
Antimicrobial Resistant Salmonella in Chelonians: Assessing Its Potential Risk in Zoological Institutions in Spain

 ,
,  ,
,  , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Salmonella Detection and Serotyping
2.3. Antimicrobial Susceptibility Testing
2.4. Molecular Typing of Salmonella Isolates
2.5. Statistical analysis
3. Results

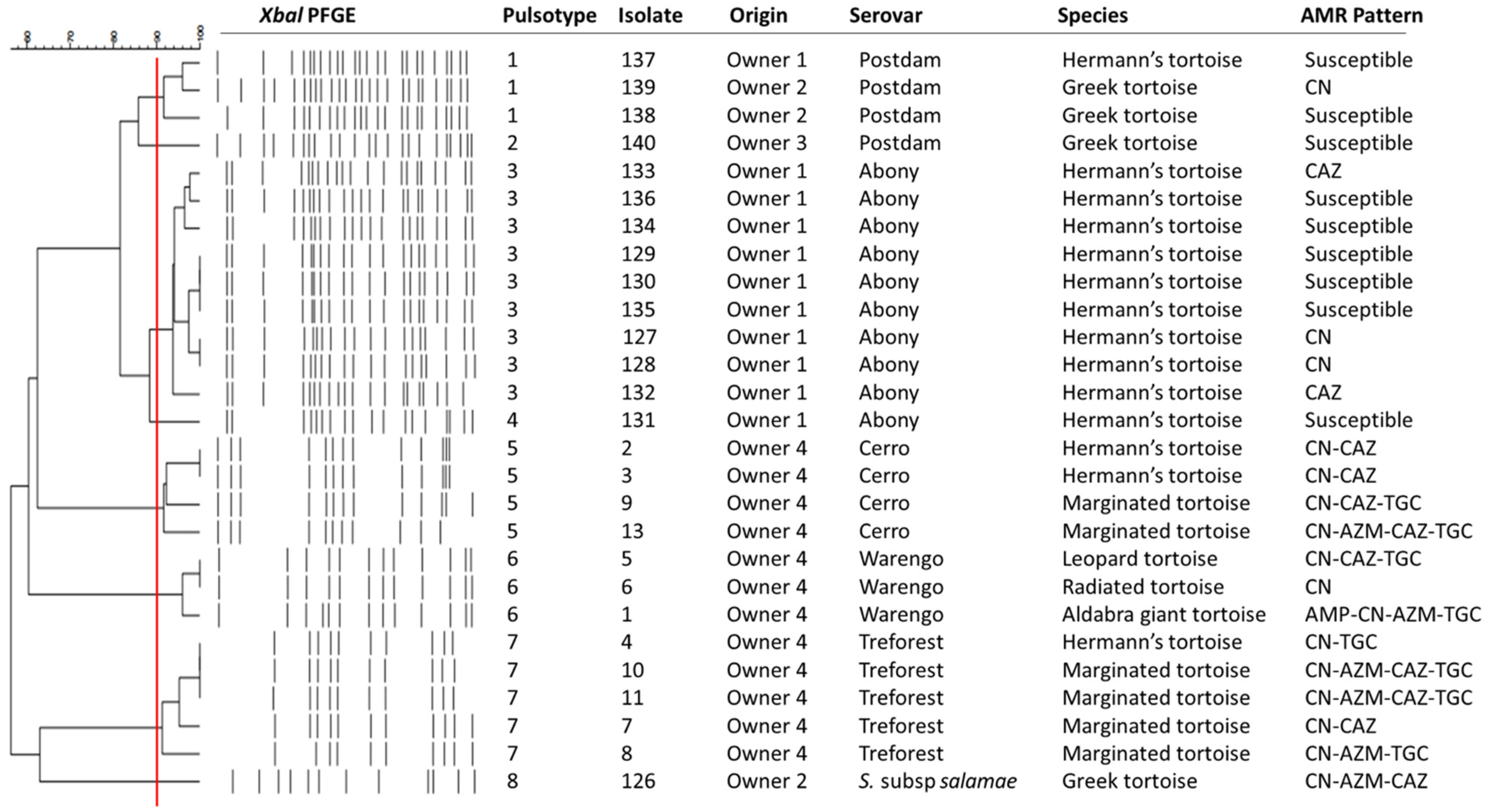{kind=link}
| Specie | Serovar | n | CIP | NA | CN | SXT | C | TM | AMP | CTX | CAZ | AZM | COL | TGC |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S. enterica subsp enterica | Abony | 10 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 |
| Postdam | 4 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Treforest | 7 | 0 | 0 | 7 | 0 | 0 | 0 | 0 | 0 | 5 | 5 | 0 | 6 | |
| Cerro | 4 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 4 | 1 | 0 | 2 | |
| Warengo | 3 | 0 | 0 | 3 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 2 | |
| S. enterica subsp salamae | 9,12:z29:1,5 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 |
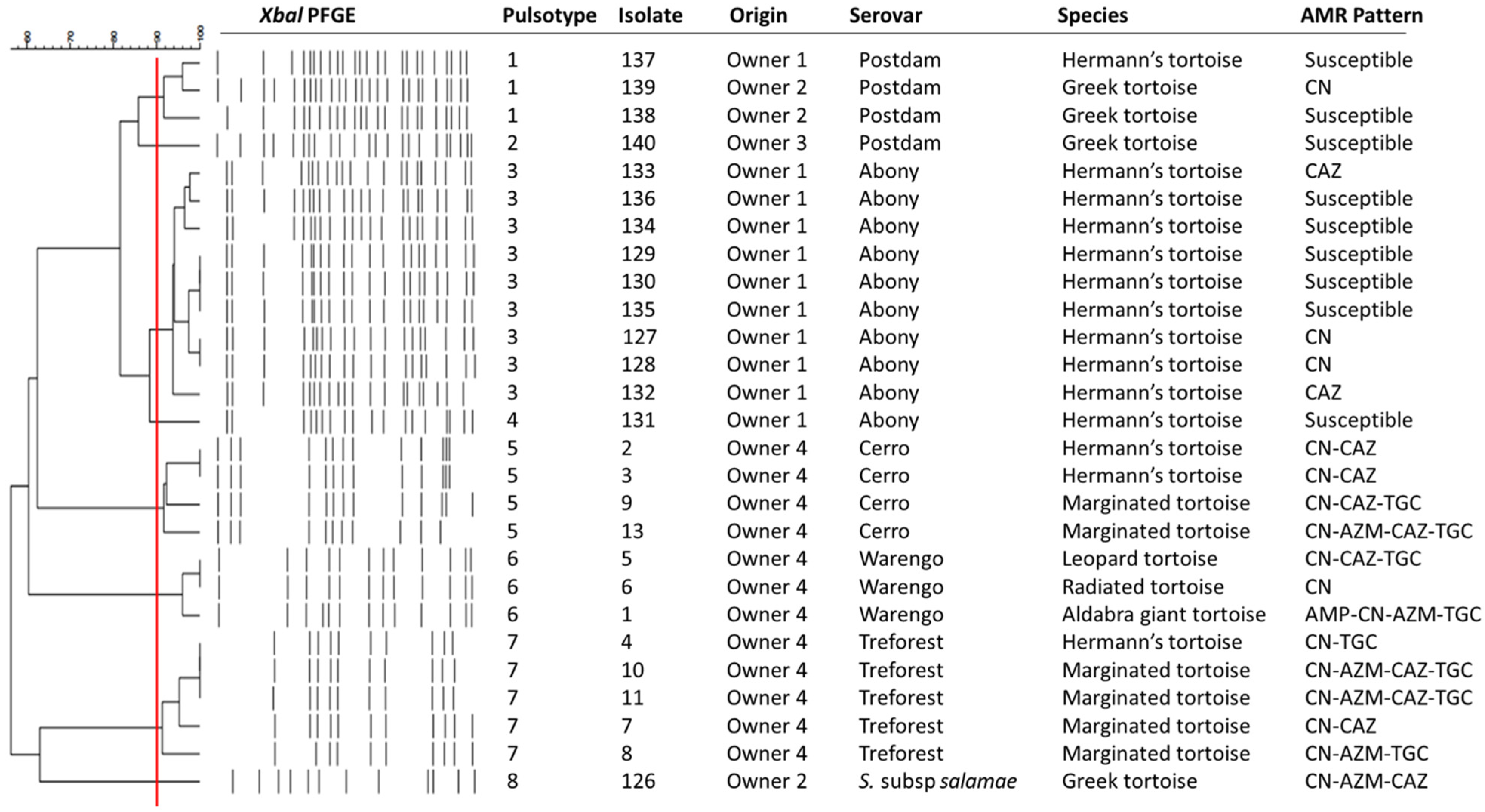
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O’Brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M. The Global Burden of Nontyphoidal Salmonella Gastroenteritis. Clin. Infect. Dis. 2010, 50, 882–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO Salmonella (Non-Typhoidal). Available online: https://www.who.int/news-room/fact-sheets/detail/salmonella-(non-typhoidal) (accessed on 22 July 2021).
- EFSA; ECDC. The European Union One Health 2019 Zoonoses Report. EFSA J. 2021, 19, 6406. [Google Scholar] [CrossRef]
- Kapoor, R.; Azad, S.M.; Mukherjee, A.; Dhar, S. Reiter’s Disease in a 8-Year-Old Boy. Indian J. Paediatr. Dermatol. 2018, 19, 154. [Google Scholar] [CrossRef]
- Hoelzer, K.; Switt, A.I.M.; Wiedmann, M. Animal Contact as a Source of Human Non-Typhoidal Salmonellosis. Vet. Res. 2011, 42, 34. [Google Scholar] [CrossRef] [Green Version]
- Kikillus, K.H.; Gartrell, B.D.; Motion, E. Prevalence of Salmonella spp., and Serovars Isolated from Captive Exotic Reptiles in New Zealand. N. Z. Vet. J. 2011, 59, 174–178. [Google Scholar] [CrossRef]
- Hernandez, S.M.; Keel, K.; Sanchez, S.; Trees, E.; Gerner-Smidt, P.; Adams, J.K.; Cheng, Y.; Ray, A.; Martin, G.; Presotto, A.; et al. Epidemiology of a Salmonella Enterica Subsp. Enterica Serovar Typhimurium Strain Associated with a Songbird Outbreak. Appl. Environ. Microbiol. 2012, 78, 7290–7298. [Google Scholar] [CrossRef] [Green Version]
- Pees, M.; Rabsch, W.; Plenz, B.; Fruth, A.; Prager, R.; Simon, S.; Schmidt, V.; Münch, S.; Braun, P.G. Evidence for the Transmission of Salmonella from Reptiles to Children in Germany, July 2010 to October 2011. Euro Surveill. 2013, 18, 20634. [Google Scholar] [CrossRef] [Green Version]
- Bjelland, A.M.; Sandvik, L.M.; Skarstein, M.M.; Svendal, L.; Debenham, J.J. Prevalence of Salmonella Serovars Isolated from Reptiles in Norwegian Zoos. Acta Vet. Scand. 2020, 62, 3. [Google Scholar] [CrossRef]
- Bertrand, S.; Rimhanen-Finne, R.; Weill, F.-X.; Rabsch, W.; Thornton, L.; Perevoščikovs, J.; van Pelt, W.; Heck, M. Salmonella Infections Associated with Reptiles: The Current Situation in Europe. Euro Surveill. 2008, 13, 18902. [Google Scholar] [CrossRef]
- Bosch, S.; Tauxe, R.V.; Behravesh, C.B. Turtle-Associated Salmonellosis, United States, 2006–2014. Emerg. Infect. Dis. 2016, 22, 1149. [Google Scholar] [CrossRef]
- Zając, M.; Skarżyńska, M.; Lalak, A.; Kwit, R.; Śmiałowska-Węglińska, A.; Pasim, P.; Szulowski, K.; Wasyl, D. Salmonella in Captive Reptiles and Their Environment—Can We Tame the Dragon? Microorganisms 2021, 9, 1012. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.L.; Potter, M.; Pollard, R.; Feldman, R.A. Turtle-Associated Salmonellosis in the United States: Effect of Public Health Action, 1970 to 1976. JAMA 1980, 243, 1247–1249. [Google Scholar] [CrossRef] [PubMed]
- De Jong, B.; Andersson, Y.; Ekdahl, K. Effect of Regulation and Education on Reptile-Associated Salmonellosis. Emerg. Infect. Dis. 2005, 11, 398–403. [Google Scholar] [CrossRef]
- EC Commission Regulation (EU) 2020/205 of 14 February 2020 Amending Regulation (EC) No 2073/2005 as Regards Salmonella in Reptile. 2020. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32020R0205&from=EN (accessed on 22 July 2021).
- Meletiadis, A.; Biolatti, C.; Mugetti, D.; Zaccaria, T.; Cipriani, R.; Pitti, M.; Decastelli, L.; Cimino, F.; Dondo, A.; Maurella, C.; et al. Surveys on Exposure to Reptile-Associated Salmonellosis (RAS) in the Piedmont Region—Italy. Animals 2022, 12, 906. [Google Scholar] [CrossRef] [PubMed]
- Corrente, M.; Sangiorgio, G.; Grandolfo, E.; Bodnar, L.; Catella, C.; Trotta, A.; Martella, V.; Buonavoglia, D. Risk for Zoonotic Salmonella Transmission from Pet Reptiles: A Survey on Knowledge, Attitudes and Practices of Reptile-Owners Related to Reptile Husbandry. Prev. Vet. Med. 2017, 146, 73–78. [Google Scholar] [CrossRef]
- Hidalgo-Vila, J.; Díaz-Paniagua, C.; de Frutos-Escobar, C.; Jiménez-Martínez, C.; Pérez-Santigosa, N. Salmonella in Free Living Terrestrial and Aquatic Turtles. Vet. Microbiol. 2007, 119, 311–315. [Google Scholar] [CrossRef] [Green Version]
- Marin, C.; Ingresa-Capaccioni, S.; González-Bodi, S.; Marco-Jiménez, F.; Vega, S. Free-Living Turtles Are a Reservoir for Salmonella but Not for Campylobacter. PLoS ONE 2013, 8, e72350. [Google Scholar] [CrossRef] [Green Version]
- Nakadai, A.; Kuroki, T.; Kato, Y.; Suzuki, R.; Yamai, S.; Yaginuma, C.; Shiotani, R.; Yamanouchi, A.; Hayashidani, H. Prevalence of Salmonella Spp. in Pet Reptiles in Japan. J. Vet. Med. Sci. 2005, 67, 97–101. [Google Scholar] [CrossRef] [Green Version]
- Marin, C.; Vega, S.; Marco-Jiménez, F. Tiny Turtles Purchased at Pet Stores Are a Potential High Risk for Salmonella Human Infection in the Valencian Region, Eastern Spain. Vector Borne Zoonotic Dis. 2016, 16, 455–460. [Google Scholar] [CrossRef]
- Marin, C.; Lorenzo-Rebenaque, L.; Laso, O.; Villora-Gonzalez, J.; Vega, S. Pet Reptiles: A Potential Source of Transmission of Multidrug-Resistant Salmonella. Front. Vet. Sci. 2021, 7, 613718. [Google Scholar] [CrossRef]
- Cota, J.B.; Carvalho, A.C.; Dias, I.; Reisinho, A.; Bernardo, F.; Oliveira, M. Salmonella Spp. in Pet Reptiles in Portugal: Prevalence and Chlorhexidine Gluconate Antimicrobial Efficacy. Antibiotics 2021, 10, 324. [Google Scholar] [CrossRef]
- La Tela, I.; Peruzy, M.F.; D’Alessio, N.; di Nocera, F.; Casalinuovo, F.; Carullo, M.R.; Cardinale, D.; Cristiano, D.; Capuano, F. Serotyping and Evaluation of Antimicrobial Resistance of Salmonella Strains Detected in Wildlife and Natural Environments in Southern Italy. Antibiotics 2021, 10, 353. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority. The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-Borne Outbreaks in 2017. EFSA J. 2018, 16, e05500. [Google Scholar] [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, Research, and Development of New Antibiotics: The WHO Priority List of Antibiotic-Resistant Bacteria and Tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Threlfall, E.J.; Fisher, I.S.T.; Berghold, C.; Gerner-Smidt, P.; Tschäpe, H.; Cormican, M.; Luzzi, L.; Schneider, F.; Wannet, W.; Machado, J.; et al. Antimicrobial Drug Resistance in Isolates of Salmonella Enterica from Cases of Salmonellosis in Humans in Europe in 2000: Results of International Multi-Centre Surveillance. Eurosurveillance 2003, 8, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Ferstl, P.G.; Reinheimer, C.; Jozsa, K.; Zeuzem, S.; Kempf, V.A.J.; Waidmann, O.; Grammatikos, G. Severe Infection with Multidrug-Resistant Salmonella Choleraesuis in a Young Patient with Primary Sclerosing Cholangitis. World J. Gastroenterol. 2017, 23, 2086–2089. [Google Scholar] [CrossRef] [PubMed]
- Ebani, V.; Cerri, D.; Fratini, F.; Meille, N.; Valentini, P.; Andreani, E. Salmonella Enterica Isolates from Faeces of Domestic Reptiles and a Study of Their Antimicrobial in Vitro Sensitivity. Res. Vet. Sci. 2005, 78, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Conrad, C.; Stanford, K.; Narvaez-Bravo, C.; Callaway, T.; McAllister, T. Farm Fairs and Petting Zoos: A Review of Animal Contact as a Source of Zoonotic Enteric Disease. Foodborne Pathog. Dis. 2017, 14, 59–73. [Google Scholar] [CrossRef]
- Casalino, G.; Bellati, A.; Pugliese, N.; Camarda, A.; Faleo, S.; Lombardi, R.; Occhiochiuso, G.; D’Onghia, F.; Circella, E. Salmonella Infection in Turtles: A Risk for Staff Involved in Wildlife Management? Animals 2021, 11, 1529. [Google Scholar] [CrossRef]
- Por El Que Se Establecen Las Normas Básicas Aplicables Para La Protección de Los Animales Utilizados En Experimentación y Otros Fines Científicos, Incluyendo La Docencia; Spain Spain 2013. Royal Degree 53/1 February 2013; Boletín Oficial del Estado: Madrid, Spain, 2013; pp. 11370–11421.
- Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Detection of Salmonella spp. Available online: https://www.iso.org/standard/29315.html (accessed on 7 October 2021).
- Grimont, P.A.D.; Weill, F.X. Antigenic Formulae of the Salmonella Serovars, 9th ed.; WHO Collaborating Centre for Reference and Research on Salmonella: Paris, France, 2007. [Google Scholar]
- Matuschek, E.; Brown, D.F.J.; Kahlmeter, G. Development of the EUCAST Disk Diffusion Antimicrobial Susceptibility Testing Method and Its Implementation in Routine Microbiology Laboratories. Clin. Microbiol. Infect. 2014, 20, O255–O266. [Google Scholar] [CrossRef] [Green Version]
- Boletín Oficial del Estado Decisión 2013/652/UE, 12 de Noviembre Sobre El Seguimiento y La Notificación de La Resistencia de Las Bacterias Zoonóticas y Comensales a Los Antibióticos. 2013, pp. 26–39. Available online: https://www.boe.es/doue/2013/303/L00026-00039.pdf (accessed on 22 July 2021).
- EUCAST New Definitions of S, I and R from 2019. Available online: https://www.eucast.org/newsiandr/ (accessed on 7 October 2021).
- EFSA; ECDC. The European Union Summary Report on Antimicrobial Resistance in Zoonotic and Indicator Bacteria from Humans, Animals and Food in 2017. EFSA J. 2019, 17, e05598. [Google Scholar] [CrossRef]
- Whitten, T.; Bender, J.B.; Smith, K.; Leano, F.; Scheftel, J. Reptile-Associated Salmonellosis in Minnesota, 1996–2011. Zoonoses Public Health 2015, 62, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Kanagarajah, S.; Waldram, A.; Dolan, G.; Jenkins, C.; Ashton, P.M.; Martin, A.I.C.; Davies, R.; Frost, A.; Dallman, T.J.; de Pinna, E.M.; et al. Whole Genome Sequencing Reveals an Outbreak of Salmonella Enteritidis Associated with Reptile Feeder Mice in the United Kingdom, 2012–2015. Food Microbiol. 2018, 71, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Gambino-Shirley, K.; Stevenson, L.; Concepción-Acevedo, J.; Trees, E.; Wagner, D.; Whitlock, L.; Roberts, J.; Garrett, N.; van Duyne, S.; McAllister, G.; et al. Flea Market Finds and Global Exports: Four Multistate Outbreaks of Human Salmonella Infections Linked to Small Turtles, United States—2015. Zoonoses Public Health 2018, 65, 560–568. [Google Scholar] [CrossRef]
- Gay, N.; le Hello, S.; Weill, F.-X.; de Thoisy, B.; Berger, F. Salmonella Serotypes in Reptiles and Humans, French Guiana. Vet. Microbiol. 2014, 170, 167–171. [Google Scholar] [CrossRef] [Green Version]
- Russo, T.P.; Varriale, L.; Borrelli, L.; Pace, A.; Latronico, M.; Menna, L.F.; Fioretti, A.; Dipineto, L. Salmonella Serotypes Isolated in Geckos Kept in Seven Collections in Southern Italy. J. Small Anim. Pract. 2018, 59, 294–297. [Google Scholar] [CrossRef]
- Chiodini, R.J.; Sundberg, J.P. Salmonellosis in Reptiles: A Review. Am. J. Epidemiol. 1981, 113, 494–499. [Google Scholar] [CrossRef]
- Strohl, P.; Tilly, B.; Frémy, S.; Brisabois, A.; Guérin-Faublée, V. Prevalence of Salmonella Shedding in Faeces by Captive Chelonians. Vet. Rec. 2004, 154, 56–58. [Google Scholar] [CrossRef]
- Percipalle, M.; Giardina, G.; Lipari, L.; Piraino, C.; Macrì, D.; Ferrantelli, V. Salmonella Infection in Illegally Imported Spur-Thighed Tortoises (Testudo graeca). Zoonoses Public Health 2011, 58, 262–269. [Google Scholar] [CrossRef]
- Candela, M.G.; Atance, P.M.; Seva, J.; Pallarés, F.J.; Vizcaíno, L.L. Granulomatous Hepatitis Caused by Salmonella Typhimurium in a Spur-Thighed Tortoise (Testudo graeca). Vet. Rec. 2005, 157, 236–237. [Google Scholar] [CrossRef] [PubMed]
- Pasmans, F.; Blahak, S.; Martel, A.; Pantchev, N. Introducing Reptiles into a Captive Collection: The Role of the Veterinarian. Vet. J. 2008, 175, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Tomastikova, Z.; Romero, S.B.; Knotek, Z.; Karpiskova, R. Prevalence and Characteristics of Salmonella Species Isolated from Captive Reptiles in the Czech Republic. Orig. Pap. Vet. Med. 2017, 62, 456–469. [Google Scholar] [CrossRef] [Green Version]
- Richards, J.M.; Brown, J.D.; Kelly, T.R.; Fountain, A.L.; Sleeman, J.M. Absence of Detectable Salmonella Cloacal Shedding in Free-Living Reptiles on Admission to the Wildlife Center of Virginia. J. Zoo Wildl. Med. 2004, 35, 562–563. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Chen, W.-C.; Chin, S.-C.; Lai, Y.-H.; Tung, K.-C.; Chiou, C.-S.; Hsu, Y.-M.; Chang, C.-C. Prevalence and Antimicrobial Susceptibility of Salmonellae Isolates from Reptiles in Taiwan. J. Vet. Diagn. Investig. 2010, 22, 44–50. [Google Scholar] [CrossRef] [Green Version]
- Pitiriga, V.; Dendrinos, J.; Nikitiadis, E.; Vrioni, G.; Tsakris, A. First Case of Lung Abscess Due to Salmonella Enterica Serovar Abony in an Immunocompetent Adult Patient. Case Rep. Infect. Dis. 2016, 2016, 3159031. [Google Scholar] [CrossRef] [Green Version]
- Van Meervenne, E.; Botteldoorn, N.; Lokietek, S.; Vatlet, M.; Cupa, A.; Naranjo, M.; Dierick, K.; Bertrand, S. Turtle-Associated Salmonella Septicaemia and Meningitis in a 2-Month-Old Baby. J. Med. Microbiol. 2009, 58, 1379–1381. [Google Scholar] [CrossRef]
- Gicquelais, R.E.; Morris, J.F.; Matthews, H.S.; Gladden, L.; Safi, H.; Grayson, C.; Slayton, R.B.; Newton, A.E.; Bordonaro, R.; Wheeler, J.G.; et al. Multiple-Serotype Salmonella Outbreaks in Two State Prisons—Arkansas, August 2012. MMWR. Morb. Mortal. Wkly. Rep. 2014, 63, 169. [Google Scholar]
- Cheng, W.; Lian, K.; Luo, D.; Lin, D.; Feng, W.; Xian, H.; Li, T. Salmonella Potsdam Causing Lumbar Vertebral Osteomyelitis: A Case Report. Medicine 2018, 97, e0682. [Google Scholar] [CrossRef]
- Briones, V.; Téllez, S.; Goyache, J.; Ballesteros, C.; Lanzarot, M.D.P.; Domínguez, L.; Fernández-Garayzábal, J.F. Salmonella Diversity Associated with Wild Reptiles and Amphibians in Spain. Environ. Microbiol. 2004, 6, 868–871. [Google Scholar] [CrossRef]
- Holmes, A.H.; Moore, L.S.P.; Sundsfjord, A.; Steinbakk, M.; Regmi, S.; Karkey, A.; Guerin, P.J.; Piddock, L.J.V. Understanding the Mechanisms and Drivers of Antimicrobial Resistance. Lancet 2016, 387, 176–187. [Google Scholar] [CrossRef]
- Forsberg, K.J.; Reyes, A.; Wang, B.; Selleck, E.M.; Sommer, M.O.A.; Dantas, G. The Shared Antibiotic Resistome of Soil Bacteria and Human Pathogens. Science 2012, 337, 1107–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, L.J.; Harris, E.; Williams, M.; Ryan, J.J.; Kookana, R.S.; Boxall, A.B.A. Fate and Uptake of Pharmaceuticals in Soil—Plant Systems. J. Agric. Food Chem. 2014, 62, 816–825. [Google Scholar] [CrossRef]
- Baquero, F.; Martínez, J.L.; Cantón, R. Antibiotics and Antibiotic Resistance in Water Environments. Curr. Opin. Biotechnol. 2008, 19, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Giacopello, C.; Foti, M.; Fisichella, V.; Latella, G.; Aleo, A.; Mammina, C. Antibiotic Resistance in Salmonella Isolated from Tegus (Tupinambis spp.). J. Exot. Pet Med. 2012, 21, 328–331. [Google Scholar] [CrossRef]
- Gibbons, P.M.; Steffes, Z.J. Emerging Infectious Diseases of Chelonians. Vet. Clin. Exot. Anim. Pract. 2013, 16, 303–317. [Google Scholar] [CrossRef]
- Cerreta, A.J.; Masterson, C.A.; Lewbart, G.A.; Dise, D.R.; Papich, M.G. Pharmacokinetics of Ketorolac in Wild Eastern Box Turtles (Terrapene carolina carolina) after Single Intramuscular Administration. J. Vet. Pharmacol. Ther. 2019, 42, 154–159. [Google Scholar] [CrossRef]
- Collignon, P.; Aarestrup, F.M. Extended-Spectrum β-Lactamases, Food, and Cephalosporin Use in Food Animals. Clin. Infect. Dis. 2007, 44, 1391–1392. [Google Scholar] [CrossRef]
- Clancy, M.M.; Davis, M.; Valitutto, M.T.; Nelson, K.; Sykes, J.M. Salmonella Infection and Carriage in Reptiles in a Zoological Collection. J. Am. Vet. Med. Assoc. 2016, 248, 1050–1059. [Google Scholar] [CrossRef]
- Bertelloni, F.; Chemaly, M.; Cerri, D.; Le Gall, F.; Ebani, V.V. Salmonella Infection in Healthy Pet Reptiles: Bacteriological Isolation and Study of Some Pathogenic Characters. Acta Microbiol. Et Immunol. Hung. 2016, 63, 203–216. [Google Scholar] [CrossRef] [Green Version]
- Sajib, M.; Tanmoy, A.M.; Hooda, Y.; Rahman, H.; Andrews, J.R.; Garrett, D.O.; Endtz, H.P.; Saha, S.K.; Saha, S. Tracking the Emergence of Azithromycin Resistance in Multiple Genotypes of Typhoidal Salmonella. mBio 2021, 12, e03481-20. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Schneiders, T. Tigecycline Challenge Triggers SRNA Production in Salmonella Enterica Serovar Typhimurium. BMC Microbiol. 2012, 12, 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tardón, A.; Bataller, E.; Llobat, L.; Jiménez-Trigos, E. Bacteria and Antibiotic Resistance Detection in Fractures of Wild Birds from Wildlife Rehabilitation Centres in Spain. Comp. Immunol. Microbiol. Infect. Dis. 2021, 74, 101575. [Google Scholar] [CrossRef] [PubMed]
- Hsu, B.B.; Gibson, T.E.; Yeliseyev, V.; Liu, Q.; Lyon, L.; Bry, L.; Silver, P.A.; Gerber, G.K. Dynamic Modulation of the Gut Microbiota and Metabolome by Bacteriophages in a Mouse Model. Cell Host Microbe 2019, 25, 803–814.e5. [Google Scholar] [CrossRef] [Green Version]
| Zoological Nucleus | Chelonian Species | n | Origin | |
|---|---|---|---|---|
| Captivity | Nature | |||
| GREFA | Greek tortoises (Testudo graeca) | 16 | 16 | 0 |
| Hermann’s tortoises (Testudo hermanni) | 24 | 22 | 2 | |
| European pond turtles (Emys orbicularis) | 18 | 18 | 0 | |
| Mediterranean pond turtles (Mauremys leprosa) | 19 | 10 | 9 | |
| OCE | Marginated tortoise (Testudo marginata) | 20 | 20 | 0 |
| Hermann’s tortoises (Testudo hermanni) | 3 | 3 | 3 | |
| Radiated tortoise (Astrochelys radiata) | 14 | 14 | 0 | |
| Aldabra giant tortoise (Aldabrachelys gigantea) | 2 | 2 | 0 | |
| Leopard tortoise (Stigmochelys pardalis) | 2 | 2 | 0 | |
| Loggerhead sea turtle (Caretta caretta) | 34 | 0 | 34 | |
| Zoological Nucleus | Chelonian Type | Chelonian Species | n | Salmonella-Positive (%) |
|---|---|---|---|---|
| GREFA | Tortoise | Greek tortoise (Testudo graeca) | 16 | 25 |
| Hermann’s tortoise (Testudo hermanni) | 24 | 45.8 | ||
| Pond Turtle | European pond turtle (Emys orbicularis) | 18 | 0 | |
| Mediterranean pond turtle (Mauremys leprosa) | 19 | 0 | ||
| OCE | Tortoise | Marginated tortoise (Testudo marginata) | 20 | 40 |
| Hermann’s tortoises (Testudo hermanni) | 3 | 100 | ||
| Radiated tortoise (Astrochelys radiata) | 14 | 100 | ||
| Aldabra giant tortoise (Aldabrachelys gigantea) | 2 | 50 | ||
| Leopard tortoise (Stigmochelys pardalis) | 2 | 50 | ||
| Sea Turtles | Loggerhead sea turtle (Caretta caretta) | 34 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marin, C.; Martín-Maldonado, B.; Cerdà-Cuéllar, M.; Sevilla-Navarro, S.; Lorenzo-Rebenaque, L.; Montoro-Dasi, L.; Manzanares, A.; Ayats, T.; Mencía-Gutiérrez, A.; Jordá, J.; et al. Antimicrobial Resistant Salmonella in Chelonians: Assessing Its Potential Risk in Zoological Institutions in Spain. Vet. Sci. 2022, 9, 264. https://doi.org/10.3390/vetsci9060264
Marin C, Martín-Maldonado B, Cerdà-Cuéllar M, Sevilla-Navarro S, Lorenzo-Rebenaque L, Montoro-Dasi L, Manzanares A, Ayats T, Mencía-Gutiérrez A, Jordá J, et al. Antimicrobial Resistant Salmonella in Chelonians: Assessing Its Potential Risk in Zoological Institutions in Spain. Veterinary Sciences. 2022; 9(6):264. https://doi.org/10.3390/vetsci9060264
Chicago/Turabian StyleMarin, Clara, Bárbara Martín-Maldonado, Marta Cerdà-Cuéllar, Sandra Sevilla-Navarro, Laura Lorenzo-Rebenaque, Laura Montoro-Dasi, Alicia Manzanares, Teresa Ayats, Aida Mencía-Gutiérrez, Jaume Jordá, and et al. 2022. "Antimicrobial Resistant Salmonella in Chelonians: Assessing Its Potential Risk in Zoological Institutions in Spain" Veterinary Sciences 9, no. 6: 264. https://doi.org/10.3390/vetsci9060264
APA StyleMarin, C., Martín-Maldonado, B., Cerdà-Cuéllar, M., Sevilla-Navarro, S., Lorenzo-Rebenaque, L., Montoro-Dasi, L., Manzanares, A., Ayats, T., Mencía-Gutiérrez, A., Jordá, J., González, F., Rojo-Solís, C., Barros, C., García-Párraga, D., & Vega, S. (2022). Antimicrobial Resistant Salmonella in Chelonians: Assessing Its Potential Risk in Zoological Institutions in Spain. Veterinary Sciences, 9(6), 264. https://doi.org/10.3390/vetsci9060264