
Molecular Characterization and Phylogenetic Analysis of Spirometra Tapeworms from Snakes in Hunan Province




Abstract
:1. Introduction
2. Materials and Methods
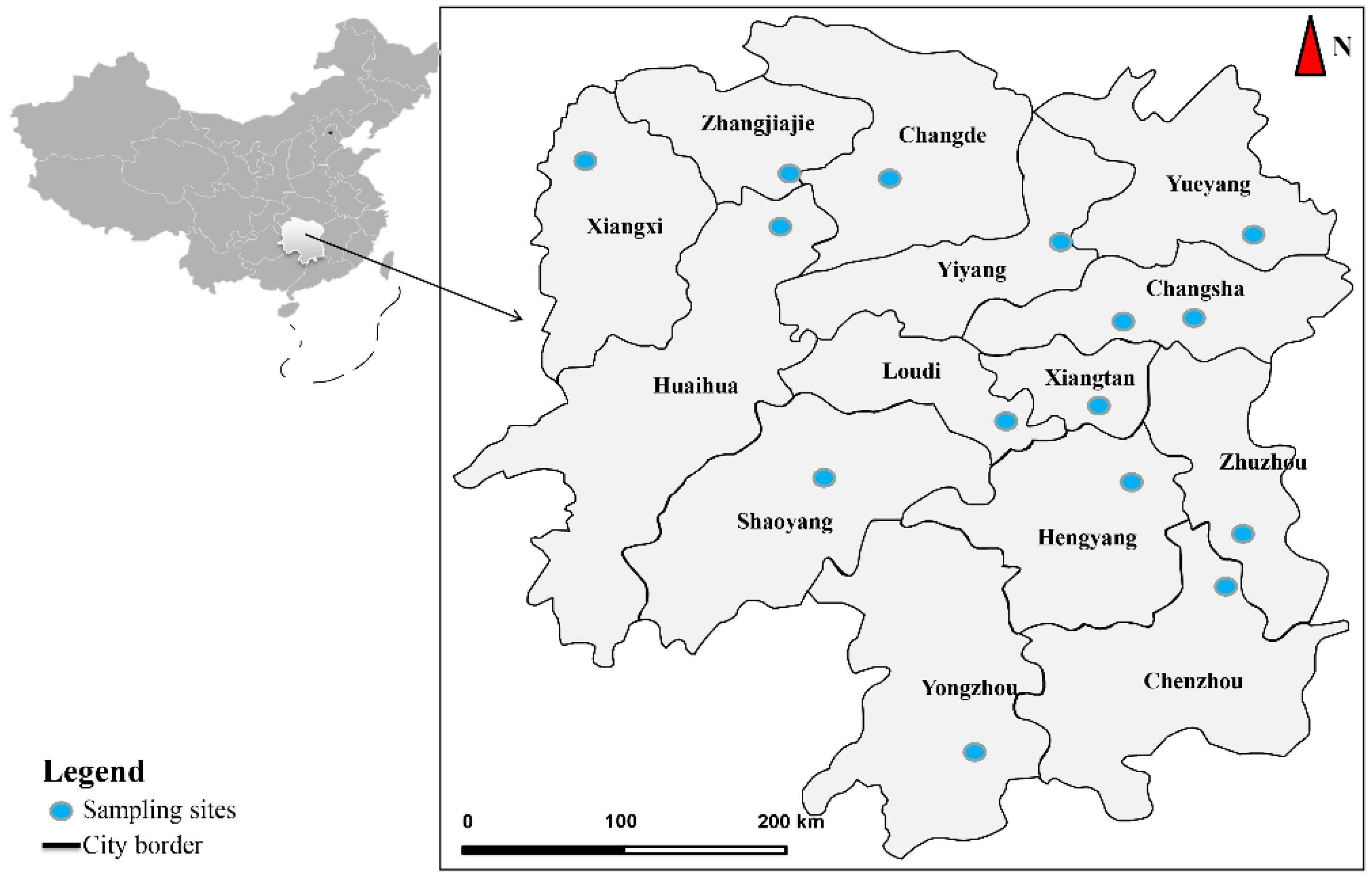
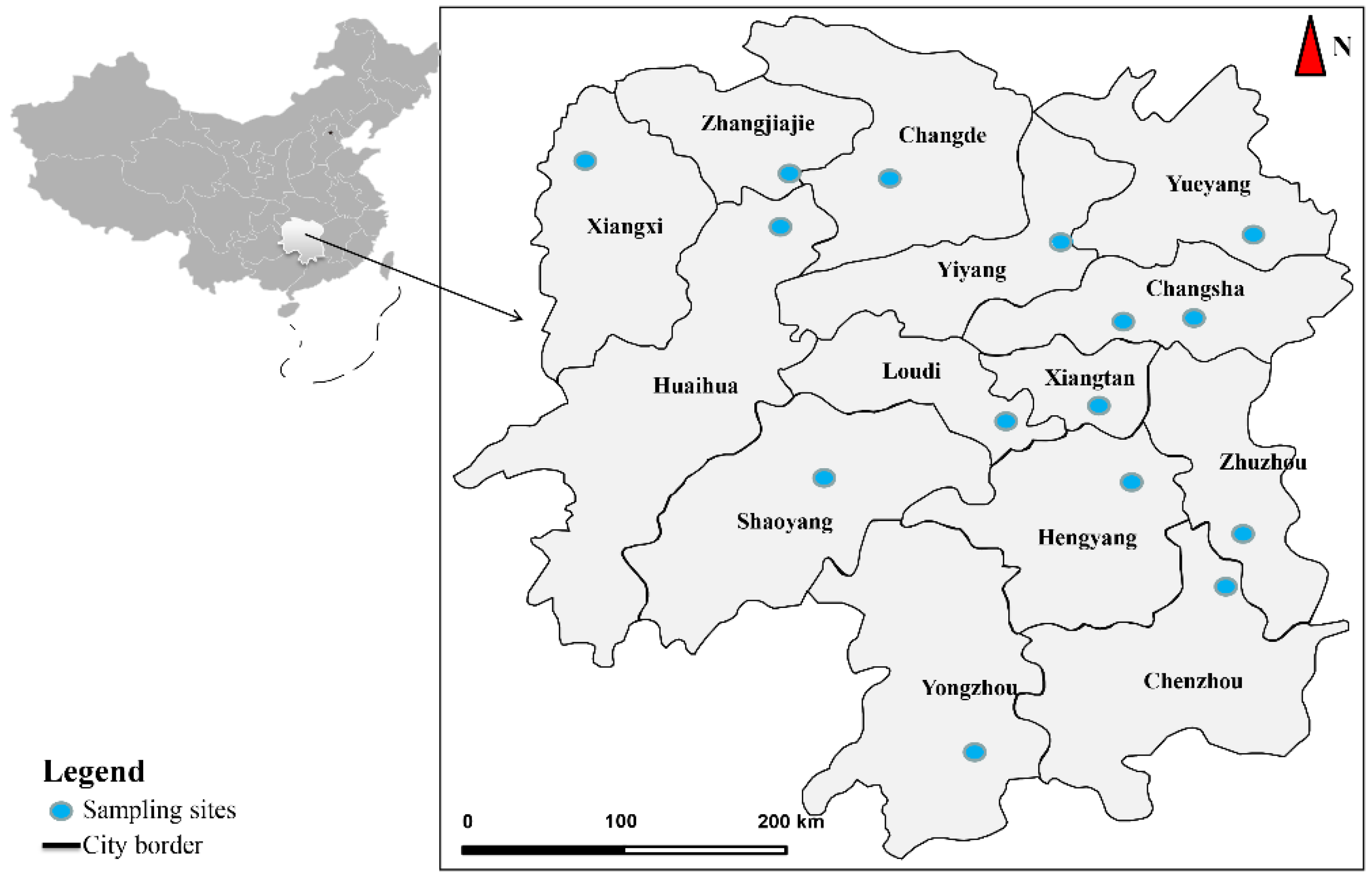
2.1. Sample Collection
2.2. Morphological Observations
2.3. DNA Extraction and Enzymatic Amplification
2.4. Sequence Analysis
2.5. Phylogenetic Analysis
3. Results
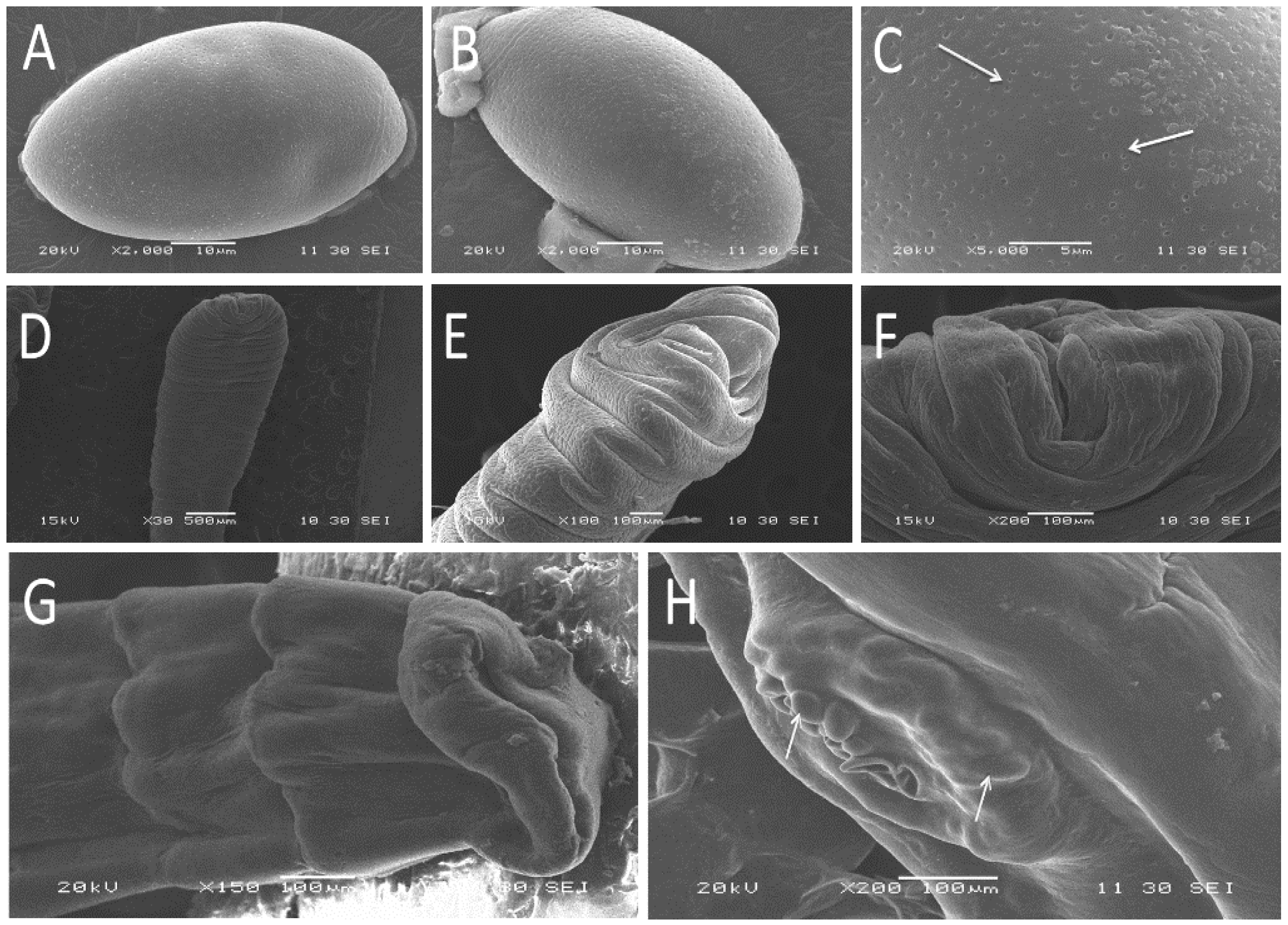
3.1. Morphological Characteristics
3.2. Genetic Characterisations of Spirometra Tapeworms
3.3. Phylogenetic Relationship of S. erinaceieuropaei
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Gene | Name | Sequence (5′–3′) | References |
|---|---|---|---|
| 18S | PL3F | ACCTGGTTGATCCTGCCAG | Barta et al., 1997 |
| PL3R | CTTCCGCTGGTTCACCTACGG | ||
| 28S | 28S-F | TGATAGGTTATTTAAACTGGC | This study |
| 28S-R | ACCCGACCCGTCTTGAAACA |
Appendix B
| Species | Country of Origin | Host | Sample Codes | Accession Number | References | |
|---|---|---|---|---|---|---|
| 18S | 28S | |||||
| Spirometra erinaceieuropaei | Yiyang City, Hunan Province, China | Zaocys dhumnades | HuN-YiY1 | MZ267595 | MZ293029 | This study |
| Z. dhumnades | HuN-YiY2 | MZ267596 | MZ293030 | This study | ||
| Elaphe carinata | HuN-YiY3 | MZ267597 | MZ293031 | This study | ||
| Changde City, Hunan Province, China | Z. dhumnades | HuN-CD1 | MZ267569 | MZ293003 | This study | |
| Z. dhumnades | HuN-CD2 | MZ267570 | MZ293004 | This study | ||
| E. carinata | HuN-CD3 | MZ267571 | MZ293005 | This study | ||
| Yongzhou City, Hunan Province, China | Z. dhumnades | HuN-YZ1 | MZ267598 | MZ293035 | This study | |
| Z. dhumnades | HuN-YZ2 | MZ267599 | MZ293036 | This study | ||
| Z. dhumnades | HuN-YZ3 | MZ267600 | MZ293037 | This study | ||
| Hengyang City, Hunan Province, China | Z. dhumnades | HuN-HY1 | MZ267583 | MZ293017 | This study | |
| Z. dhumnades | HuN-HY2 | MZ267584 | MZ293018 | This study | ||
| E. carinata | HuN-HY3 | MZ267585 | MZ293019 | This study | ||
| Xiangtan City, Hunan Province, China | Z. dhumnades | HuN-XT1 | MZ267589 | MZ293023 | This study | |
| Z. dhumnades | HuN-XT2 | MZ267590 | MZ293024 | This study | ||
| E. carinata | HuN-XT3 | MZ267591 | MZ293025 | This study | ||
| Shaoyang City, Hunan Province, China | Z. dhumnades | HuN-SY1 | MZ267586 | MZ293020 | This study | |
| Z. dhumnades | HuN-SY2 | MZ267587 | MZ293021 | This study | ||
| E. carinata | HuN-SY3 | MZ267588 | MZ293022 | This study | ||
| Zhuzhou City, Hunan Province, China | Z. dhumnades | HuN-ZZ1 | MZ267604 | MZ293041 | This study | |
| Z. dhumnades | HuN-ZZ2 | MZ267605 | MZ293042 | This study | ||
| E. taeniura | HuN-ZZ3 | MZ267606 | MZ293043 | This study | ||
| Changsha City, Hunan Province, China | Z. dhumnades | HuN-CS1 | MZ267572 | MZ293006 | This study | |
| Z. dhumnades | HuN-CS2 | MZ267573 | MZ293007 | This study | ||
| White Tiger | HuN- CS3 | MZ267607 | MZ292995 | This study | ||
| W. Tiger | HuN- CS4 | MZ267608 | MZ292996 | This study | ||
| Panthera tigris | HuN- CS5 | MZ267609 | MZ292997 | This study | ||
| P. tigris | HuN- CS6 | MZ267610 | MZ292998 | This study | ||
| Prionailurus bengalensis | HuN- CS7 | MZ267611 | MZ292999 | This study | ||
| P. bengalensis | HuN- CS8 | MZ267612 | MZ293000 | This study | ||
| Cat | HuN- CS9 | MZ267613 | MZ293001 | This study | ||
| Cat | HuN- CS10 | MZ267614 | MZ293000 | This study | ||
| Loudi City, Hunan Province, China | E. carinata | HuN-LD1 | MZ267580 | MZ293014 | This study | |
| E. carinata | HuN-LD2 | MZ267581 | MZ293015 | This study | ||
| E. carinata | HuN-LD3 | MZ267582 | MZ293016 | This study | ||
| Chenzhou City, Hunan Province, China | Z. dhumnades | HuN-CZ1 | MZ267574 | MZ293008 | This study | |
| Z. dhumnades | HuN-CZ2 | MZ267575 | MZ293009 | This study | ||
| Z. dhumnades | HuN-CZ3 | MZ267576 | MZ293010 | This study | ||
| Huaihua City, Hunan Province, China | Z. dhumnades | HuN-HH1 | MZ267577 | MZ293011 | This study | |
| Z. dhumnades | HuN-HH2 | MZ267578 | MZ293012 | This study | ||
| Z. dhumnades | HuN-HH3 | MZ267579 | MZ293013 | This study | ||
| Zhangjiajie City, Hunan Province, China | Z. dhumnades | HuN-ZZJ1 | MZ267601 | MZ293038 | This study | |
| Z. dhumnades | HuN-ZZJ2 | MZ267602 | MZ293039 | This study | ||
| Z. dhumnades | HuN-ZZJ3 | MZ267603 | MZ293040 | This study | ||
| Yueyang City, Hunan Province, China | Z. dhumnades | HuN-YuY1 | MZ267566 | MZ293032 | This study | |
| Z. dhumnades | HuN-YuY2 | MZ267567 | MZ293033 | This study | ||
| E. taeniura | HuN-YuY3 | MZ267568 | MZ293034 | This study | ||
| Xiangxi City, Hunan Province, China | Z. dhumnades | HuN-XX1 | MZ267592 | MZ293026 | This study | |
| Z. dhumnades | HuN-XX2 | MZ267593 | MZ293027 | This study | ||
| Z. dhumnades | HuN-XX3 | MZ267594 | MZ293028 | This study | ||
| Guilin City, Guangxi Province, China | Amphiesma stolatum | HQ228991 | HQ288992 | Lee et al., 2010 | ||
| Xiangtan City, Hunan Province, China | Rana nigromaculata | KX528089 | Zhang et al., 2017 | |||
| Australia | Canis familiaris | KY552801 | KY552835 | Kuchta et al., 2017 | ||
| Vietnam | Xenochrophis flavipunctatus | KY552802 | KY552836 | Kuchta et al., 2017 | ||
| Adenocephalus pacificus | Australia | Arctocephalus pusillus | KY552774 | KY552808 | Kuchta et al., 2017 | |
| USA | Callorhinus ursinus | KY552775 | KY552810 | Kuchta et al., 2017 | ||
| Australia | Neophoca cinerea | KY552776 | KY552809 | Kuchta et al., 2017 | ||
| Bothridium pithonis | Czech Republic | Chondropython viridis | KY552803 | KY552838 | Kuchta et al., 2017 | |
| Vietnam | Xenopeltis unicolor | KY552804 | KY552839 | Kuchta et al., 2017 | ||
| Dibothriocephalus nihonkaiensis | Japan | Homo sapiens | AB512013 | LC312467 | Yanagida et al., 2021 Yamasaki et al., 2021 | |
| Dibothriocephalus latus | Russia | Gymnocephalus cernuus | DQ925309 | DQ925326 | Brabec et al., 2016 | |
| Dibothriocephalus dendriticus | USA | Larus hyperboreus | KY552779 | KY552814 | Kuchta et al., 2017 | |
| United Kingdom | Coregonus lavaretus | KY552778 | KY552812 | Kuchta et al., 2017 | ||
| Dibothriocephalus ditremus | United Kingdom | Salvelinus alpinus | KY552780 | KY552813 | Kuchta et al., 2017 | |
| USA | Oncorhynchus tshawytscha | KY552787 | KY552815 | Kuchta et al., 2017 | ||
| Diphyllobothrium scoticum | Australia | Mirounga leonina | KY552777 | KY552811 | Kuchta et al., 2017 | |
| Diphyllobothrium dendriticum | USA | Larus hyperboreus | KY552779 | KY552814 | Kuchta et al., 2017 | |
| Diphyllobothrium schistochilos | Norway | Pusa hispida | KY552782 | KY552821 | Kuchta et al., 2017 | |
| Diphyllobothrium tetrapterum | USA | Callorhinus ursinus | KY552786 | KY552826 | Kuchta et al., 2017 | |
| Diphyllobothrium cordatum | USA | Erignathus barbatus | KY552788 | KY552882 | Kuchta et al., 2017 | |
| Diphyllobothrium lanceolatum | USA | Erignathus barbatus | KY552789 | KY552823 | Kuchta et al., 2017 | |
| Diphyllobothrium stemmacephalum | USA | Lagenorhynchus acutus | AF124459 | AF286943 | Kuchta et al., 2017 | |
| Diphyllobothrium balaenopterae | Japan | Homo sapiens | KY552792 | KY552824 | Kuchta et al., 2017 | |
| Duthiersia fimbriata | Ghana | Varanus exanthematicus | AF267290 | DQ925328 | Kodedova et al., 2001 Brabec et al., 2006 | |
| Duthiersia expansa | Vietnam | Varanus salvator | KY552806 | KY552840 | ||
| Ligula intestinalis | USA | Oncorhynchus tshawytscha | KY552783 | KY552818 | Kuchta et al., 2017 | |
| Ligula intestinalis | Czech Republic | Podiceps cristatus | KY552785 | KY552819 | Kuchta et al., 2017 | |
| Ligula pavlovskii | Ukraine | Neogobius fluviatilis | KY552784 | KY552820 | Kuchta et al., 2017 | |
| Probothriocephalus alaini | Atlantic Ocean | Xenodermichthys copei | KR780925 | KR780881 | Brabec et al., 2015 | |
| Pyramicocephalus phocarum | Norway | Myoxocephalus scorpius | KY552790 | KY552827 | Kuchta et al., 2017 | |
| Norway | Pollachius virens | KY552791 | KY552828 | Kuchta et al., 2017 | ||
| Schistocephalus solidus | Poland | Gasterosteus aculeatus | KY552797 | KY552832 | Kuchta et al., 2017 | |
| Norway | Gasterosteus aculeatus | KY552798 | KY552833 | Kuchta et al., 2017 | ||
| Schistocephalus pungitii | Germany | Pungitius pungitius | KY552799 | KY552834 | Kuchta et al., 2017 | |
| Haplobothrium globuliforme | Canada | Amia calva | AF124458 | AF286926 | Olson et al., 1999 Olson et al., 2001 | |
References
- Hughes, A.J.; Biggs, B.A. Parasitic worms of the central nervous system: An Australian perspective. Intern. Med. J. 2002, 32, 541–553. [Google Scholar] [CrossRef]
- Oda, F.H.; Borteiro, C.; da Graça, R.J.; Tavares, L.E.R.; Crampet, A.; Guerra, V.; Lima, F.S.; Bellay, S.; Karling, L.C.; Castro, O.; et al. Parasitism by larval tapeworms genus Spirometra in South American amphibians and reptiles: New records from Brazil and Uruguay, and a review of current knowledge in the region. Acta Trop. 2016, 164, 150–164. [Google Scholar] [CrossRef]
- Hong, D.; Xie, H.; Wan, H.; An, N.; Xu, C.; Zhang, J. Efficacy comparison between long-term high-dose praziquantel and surgical therapy for cerebral sparganosis: A multicenter retrospective cohort study. PLoS Negl. Trop. Dis. 2016, 12, e0006918. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Li, W.; Hua, L.; Gong, S.; Xiao, J.; Hou, F.; Ge, Y.; Yang, G.D. Spirometra (Pseudophyllidea, Diphyllobothriidae) severely infecting wild-caught snakes from food markets in Guangzhou and Shenzhen, Guangdong, China: Implications for public health. Sci. World J. 2014, 874014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Li, M.W.; Wang, Z.D.; Zhao, G.H.; Zhu, X.Q. Human sparganosis, a neglected food borne zoonosis. Lancet Infect. Dis. 2015, 15, 1226–1235. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, H.; Cui, J.; Jiang, P.; Lin, M.L.; Zhang, Y.L.; Liu, R.D.; Wang, Z.Q. The phylogenetic diversity of Spirometra erinaceieuropaei isolates from southwest China revealed by multi genes. Acta Trop. 2016, 156, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Kuchta, R.; Kołodziej-Sobocińska, M.; Brabec, J.; Młocicki, D.; Sałamatin, R.; Scholz, T. Sparganosis (Spirometra) in Europe in the Molecular Era. Clin. Infect. Dis. 2021, 72, 882–890. [Google Scholar] [CrossRef]
- Anantaphruti, M.T.; Nawa, Y.; Vanvanitchai, Y. Human sparganosis in Thailand: An overview. Acta Trop. 2011, 118, 171–176. [Google Scholar] [CrossRef]
- Li, M.W.; Song, H.Q.; Li, C.; Lin, H.Y.; Xie, W.T.; Lin, R.Q.; Zhu, X.Q. Sparganosis in mainland China. Int. J. Infect. Dis. 2011, 15, e154–e156. [Google Scholar] [CrossRef] [Green Version]
- Bennett, H.M.; Mok, H.P.; Gkrania-Klotsas, E.; Tsai, I.J.; Stanley, E.J.; Antoun, N.M.; Coghlan, A.; Harsha, B.; Berriman, M. The genome of the sparganosis tapeworm Spirometra erinaceieuropaei isolated from the biopsy of a migrating brain lesion. Genome Biol. 2014, 15, 510. [Google Scholar] [CrossRef]
- Zhang, X.; Hong, X.; Liu, S.N.; Jiang, P.; Zhao, S.C.; Sun, C.X.; Wang, Z.Q.; Cui, J. Large-scale survey of a neglected agent of sparganosis Spirometra erinaceieuropaei (Cestoda: Diphyllobothriidae) in wild frogs in China. PLoS Negl. Trop. Dis. 2020, 14, e0008019. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Tang, H.; Abuzeid, A.M.I.; Tan, L.; Wang, A.; Wan, X.; Zhang, H.J.; Liu, Y.S.; Li, G.Q. Protein phosphorylation networks in spargana of Spirometra erinaceieuropaei revealed by phosphoproteomic analysis. Parasit. Vectors 2020, 13, 248. [Google Scholar] [CrossRef]
- Eom, K.S.; Park, H.; Lee, D.; Choe, S.; Kim, K.H.; Jeon, H.K. Mitochondrial Genome Sequences of Spirometra erinaceieuropaei and S. decipiens (Cestoidea: Diphyllobothriidae). Korean J. Parasitol. 2015, 53, 455–463. [Google Scholar] [CrossRef]
- Jeon, H.K.; Park, H.; Lee, D.; Choe, S.; Kim, K.H.; Sohn, W.M.; Eom, S. Genetic Identification of Spirometra decipiens Plerocercoids in Terrestrial Snakes from Korea and China. Korean J. Parasitol. 2016, 54, 181–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamasaki, H.; Sanpool, O.; Rodpai, R.; Sadaow, L.; Laummaunwai, P.; Un, M.; Thanchomnang, T.; Laymanivong, S.; Aung, W.P.P.; Intapan, P.M.; et al. Spirometra species from Asia: Genetic diversity and taxonomic challenges. Parasitol. Int. 2021, 80, 102181. [Google Scholar] [CrossRef]
- Zhang, X.; Duan, J.Y.; Shi, Y.L.; Jiang, P.; Zeng, D.J.; Wang, Z.Q.; Cui, J. Comparative mitochondrial genomics among Spirometra (Cestoda: Diphyllobothriidae) and the molecular phylogeny of related tapeworms. Mol. Phylogenet. Evol. 2017, 117, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Brabec, J.; Kuchta, R.; Scholz, T. Paraphyly of the Pseudophyllidea (Platyhelminthes: Cestoda): Circumscription of monophyletic clades based on phylogenetic analysis of ribosomal RNA. Int. J. Parasitol. 2006, 36, 1535–1541. [Google Scholar] [CrossRef]
- Hernández-Orts, J.S.; Scholz, T.; Brabec, J.; Kuzmina, T.; Kuchta, R. High morphological plasticity and global geographical distribution of the Pacific broad tapeworm Adenocephalus pacificus (syn. Diphyllobothrium pacificum): Molecular and morphological survey. Acta Trop. 2015, 149, 168–178. [Google Scholar] [CrossRef]
- Tan, L.; Wang, A.B.; Zheng, S.Q.; Zhang, X.L.; Huang, C.J.; Liu, W. Molecular characterization and phylogenetic analysis of Taenia multiceps from China. Acta Parasitol. 2018, 63, 721–727. [Google Scholar] [CrossRef] [PubMed]
- Olson, P.D.; Caira, J.N. Evolution of the major lineages of tapeworms (Platyhelminthes: Cestoidea) inferred from 18S ribosomal DNA and elongation factor-1alpha. J. Parasitol. 1999, 85, 1134–1159. [Google Scholar] [CrossRef] [Green Version]
- Waeschenbach, A.; Brabec, J.; Scholz, T.; Littlewood, D.T.J.; Kuchta, R. The catholic taste of broad tapeworms-multiple routes to human infection. Int. J. Parasitol. 2017, 47, 831–843. [Google Scholar] [CrossRef]
- Avcioglu, H.; Yildirim, A.; Duzlu, O.; Inci, A.; Terim, K.A.K.; Balkaya, I. Prevalence and molecular characterization of bovine coenurosis from Eastern Anatolian region of Turkey. Vet. Parasitol. 2011, 176, 59–64. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, X. DAMBE7, New and Improved Tools for Data Analysis in Molecular Biology and Evolution. Mol. Biol. Evol. 2018, 35, 1550–1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burland, T.G. DNASTAR’s Lasergene sequence analysis software. Methods Mol. Biol. 2000, 132, 71–91. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5, a software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Duan, J.Y.; Wang, Z.Q.; Jiang, P.; Liu, R.D.; Cui, J. Using the small subunit of nuclear ribosomal DNA to reveal the phylogenetic position of the plerocercoid larvae of Spirometra tapeworms. Exp. Parasitol. 2017, 175, 1–7. [Google Scholar] [CrossRef]
- Hao, G.Y. Sequencing and Phylogenetic Analysis of Mitochondrial Cytb Gene and nad4 Gene of Cysticercus cellulosae Xichang lsolates. Prog. Vet. Med. 2015, 36, 59–63. [Google Scholar]
- Hao, G.Y.; He, X.Q.; Wang, R.K. Genetic Variation Analysis of Mitochondrial cox1 and nad4 Genes of Ascaridia galli. China Anim. Husb. Vet. Med. 2017, 44, 113–122. [Google Scholar]
- Okino, T.; Yamasaki, H.; Yamamoto, Y.; Fukuma, Y.; Kurebayashi, J.; Sanuki, F.; Moriya, T.; Ushirogawa, H.; Saito, M. A case of human breast sparganosis diagnosed as Spirometra Type I by molecular analysis in Japan. Parasitol. Int. 2021, 84, 102383. [Google Scholar] [CrossRef]


| Geographical Origins | Host | Location | Sample Codes |
|---|---|---|---|
| Yiyang City | |||
| Lanxi Town, Heshan District | Zaocys dhumnades | 112°46′ E, 28°59′ N | HuN-YiY1 |
| Z. dhumnades | 112°46′ E, 28°59′ N | HuN-YiY2 | |
| Elaphe carinata | 112°46′ E, 28°59′ N | HuN-YiY3 | |
| Changde City | |||
| Taizimiao Town, Hanshou County | Z. dhumnades | 111°96′ E, 28°77′ N | HuN-CD1 |
| Z. dhumnades | 111°96′ E, 28°77′ N | HuN-CD2 | |
| E. carinata | 111°96′ E, 28°77′ N | HuN-CD3 | |
| Yongzhou City | |||
| Taiping Town, Ningyuan County | Z. dhumnades | 112°13′ E, 25°67′ N | HuN-YZ1 |
| Z. dhumnades | 112°13′ E, 25°67′ N | HuN-YZ2 | |
| Z. dhumnades | 112°13′ E, 25°67′ N | HuN-YZ3 | |
| Hengyang City | |||
| Xuanzhou Town, Hengyang County | Z. dhumnades | 112°85′ E, 27°24′ N | HuN-HY1 |
| Z. dhumnades | 112°85′ E, 27°24′ N | HuN-HY2 | |
| E. carinata | 112°85′ E, 27°24′ N | HuN-HY3 | |
| Xiangtan City | |||
| Jinshi Country, Xiangtan County | Z. dhumnades | 112°75′ E, 27°59′ N | HuN-XT1 |
| Z. dhumnades | 112°75′ E, 27°59′ N | HuN-XT2 | |
| E. carinata | 112°75′ E, 27°59′ N | HuN-XT3 | |
| Shaoyang City | |||
| Shizhu Town, Dongkou County | Z. dhumnades | 110°73′ E, 27°25′ N | HuN-SY1 |
| Z. dhumnades | 110°73′ E, 27°25′ N | HuN-SY2 | |
| E. carinata | 110°73′ E, 27°25′ N | HuN-SY3 | |
| Zhuzhou City | |||
| Jieshou Town, Chaling County | Z. dhumnades | 113°43′ E, 26°61′N | HuN-ZZ1 |
| Z. dhumnades | 113°43′ E, 26°61′N | HuN-ZZ2 | |
| Elaphe taeniura | 113°43′ E, 26°61′N | HuN-ZZ3 | |
| Changsha City | |||
| Langli Town, Changsha County | Z. dhumnades | 113°13′ E, 28°19′ N | HuN-CS1 |
| Z. dhumnades | 113°13′ E, 28°19′ N | HuN-CS2 | |
| Changsha Ecological Zoo, Tianxin District | White Tiger | 113°01′ E, 28°04′ N | HuN-CS3 |
| W. Tiger | 113°01′ E, 28°04′ N | HuN-CS4 | |
| Panthera tigris | 113°01′ E, 28°04′ N | HuN-CS5 | |
| P. tigris | 113°01′ E, 28°04′ N | HuN-CS6 | |
| Prionailurus bengalensis | 113°01′ E, 28°04′ N | HuN-CS7 | |
| P.bengalensis | 113°01′ E, 28°04′ N | HuN-CS8 | |
| Cat | 113°01′ E, 28°04′ N | HuN-CS9 | |
| Cat | 113°01′ E, 28°04′ N | HuN-CS10 | |
| Loudi city | |||
| Suoshi Town, Shuangfeng County | E. carinata | 112°12′ E, 27°32′ N | HuN-LD1 |
| E. carinata | 112°12′ E, 27°32′ N | HuN-LD2 | |
| E. carinata | 112°12′ E, 27°32′ N | HuN-LD3 | |
| Chenzhou City | |||
| Longhai Town, Anren County | Z. dhumnades | 113°29′ E, 26°48′ N | HuN-CZ1 |
| Z. dhumnades | 113°29′ E, 26°48′ N | HuN-CZ2 | |
| Z. dhumnades | 113°29′ E, 26°48′ N | HuN-CZ3 | |
| Huaihua City | |||
| Qijiaping Town, Yuanling County | Z. dhumnades | 110°86′ E, 28°88′ N | HuN-HH1 |
| Z. dhumnades | 110°86′ E, 28°88′ N | HuN-HH2 | |
| Z. dhumnades | 110°86′ E, 28°88′ N | HuN-HH3 | |
| Zhangjiajie City | |||
| Dongxi Coutry, Cili County | Z. dhumnades | 110°83′ E, 29°14′ N | HuN-ZZJ1 |
| Z. dhumnades | 110°83′ E, 29°14′ N | HuN-ZZJ2 | |
| Z. dhumnades | 110°83′ E, 29°14′ N | HuN-ZZJ3 | |
| Yueyang City | |||
| Tongshi Town, Pingjiang County | Z. dhumnades | 113°72′ E, 28°75′ N | HuN-YuY1 |
| Z. dhumnades | 113°72′ E, 28°75′ N | HuN-YuY2 | |
| E. taeniura | 113°72′ E, 28°75′ N | HuN-YuY3 | |
| Xiangxi City | |||
| Xichehe Town, Longshan County | Z. dhumnades | 109°54′ E, 29°09′ N | HuN-XX1 |
| Z. dhumnades | 109°54′ E, 29°09′ N | HuN-XX2 | |
| Z. dhumnades | 109°54′ E, 29°09′ N | HuN-XX3 |
| N | S | H | π | Hd | K | |
|---|---|---|---|---|---|---|
| 18S | 49 | 18 | 8 | 0.00062 | 0.392 | 1.244 |
| 28S | 49 | 2 | 3 | 0.00028 | 0.275 | 0.281 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.-Y.; Gong, T.-F.; He, J.-L.; Li, F.; Li, W.-C.; Xie, L.-X.; Xie, X.-R.; Liu, Y.-S.; Zhou, Y.-F.; Liu, W. Molecular Characterization and Phylogenetic Analysis of Spirometra Tapeworms from Snakes in Hunan Province. Vet. Sci. 2022, 9, 62. https://doi.org/10.3390/vetsci9020062
Chen S-Y, Gong T-F, He J-L, Li F, Li W-C, Xie L-X, Xie X-R, Liu Y-S, Zhou Y-F, Liu W. Molecular Characterization and Phylogenetic Analysis of Spirometra Tapeworms from Snakes in Hunan Province. Veterinary Sciences. 2022; 9(2):62. https://doi.org/10.3390/vetsci9020062
Chicago/Turabian StyleChen, Shu-Yu, Teng-Fang Gong, Jun-Lin He, Fen Li, Wen-Chao Li, Li-Xing Xie, Xin-Rui Xie, Yi-Song Liu, Ying-Fang Zhou, and Wei Liu. 2022. "Molecular Characterization and Phylogenetic Analysis of Spirometra Tapeworms from Snakes in Hunan Province" Veterinary Sciences 9, no. 2: 62. https://doi.org/10.3390/vetsci9020062
APA StyleChen, S.-Y., Gong, T.-F., He, J.-L., Li, F., Li, W.-C., Xie, L.-X., Xie, X.-R., Liu, Y.-S., Zhou, Y.-F., & Liu, W. (2022). Molecular Characterization and Phylogenetic Analysis of Spirometra Tapeworms from Snakes in Hunan Province. Veterinary Sciences, 9(2), 62. https://doi.org/10.3390/vetsci9020062