
Effects of Artemisinin on Escherichia coli–Induced Mastitis in Bovine Mammary Epithelial Cells and Mice

 ,
, 

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents and Antibodies
2.2. Bacteria Strains and Culture
2.3. Cell Culture
2.4. Mastitis Mouse Model and Sample Collection
2.5. Cell Viability Assay
2.6. Quantitative Real-Time PCR (RT-qPCR)
2.7. Western Blot
2.8. Histopathological Examination
2.9. Statistical Analysis
3. Results
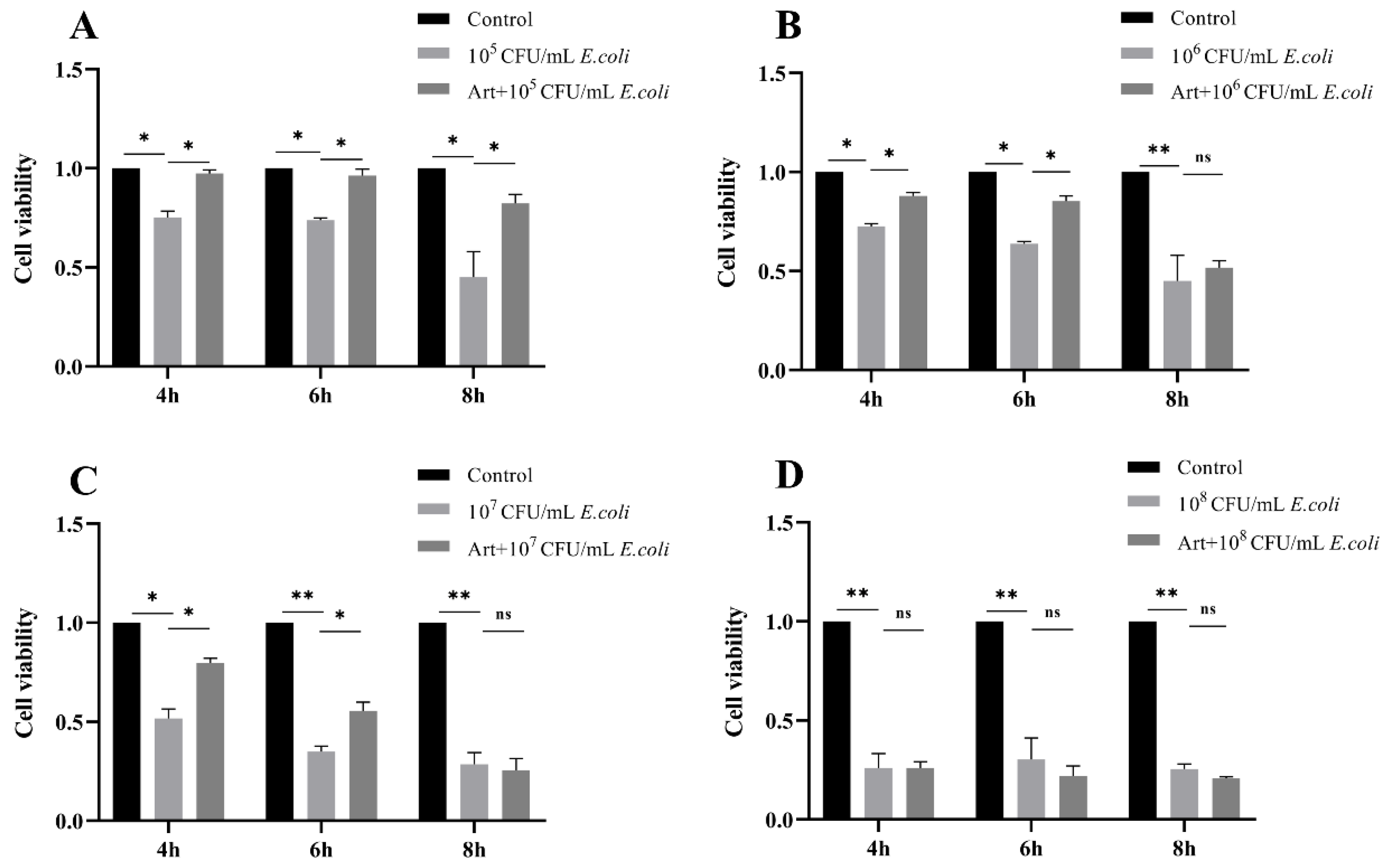
3.1. Artemisinin Reverses the Decrease of Cell Viability in E. coli–Infected MAC-T Cells
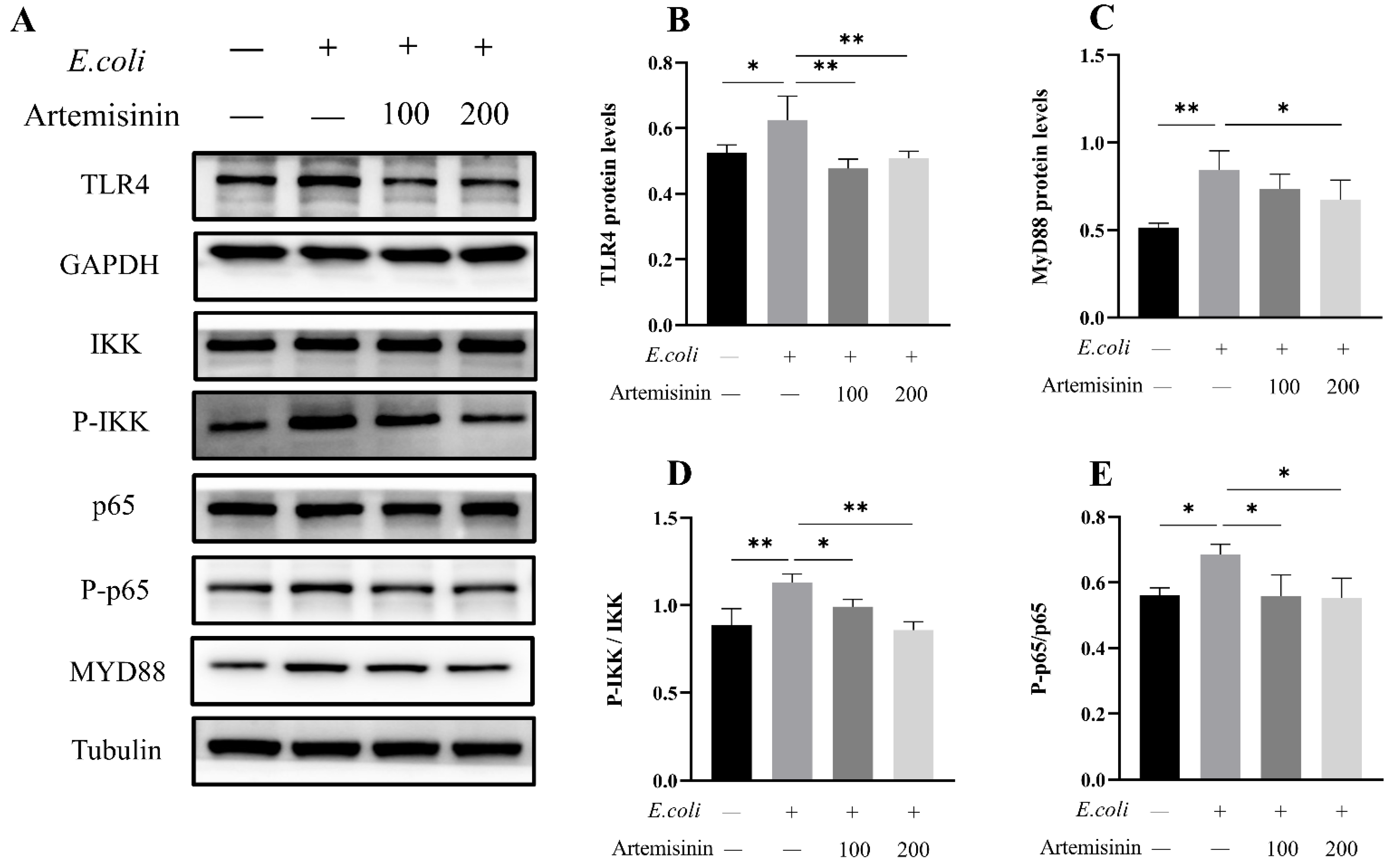
3.2. Artemisinin Inhibits the Expression of TLR4/NF-κB Inflammatory Pathway
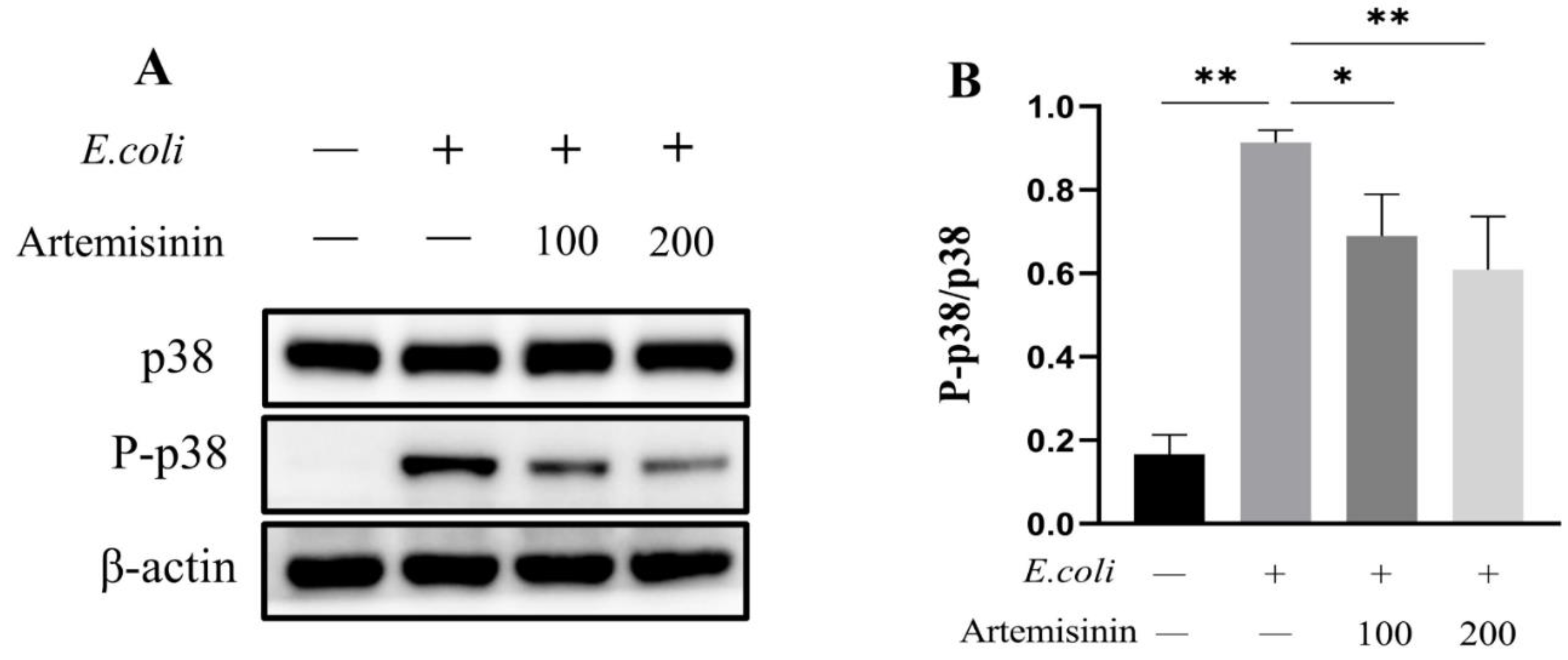
3.3. Artemisinin Inhibits the Activation of MAPK/p38 Inflammatory Pathway
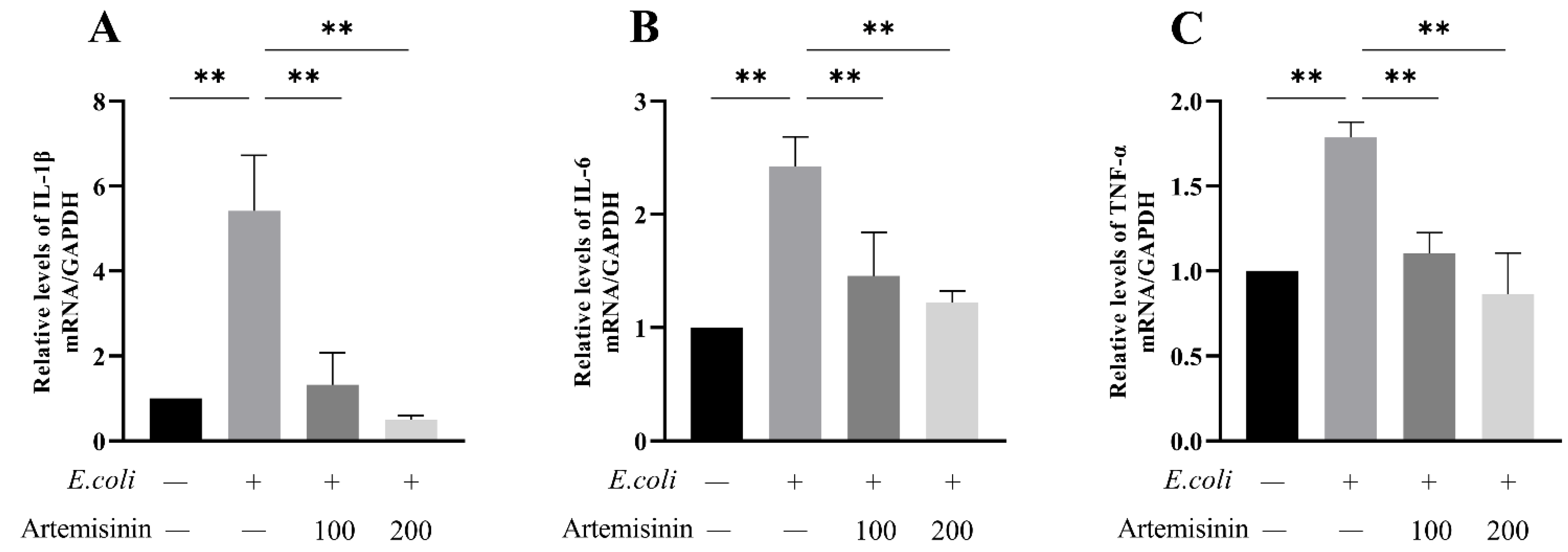
3.4. Artemisinin Reduces IL-1β, IL-6, and TNF-α mRNA Expression in E. coli–Induced MAC-T Cells
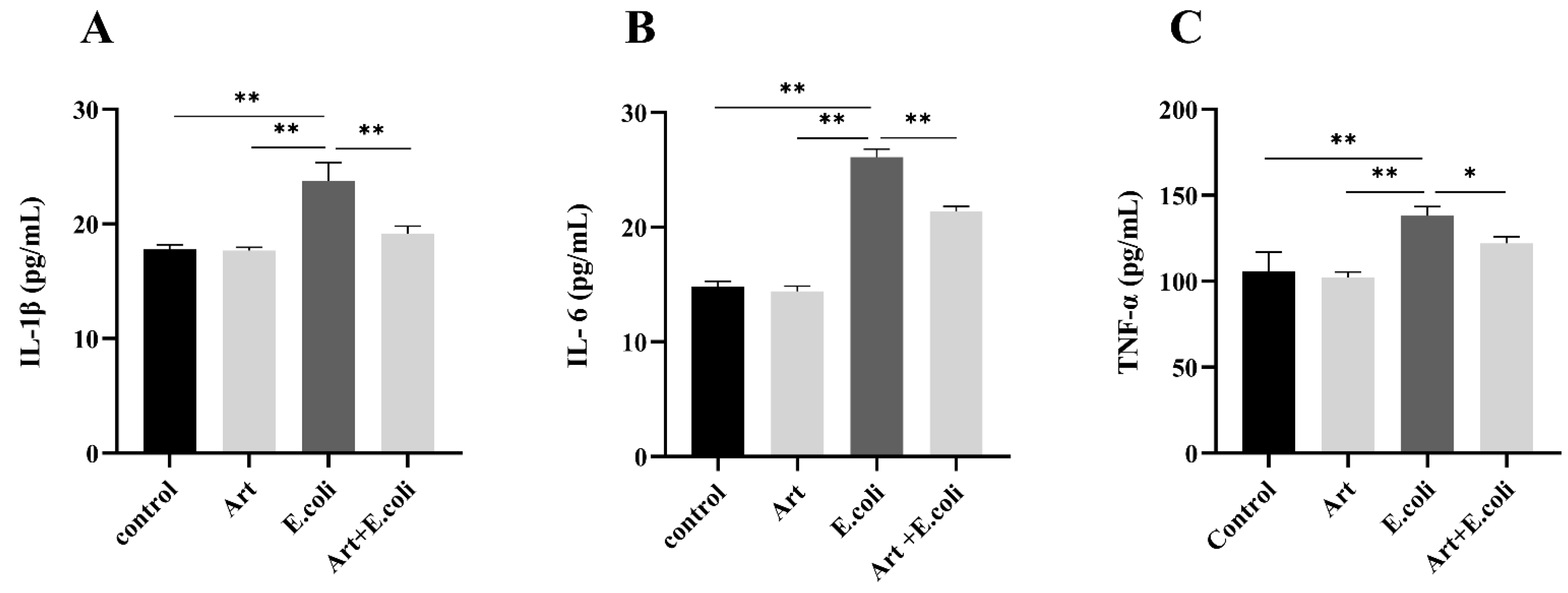
3.5. Artemisinin Reduces Serum Levels of IL-1β, IL-6 and TNF-α in Mice
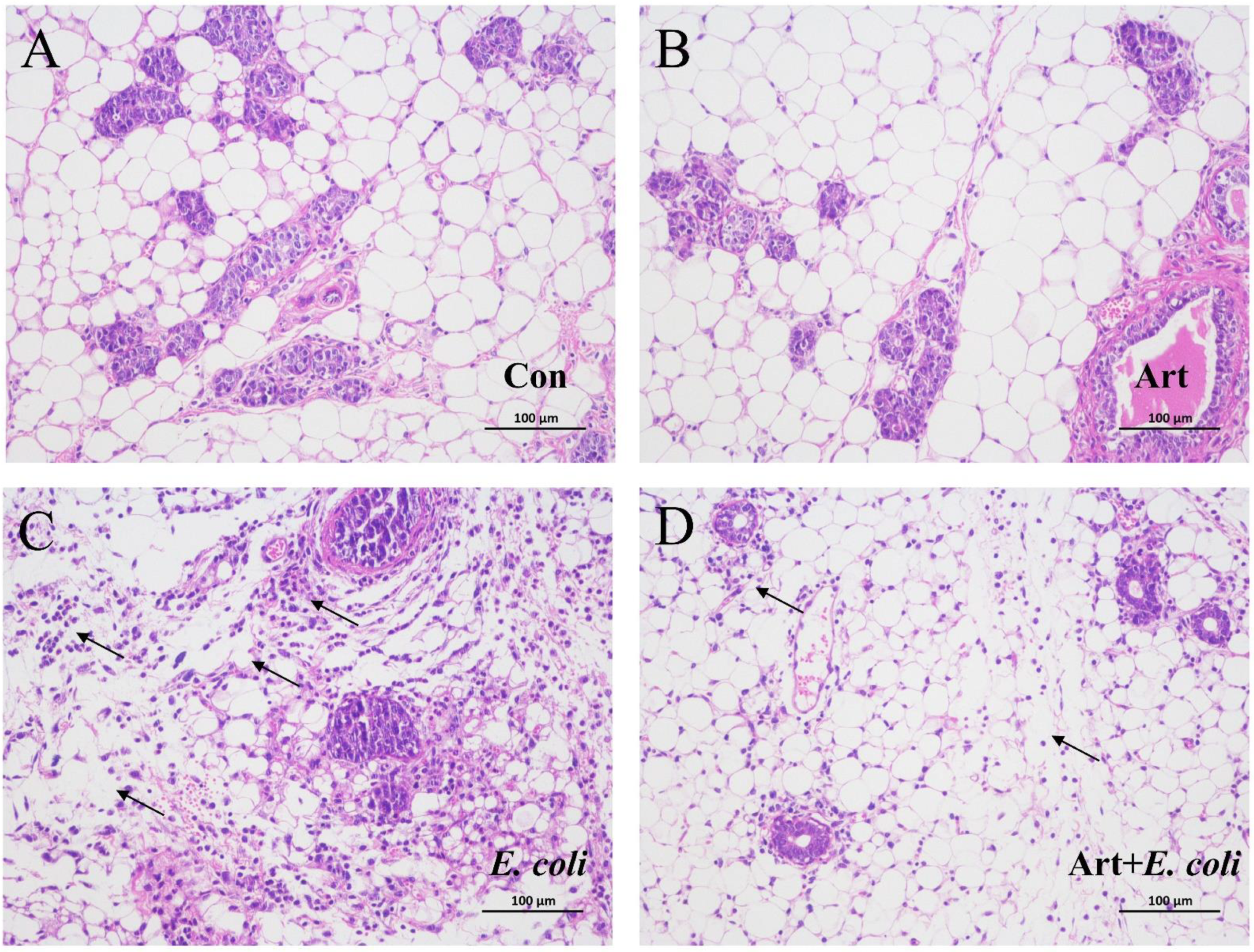
3.6. Artemisinin Relieves the Pathological Damage of Mammary Gland in E. coli–Induced Mastitis Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gomes, F.; Henriques, M. Control of Bovine Mastitis: Old and Recent Therapeutic Approaches. Curr. Microbiol. 2016, 72, 377–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riemann, T.; Bergmann, A. Epidemiology, pathogenesis, treatment and prevention of bovine acute Escherichia coli mastitis, a literature review. DTW. Dtsch. Tierarztl. Wochenschr. 2000, 107, 444–454. [Google Scholar] [PubMed]
- Sordillo, L.M.; Shafer-Weaver, K.; DeRosa, D. Immunobiology of the mammary gland. J. Dairy Sci. 1997, 80, 1851–1865. [Google Scholar] [CrossRef]
- Shimazaki, K.I.; Kawai, K. Advances in lactoferrin research concerning bovine mastitis. Biochem. Cell Biol. Biochim. Biol. Cell. 2017, 95, 69–75. [Google Scholar] [CrossRef] [Green Version]
- Kehrli, M.E., Jr.; Harp, J.A. Immunity in the mammary gland. Vet. Clin. N. Am. Food Anim. Pract. 2001, 17, 495–516. [Google Scholar] [CrossRef]
- Chaneton, L.; Tirante, L.; Maito, J.; Chaves, J.; Bussmann, L.E. Relationship between milk lactoferrin and etiological agent in the mastitic bovine mammary gland. J. Dairy Sci. 2008, 91, 1865–1873. [Google Scholar] [CrossRef] [PubMed]
- Sharun, K.; Dhama, K.; Tiwari, R.; Gugjoo, M.B.; Iqbal Yatoo, M.; Patel, S.K.; Pathak, M.; Karthik, K.; Khurana, S.K.; Singh, R.; et al. Advances in therapeutic and managemental approaches of bovine mastitis: A comprehensive review. Vet. Q. 2021, 41, 107–136. [Google Scholar] [CrossRef] [PubMed]
- Middleton, J.R.; Saeman, A.; Fox, L.K.; Lombard, J.; Hogan, J.S.; Smith, K.L. The National Mastitis Council: A Global Organization for Mastitis Control and Milk Quality, 50 Years and Beyond. J. Mammary Gland. Biol. Neoplasia 2014, 19, 241–251. [Google Scholar] [CrossRef]
- Ramos, S.; Silva, V.; Dapkevicius, M.L.E.; Caniça, M.; Tejedor-Junco, M.T.; Igrejas, G.; Poeta, P. Escherichia coli as Commensal and Pathogenic Bacteria Among Food-Producing Animals: Health Implications of Extended Spectrum β-lactamase (ESBL) Production. Animals 2020, 10, 2239. [Google Scholar] [CrossRef]
- Duse, A.; Persson-Waller, K.; Pedersen, K. Microbial Aetiology, Antibiotic Susceptibility and Pathogen-Specific Risk Factors for Udder Pathogens from Clinical Mastitis in Dairy Cows. Animals 2021, 11, 2113. [Google Scholar] [CrossRef]
- Croxen, M.A.; Finlay, B.B. Molecular mechanisms of Escherichia coli pathogenicity. Nat. Rev. Microbiol. 2010, 8, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Tang, R.; Zhao, C.; Mu, R.; Wang, Y.; Cao, Y.; Zhang, N.; Fu, Y. The Prevention Effect of Bacillus subtilis on Escherichia coli-Induced Mastitis in Mice by Suppressing the NF-kappaB and MAPK Signaling Pathways. Probiotics Antimicrob. Proteins, 2021; Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Kerro Dego, O.; Oliver, S.P.; Almeida, R.A. Host-pathogen gene expression profiles during infection of primary bovine mammary epithelial cells with Escherichia coli strains associated with acute or persistent bovine mastitis. Vet. Microbiol. 2012, 155, 291–297. [Google Scholar] [CrossRef]
- Leimbach, A.; Poehlein, A.; Vollmers, J.; Görlich, D.; Daniel, R.; Dobrindt, U. No evidence for a bovine mastitis Escherichia coli pathotype. BMC Genom. 2017, 18, 359. [Google Scholar] [CrossRef] [PubMed]
- Yair, Y.; Gophna, U. Pandemic Bacteremic Escherichia coli Strains: Evolution and Emergence of Drug-Resistant Pathogens. Curr. Top. Microbiol. Immunol. 2018, 416, 163–180. [Google Scholar] [CrossRef]
- Gao, J.; Duan, X.; Li, X.; Cao, H.; Wang, Y.; Zheng, S.J. Emerging of a highly pathogenic and multi-drug resistant strain of Escherichia coli causing an outbreak of colibacillosis in chickens. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2018, 65, 392–398. [Google Scholar] [CrossRef]
- Anes, J.; Nguyen, S.V.; Eshwar, A.K.; McCabe, E.; Macori, G.; Hurley, D.; Lehner, A.; Fanning, S. Molecular characterisation of multi-drug resistant Escherichia coli of bovine origin. Vet. Microbiol. 2020, 242, 108566. [Google Scholar] [CrossRef] [PubMed]
- Brennan, E.; Martins, M.; McCusker, M.P.; Wang, J.; Alves, B.M.; Hurley, D.; El Garch, F.; Woehrlé, F.; Miossec, C.; McGrath, L.; et al. Multidrug-Resistant Escherichia coli in Bovine Animals, Europe. Emerg. Infect. Dis. 2016, 22, 1650–1652. [Google Scholar] [CrossRef] [Green Version]
- Ma, N.; Zhang, Z.; Liao, F.; Jiang, T.; Tu, Y. The birth of artemisinin. Pharmacol. Ther. 2020, 216, 107658. [Google Scholar] [CrossRef]
- Tu, Y. Artemisinin-A Gift from Traditional Chinese Medicine to the World (Nobel Lecture). Angew. Chem. Int. Ed. 2016, 55, 10210–10226. [Google Scholar] [CrossRef]
- Ho, W.E.; Peh, H.Y.; Chan, T.K.; Wong, W.S. Artemisinins: Pharmacological actions beyond anti-malarial. Pharmacol. Ther. 2014, 142, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Efferth, T.; Oesch, F. The immunosuppressive activity of artemisinin-type drugs towards inflammatory and autoimmune diseases. Med. Res. Rev. 2021, 41, 3023–3061. [Google Scholar] [CrossRef] [PubMed]
- Kiani, B.H.; Kayani, W.K.; Khayam, A.U.; Dilshad, E.; Ismail, H.; Mirza, B. Artemisinin and its derivatives: A promising cancer therapy. Mol. Biol. Rep. 2020, 47, 6321–6336. [Google Scholar] [CrossRef] [PubMed]
- Dolivo, D.; Weathers, P.; Dominko, T. Artemisinin and artemisinin derivatives as anti-fibrotic therapeutics. Acta Pharm. Sin. B 2021, 11, 322–339. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhou, H.; Zheng, J.; Cheng, J.; Liu, W.; Ding, G.; Wang, L.; Luo, P.; Lu, Y.; Cao, H.; et al. The antimalarial artemisinin synergizes with antibiotics to protect against lethal live Escherichia coli challenge by decreasing proinflammatory cytokine release. Antimicrob. Agents Chemother. 2006, 50, 2420–2427. [Google Scholar] [CrossRef] [Green Version]
- Notebaert, S.; Meyer, E. Mouse models to study the pathogenesis and control of bovine mastitis. A review. Vet. Q. 2006, 28, 2–13. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Hu, X.; Bao, L.; Wu, K.; Feng, L.; Qiu, M.; Hao, H.; Fu, Y.; Zhang, N. Aryl hydrocarbon receptor activation by Lactobacillus reuteri tryptophan metabolism alleviates Escherichia coli-induced mastitis in mice. PLoS Pathog. 2021, 17, e1009774. [Google Scholar] [CrossRef]
- Li, Y.; Zhu, Y.; Chu, B.; Liu, N.; Chen, S.; Wang, J. Lactobacillus rhamnosus GR-1 Prevents Escherichia coli-Induced Apoptosis Through PINK1/Parkin-Mediated Mitophagy in Bovine Mastitis. Front. Immunol. 2021, 12, 715098. [Google Scholar] [CrossRef]
- Yang, F.; Zhang, S.; Shang, X.; Wang, L.; Li, H.; Wang, X. Characteristics of quinolone-resistant Escherichia coli isolated from bovine mastitis in China. J. Dairy Sci. 2018, 101, 6244–6252. [Google Scholar] [CrossRef] [Green Version]
- Dias, R.S.; Eller, M.R.; Duarte, V.S.; Pereira, Â.L.; Silva, C.C.; Mantovani, H.C.; Oliveira, L.L.; Silva Ede, A.; De Paula, S.O. Use of phages against antibiotic-resistant Staphylococcus aureus isolated from bovine mastitis. J. Anim. Sci. 2013, 91, 3930–3939. [Google Scholar] [CrossRef]
- El-Sayed, A.; Kamel, M. Bovine mastitis prevention and control in the post-antibiotic era. Trop. Anim. Health Prod. 2021, 53, 236. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhang, X.; Lin, C.; Wu, S.; Wang, F.; Wang, H.; Wang, Y.; Peng, Y.; Hutchinson, M.R.; Li, H.; et al. Artemisinin inhibits TLR4 signaling by targeting co-receptor MD2 in microglial BV-2 cells and prevents lipopolysaccharide-induced blood-brain barrier leakage in mice. J. Neurochem. 2021, 157, 611–623. [Google Scholar] [CrossRef] [PubMed]
- Qiao, S.; Zhang, H.; Sun, F.; Jiang, Z. Molecular Basis of Artemisinin Derivatives Inhibition of Myeloid Differentiation Protein 2 by Combined in Silico and Experimental Study. Molecules 2021, 26, 5698. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.S.; Choi, W.J.; Lee, S.; Kim, W.J.; Lee, D.C.; Sohn, U.D.; Shin, H.S.; Kim, W. Anti-inflammatory, Antioxidant and Antimicrobial Effects of Artemisinin Extracts from Artemisia annua L. Korean J. Physiol. Pharmacol. 2015, 19, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Bramley, A.J.; Dodd, F.H. Reviews of the progress of dairy science: Mastitis control-progress and prospects. J. Dairy Res. 1984, 51, 481–512. [Google Scholar] [CrossRef]
- Schukken, Y.H.; Günther, J.; Fitzpatrick, J.; Fontaine, M.C.; Goetze, L.; Holst, O.; Leigh, J.; Petzl, W.; Schuberth, H.J.; Sipka, A.; et al. Host-response patterns of intramammary infections in dairy cows. Vet. Immunol. Immunopathol. 2011, 144, 270–289. [Google Scholar] [CrossRef]
- Wellnitz, O.; Bruckmaier, R.M. The innate immune response of the bovine mammary gland to bacterial infection. Vet. J. 2012, 192, 148–152. [Google Scholar] [CrossRef]
- Zhang, W.; Xiong, L.; Chen, J.; Tian, Z.; Liu, J.; Chen, F.; Ren, M.; Guan, W.; Zhang, S. Artemisinin Protects Porcine Mammary Epithelial Cells against Lipopolysaccharide-Induced Inflammatory Injury by Regulating the NF-κB and MAPK Signaling Pathways. Animals 2021, 11, 1528. [Google Scholar] [CrossRef]
- Li, B.; Li, J.; Pan, X.; Ding, G.; Cao, H.; Jiang, W.; Zheng, J.; Zhou, H. Artesunate protects sepsis model mice challenged with Staphylococcus aureus by decreasing TNF-alpha release via inhibition TLR2 and Nod2 mRNA expressions and transcription factor NF-kappaB activation. Int. Immunopharmacol. 2010, 10, 344–350. [Google Scholar] [CrossRef]
- Yuan, X.; Li, J.; Li, Y.; Deng, Z.; Zhou, L.; Long, J.; Tang, Y.; Zuo, Z.; Zhang, Y.; Xie, H. Artemisinin, a potential option to inhibit inflammation and angiogenesis in rosacea. Biomed. Pharmacother. Biomed. Pharmacother. 2019, 117, 109181. [Google Scholar] [CrossRef]
- Hou, L.F.; He, S.J.; Li, X.; Yang, Y.; He, P.L.; Zhou, Y.; Zhu, F.H.; Yang, Y.F.; Li, Y.; Tang, W.; et al. Oral administration of artemisinin analog SM934 ameliorates lupus syndromes in MRL/lpr mice by inhibiting Th1 and Th17 cell responses. Arthritis Rheum. 2011, 63, 2445–2455. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.X.; Tang, W.; Zhou, R.; Wan, J.; Shi, L.P.; Zhang, Y.; Yang, Y.F.; Li, Y.; Zuo, J.P. The new water-soluble artemisinin derivative SM905 ameliorates collagen-induced arthritis by suppression of inflammatory and Th17 responses. Br. J. Pharmacol. 2008, 153, 1303–1310. [Google Scholar] [CrossRef] [Green Version]
- Ma, N.; Chang, G.; Huang, J.; Wang, Y.; Gao, Q.; Cheng, X.; Liu, J.; Shen, X. cis-9, trans-11-Conjugated Linoleic Acid Exerts an Anti-inflammatory Effect in Bovine Mammary Epithelial Cells after Escherichia coli Stimulation through NF-κB Signaling Pathway. J. Agric. Food Chem. 2019, 67, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Yue, Y.; Li, D.; Duan, C.; Qiu, X.; Zou, Y.; Zhu, Y.; Lauridsen, C.; Wang, J. Antibacterial and immunomodulatory effects of Pheromonicin-NM on Escherichia coli-challenged bovine mammary epithelial cells. Int. Immunopharmacol. 2020, 84, 106569. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Guo, C.; Wei, Z.; He, X.; Kou, J.; Zhou, E.; Yang, Z.; Fu, Y. Morin suppresses inflammatory cytokine expression by downregulation of nuclear factor-κB and mitogen-activated protein kinase (MAPK) signaling pathways in lipopolysaccharide-stimulated primary bovine mammary epithelial cells. J. Dairy Sci. 2016, 99, 3016–3022. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Zhu, Y.H.; Xu, J.; Liu, X.; Duan, C.; Wang, M.J.; Wang, J.F. Lactobacillus rhamnosus GR-1 Ameliorates Escherichia coli-Induced Activation of NLRP3 and NLRC4 Inflammasomes With Differential Requirement for ASC. Front. Microbiol. 2018, 9, 1661. [Google Scholar] [CrossRef] [Green Version]
- Ma, M.; Pei, Y.; Wang, X.; Feng, J.; Zhang, Y.; Gao, M.Q. LncRNA XIST mediates bovine mammary epithelial cell inflammatory response via NF-κB/NLRP3 inflammasome pathway. Cell Prolif. 2019, 52, e12525. [Google Scholar] [CrossRef] [Green Version]
- Machado, T.R.; Machado, T.R.; Pascutti, P.G. The p38 MAPK Inhibitors and Their Role in Inflammatory Diseases. Chem. Sel. 2021, 6, 5729–5742. [Google Scholar] [CrossRef]
- Yong, H.Y.; Koh, M.S.; Moon, A. The p38 MAPK inhibitors for the treatment of inflammatory diseases and cancer. Expert Opin. Investig. Drugs 2009, 18, 1893–1905. [Google Scholar] [CrossRef]
- Kany, S.; Vollrath, J.T.; Relja, B. Cytokines in Inflammatory Disease. Int. J. Mol. Sci. 2019, 20, 6008. [Google Scholar] [CrossRef] [Green Version]
- Zecconi, A.; Hamann, J.; Bronzo, V.; Moroni, P.; Giovannini, G.; Piccinini, R. Relationship between teat tissue immune defences and intramammary infections. Adv. Exp. Med. Biol. 2000, 480, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Reyher, K.K.; Haine, D.; Dohoo, I.R.; Revie, C.W. Examining the effect of intramammary infections with minor mastitis pathogens on the acquisition of new intramammary infections with major mastitis pathogens--a systematic review and meta-analysis. J. Dairy Sci. 2012, 95, 6483–6502. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers Name | Primers Sequence (5′→3′) |
|---|---|
| GAPDH | F: GATGGTGAAGGTCGGAGTGAAC |
| R: GTCATTGATGGCGACGATGT | |
| IL-1β | F: CCTATTCTCTCCAGCCAACCT |
| R: CTCATTCTCGTCACTGTAGTAAGC | |
| IL-6 | F: GGACTACCTCCAGAACGAGTATGA |
| R: TCTTCTCCAGCAGGTCAGTGT | |
| TNF-α | F: GCCCTCTGGTTCAAACACTCA |
| R: CGGAGAGTTGATGTCGGCTAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Hu, J.; Wang, X.; Du, Y.; Yin, J.; Gao, J.; Han, B.; Cui, S.; Liu, Y.; Liu, J. Effects of Artemisinin on Escherichia coli–Induced Mastitis in Bovine Mammary Epithelial Cells and Mice. Vet. Sci. 2022, 9, 381. https://doi.org/10.3390/vetsci9080381
Li Z, Hu J, Wang X, Du Y, Yin J, Gao J, Han B, Cui S, Liu Y, Liu J. Effects of Artemisinin on Escherichia coli–Induced Mastitis in Bovine Mammary Epithelial Cells and Mice. Veterinary Sciences. 2022; 9(8):381. https://doi.org/10.3390/vetsci9080381
Chicago/Turabian StyleLi, Zhaoming, Jiaqing Hu, Xiaozhou Wang, Yongzhen Du, Jinhua Yin, Jian Gao, Bo Han, Shuai Cui, Yongxia Liu, and Jianzhu Liu. 2022. "Effects of Artemisinin on Escherichia coli–Induced Mastitis in Bovine Mammary Epithelial Cells and Mice" Veterinary Sciences 9, no. 8: 381. https://doi.org/10.3390/vetsci9080381
APA StyleLi, Z., Hu, J., Wang, X., Du, Y., Yin, J., Gao, J., Han, B., Cui, S., Liu, Y., & Liu, J. (2022). Effects of Artemisinin on Escherichia coli–Induced Mastitis in Bovine Mammary Epithelial Cells and Mice. Veterinary Sciences, 9(8), 381. https://doi.org/10.3390/vetsci9080381