestinal immunity, and epithelial barrier integrity in broiler chickens infected with Eimeria maxima. Poult. Sci. 2020, 99, 725–733. [Google Scholar] [CrossRef]
- Urban, J.F., Jr.; Nielsen, M.K.; Gazzola, D.; Xie, Y.; Beshah, E.; Hu, Y.; Li, H.; Rus, F.; Flanagan, K.; Draper, A.; et al. An inactivated bacterium (paraprobiotic) expressing Bacillus thuringiensis Cry5B as a therapeutic for Ascaris and Parascaris spp. infections in large animals. One Health 2021, 12, 100241. [Google Scholar] [CrossRef]
- Wang, Y.; Lv, X.; Li, X.; Zhao, J.; Zhang, K.; Hao, X.; Liu, K.; Liu, H. Protective effect of Lactobacillus plantarum P8 on growth performance, intestinal health, and microbiota in Eimeria-infected broilers. Front. Microbiol. 2021, 12, 705758. [Google Scholar] [CrossRef]
- Yan, X.; Han, W.; Jin, X.; Sun, Y.; Gao, J.; Yu, X.; Guo, J. Study on the effect of koumiss on the intestinal microbiota of mice infected with Toxoplasma gondii. Sci. Rep. 2022, 12, 1271. [Google Scholar] [CrossRef]
- Cao, J.; Yu, Z.; Liu, W.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Probiotic characteristics of Bacillus coagulans and associated implications for human health and diseases. J. Funct. Foods 2020, 64, 103643. [Google Scholar] [CrossRef]
- Matsuzawa, T.; Iwado, S.; Kitano, N.; Suzuki, Y. The biological effects of the spore bearing lactic acid bacteria, Lactobacillus sporogenes, in chickens. Jpn. Poult. Sci. 1972, 9, 153–158. [Google Scholar] [CrossRef][Green Version]
- EFSA FEEDAP Panel (EFSA Panel on Additives and Products or Substances Used in Animal Feed); Azimonti, G.; Bampidis, V.; Bastos, M.L.; Christensen, H.; Dusemund, B.; Fašmon Durjava, M.; Kouba, M.; López-Alonso, M.; López Puente, S.; et al. Safety and efficacy of a feed additive consisting of lactic acid produced by Weizmannia coagulans (synonym Bacillus coagulans) DSM 32789 for all animal species except for fish (Jungbunzlauer SA). EFSA J. 2022, 20, e07268. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, H.; Yu, Y.; Zhang, R.; Wu, Y.; Yue, M.; Yang, C. Effects of Bacillus coagulans on growth performance, antioxidant capacity, immunity function, and gut health in broilers. Poult. Sci. 2021, 100, 101168. [Google Scholar] [CrossRef]
- Sasaki, K.; Sasaki, D.; Inoue, J.; Hoshi, N.; Maeda, T.; Yamada, R.; Kondo, A. Bacillus coagulans SANK 70258 suppresses Enterobacteriaceae in the microbiota of ulcerative colitis in vitro and enhances butyrogenesis in healthy microbiota. Appl. Microbiol. Biotechnol. 2020, 104, 3859–3867. [Google Scholar] [CrossRef]
- Faber, T.A.; Dilger, R.N.; Hopkins, A.C.; Price, N.P.; Fahey, G.C., Jr. The effects of a galactoglucomannan oligosaccharide-arabinoxylan (GGMO-AX) complex in broiler chicks challenged with Eimeria acervulina. Poult. Sci. 2012, 91, 1089–1096. [Google Scholar] [CrossRef]
- Yu, Y.; Li, Q.; Zeng, X.; Xu, Y.; Jin, K.; Liu, J.; Cao, G. Effects of probiotics on the growth performance, antioxidant functions, immune responses, and caecal microbiota of broilers challenged by lipopolysaccharide. Front. Vet. Sci. 2022, 9, 846649. [Google Scholar] [CrossRef]
- Johnson, J.; Reid, W.M. Anticoccidial drugs: Lesion scoring techniques in battery and floor-pen experiments with chickens. Exp. Parasitol. 1970, 28, 30–36. [Google Scholar] [CrossRef]
- Tsukahara, T.; Inoue, R.; Nakayama, K.; Inatomi, T. Inclusion of Bacillus amyloliquefaciens strain TOA5001 in the diet of broilers suppresses the symptoms of coccidiosis by modulating intestinal microbiota. Anim. Sci. J. 2018, 89, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Tsukahara, T.; Inoue, R.; Yamada, K.; Yajima, T. A mouse model study for the villous atrophy of the early weaning piglets. J. Vet. Med. Sci. 2010, 72, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Sinks, G.D.; Quigley, J.D., 3rd. Reinemeyer, C.R. Effects of lasalocid on coccidial infection and growth in young dairy calves. J. Am. Vet. Med. Assoc. 1992, 200, 1947–1951. [Google Scholar]
- Stromberg, B.E.; Schlotthauer, J.C.; Armstrong, B.D.; Brandt, W.E.; Liss, C. Efficacy of lasalocid sodium against coccidiosis (Eimeria zuernii and Eimeria bovis) in calves. Am. J. Vet. Res. 1982, 43, 583–585. [Google Scholar]
- Hessenberger, S.; Schatzmayr, G.; Teichmann, K. In vitro inhibition of Eimeria tenella sporozoite invasion into host cells by probiotics. Vet. Parasitol. 2016, 229, 93–98. [Google Scholar] [CrossRef]
- Pogány Simonová, M.; Chrastinová, Ľ.; Lauková, A. Autochtonous strain Enterococcus faecium EF2019(CCM7420), its bacteriocin and their beneficial effects in broiler rabbits—A review. Animals 2020, 10, 1188. [Google Scholar] [CrossRef]
- Hyronimus, B.; Le Marrec, C.; Urdaci, M.C. Coagulin, a bacteriocin-like inhibitory substance produced by Bacillus coagulans I4. J. Appl. Microbiol. 1998, 85, 42–50. [Google Scholar] [CrossRef]
- Lee, S.; Lillehoj, H.S.; Park, D.W.; Hong, Y.H.; Lin, J.J. Effects of Pediococcus- and Saccharomyces-based probiotic (MitoMax®) on coccidiosis in broiler chickens. Comp. Immunol. Microbiol. Infect. Dis. 2007, 30, 261–268. [Google Scholar] [CrossRef]
- Zhen, W.; Shao, Y.; Gong, X.; Wu, Y.; Geng, Y.; Wang, Z.; Guo, Y. Effect of dietary Bacillus coagulans supplementation on growth performance and immune responses of broiler chickens challenged by Salmonella enteritidis. Poult. Sci. 2018, 97, 2654–2666. [Google Scholar] [CrossRef]
- Fu, L.; Peng, J.; Zhao, S.; Zhang, Y.; Su, X.; Wang, Y. Lactic acid bacteria-specific induction of CD4+Foxp3+T cells ameliorates shrimp tropomyosin-induced allergic response in mice via suppression of mTOR signaling. Sci. Rep. 2017, 7, 1987. [Google Scholar] [CrossRef]
- Shinde, T.; Perera, A.P.; Vemuri, R.; Gondalia, S.V.; Karpe, A.V.; Beale, D.J.; Shastri, S.; Southam, B.; Eri, R.; Stanley, R. Synbiotic supplementation containing whole plant sugar cane fibre and probiotic spores potentiates protective synergistic effects in mouse model of IBD. Nutrients 2019, 11, 818. [Google Scholar] [CrossRef]
- Wu, Y.; Shao, Y.; Song, B.; Zhen, W.; Wang, Z.; Guo, Y.; Shahid, M.S.; Nie, W. Effects of Bacillus coagulans supplementation on the growth performance and gut health of broiler chickens with Clostridium perfringens-induced necrotic enteritis. J. Anim. Sci. Biotechnol. 2018, 9, 9. [Google Scholar] [CrossRef]
- Chaudhari, A.A.; Lee, Y.; Lillehoj, H.S. Beneficial effects of dietary supplementation of Bacillus strains on growth performance and gut health in chickens with mixed coccidiosis infection. Vet. Parasitol. 2020, 277, 109009. [Google Scholar] [CrossRef]
- Doris, V.; Gunter, K.; Kevin, D.; Sara, V.S.; Mireia, V.C.; Joao, S.; Jun, W.; Raul, Y.T.; Lindsey, D.C.; Youssef, D.; et al. Quantitative microbiome profiling links gut community variation to microbial load. Nature 2017, 551, 507–511. [Google Scholar] [CrossRef]
- Gu, S.L.; Gong, Y.; Zhang, J.; Chen, Y.; Wu, Z.; Xu, Q.; Fang, Y.; Wang, J.; Tang, L.L. Effect of the short-term use of fluoroquinolone and β-lactam antibiotics on mouse gut microbiota. Infect. Drug Resist. 2020, 13, 4547–4558. [Google Scholar] [CrossRef]
- Danzeisen, J.L.; Kim, H.B.; Isaacson, R.E.; Tu, Z.J.; Johnson, T.J. Modulations of the chicken cecal microbiome and metagenome in response to anticoccidial and growth promoter treatment. PLoS ONE 2011, 6, e27949. [Google Scholar] [CrossRef]
- Moradi, S.; Moradi, A.; Atabaigi Elmi, V.; Reza Abdollahi, M. Interactive effect of corn particle size and insoluble fiber source on performance, nutrient utilization and intestine morphology in broilers fed pelleted diets. J. Anim. Physiol. Anim. Nutr. 2021, 105, 1113–1126. [Google Scholar] [CrossRef]
- Lee, D.H.; Zo, Y.G.; Kim, S.J. Nonradioactive method to study genetic profiles of natural bacterial communities by PCR–sin- gle-strand-conformation polymorphism. Appl. Environ. Microbiol. 1996, 62, 3112–3120. [Google Scholar] [CrossRef]
- Huijsdens, X.W.; Linskens, R.K.; Mak, M.; Meuwissen, S.G.; Vandenbroucke-Grauls, C.M.; Savelkoul, P.H. Quantification of bacteria adherent to gastrointestinal mucosa by real-time PCR. J. Clin. Microbiol. 2002, 40, 4423–4427. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Parameter | C | WC_0.005 | WC_0.01 | WC_0.03 | WC_0.1 | A | p Value † |
|---|---|---|---|---|---|---|---|
| Body weight (g) | |||||||
| Day 0 | 37 ± 2 | 37 ± 2 | 36 ± 1 | 36 ± 2 | 37 ± 2 | 37 ± 2 | 0.990 |
| Day 21 | 795 ± 114 | 812 ± 99 | 810 ± 105 | 864 ± 135 | 870 ± 74 | 849 ± 81 | 0.480 |
| Day 31 | 1657 ± 205 | 1698 ± 196 | 1670 ± 176 | 1697 ± 223 | 1774 ± 159 | 1740 ± 171 | 0.768 |
| Feed conversion ratio (g/g) | |||||||
| Day 0–21 | 1.87 | 1.87 | 1.86 | 1.79 | 1.63 | 1.90 | – |
| Day 21–31 | 1.84 | 1.76 | 1.79 | 1.57 | 1.43 | 1.65 | – |
| Day 0–31 | 1.86 | 1.81 | 1.82 | 1.66 | 1.52 | 1.76 | – |
| Lesion score (score 0–4 ‡) | |||||||
| Small intestine | 1.5 ± 0.6 | 1.4 ± 0.6 | 1.6 ± 0.7 | 1.1 ± 0.4 | 1.6 ± 0.6 | 1.3 ± 0.5 | 0.298 |
| Cecum | 1.5 ± 0.6 | 1.4 ± 0.6 | 1.1 ± 0.6 | 1.3 ± 0.5 | 1.2 ± 0.4 | 1.1 ± 1.4 | 0.720 |
| Oocyst numbers in the cecum (×104 oocyst/g) | 8.66 ± 1.79 | 7.69 ± 0.88 | 6.87 ± 0.78 | 6.86 ± 1.96 | 6.64 ± 1.20 | 9.92 ± 6.34 | 0.104 |
| Intestinal length (cm) | |||||||
| Small intestine | 154.6 ± 10.8 | 161.5 ± 16.3 | 175.9 ± 18.3 | 163.9 ± 20.3 | 165.3 ± 16.4 | 164.6 ± 17.3 | 0.154 |
| Cecum | 16.3 ± 2.2 | 16.4 ± 3.1 | 16.4 ± 2.5 | 16.0 ± 2.4 | 17.0 ± 1.8 | 17.9 ± 1.8 | 0.504 |
| Parameter | Control | WC | AM | p Value ‡ |
|---|---|---|---|---|
| Body weight (g) | 1502 ± 87 | 1596 ± 92 * | 1579 ± 69 | 0.054 |
| Lesion score (score 0–4 §) | ||||
| Small intestine | 1.67 ± 0.71 | 0.89 ± 0.33 * | 0.67 ± 0.50 * | 0.001 |
| Cecum | 0.89 ± 0.60 | 0.67 ± 0.50 | 0.67 ± 0.50 | 0.603 |
| Oocyst numbers (oocyst/g) | ||||
| Small intestinal contents | 8511 ± 1609 | 6478 ± 1944 † | 5644 ± 716 * | <0.001 |
| Cecum contents | 2889 ± 909 | 2533 ± 781 | 2478 ± 342 | 0.554 |
| Total bacterial counts (log cell/g) | 10.5 ± 0.1 | 10.3 ± 0.1 * | 10.3 ± 0.2 * | <0.001 |
| E. coli levels | ||||
| Counts (log cell/g) | 6.65 ± 0.42 | 6.85 ± 0.49 | 7.56 ± 0.38 * | <0.001 |
| Ratio (%) | 0.02 ± 0.02 | 0.07 ± 0.09 | 0.27 ± 0.18 * | <0.001 |
| Intestinal length (cm) | ||||
| Small intestine | 152 ± 16 | 155 ± 15 | 154 ± 19 | 0.924 |
| Cecum | 12.0 ± 1.5 | 12.2 ± 1.4 | 12.9 ± 2.2 | 0.503 |
| Parameters | Control | WC | AM | p Value ‡ |
|---|---|---|---|---|
| Body weight (g) | 3316 ± 291 | 3538 ± 228 | 3574 ± 261 | 0.096 |
| Lesion score (score 0–4 §) | ||||
| Small intestine | 1.22 ± 0.44 | 0.89 ± 0.60 | 1.00 ± 0.71 | 0.489 |
| Cecum | 0.33 ± 0.50 | 0.33 ± 0.50 | 0.11 ± 0.33 | 0.494 |
| Oocyst numbers (oocyst/g) | ||||
| Small intestinal contents | 5867 ± 660 | 5100 ± 684 * | 4700 ± 522 * | 0.002 |
| Cecum contents | 1922 ± 156 | 1567 ± 387 * | 1578 ± 323 * | 0.033 |
| Total bacterial counts (log cell/g) | 10.6 ± 0.2 | 10.5 ± 0.1 | 10.2 ± 0.3 * | <0.001 |
| E. coli levels | ||||
| Counts (log cell/g) | 6.16 ± 0.48 | 5.90 ± 0.52 | 6.40 ± 0.44 * | 0.106 |
| Ratio (%) | 0.01 ± 0.00 | 0.00 ± 0.00 | 0.03 ± 0.02 * | 0.004 |
| Intestinal length (cm) | ||||
| Small intestine | 155 ± 10 | 165 ± 8 | 158 ± 14 | 0.139 |
| Cecum | 15.4 ± 1.7 | 15.8 ± 2.0 | 16.6 ± 1.9 | 0.460 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aida, M.; Yamada, R.; Nakamura, S.-i.; Imaoka, T.; Shimonishi, H.; Matsuo, T.; Taniguchi, I.; Tsukahara, T. The Effect of Supplementation with Weizmannia coagulans Strain SANK70258 to Coccidia-Infected Broilers Is Similar to That of a Coccidiostat Administration. Vet. Sci. 2022, 9, 406. https://doi.org/10.3390/vetsci9080406
Aida M, Yamada R, Nakamura S-i, Imaoka T, Shimonishi H, Matsuo T, Taniguchi I, Tsukahara T. The Effect of Supplementation with Weizmannia coagulans Strain SANK70258 to Coccidia-Infected Broilers Is Similar to That of a Coccidiostat Administration. Veterinary Sciences. 2022; 9(8):406. https://doi.org/10.3390/vetsci9080406
Chicago/Turabian StyleAida, Masanori, Ryouichi Yamada, Shin-ichi Nakamura, Taishi Imaoka, Hikari Shimonishi, Toshiki Matsuo, Itaru Taniguchi, and Takamitsu Tsukahara. 2022. "The Effect of Supplementation with Weizmannia coagulans Strain SANK70258 to Coccidia-Infected Broilers Is Similar to That of a Coccidiostat Administration" Veterinary Sciences 9, no. 8: 406. https://doi.org/10.3390/vetsci9080406
APA StyleAida, M., Yamada, R., Nakamura, S.-i., Imaoka, T., Shimonishi, H., Matsuo, T., Taniguchi, I., & Tsukahara, T. (2022). The Effect of Supplementation with Weizmannia coagulans Strain SANK70258 to Coccidia-Infected Broilers Is Similar to That of a Coccidiostat Administration. Veterinary Sciences, 9(8), 406. https://doi.org/10.3390/vetsci9080406