

Identification of Contracaecum rudolphii (Nematoda: Anisakidae) in Great Cormorants Phalacrocorax carbo sinensis (Blumenbach, 1978) from Southern Italy
 , , , , , and
, , , , , and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
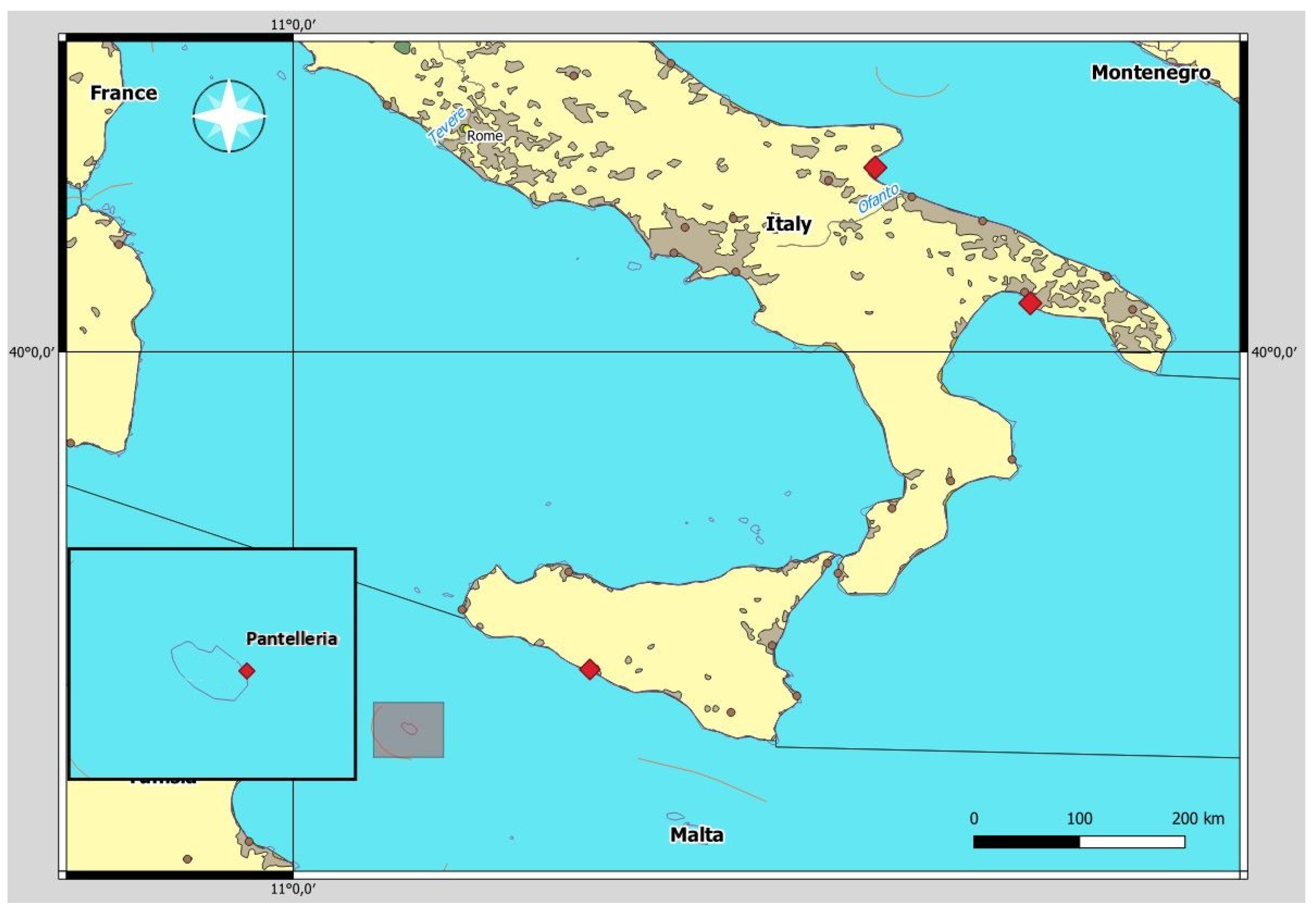
2.1. Samples Collection
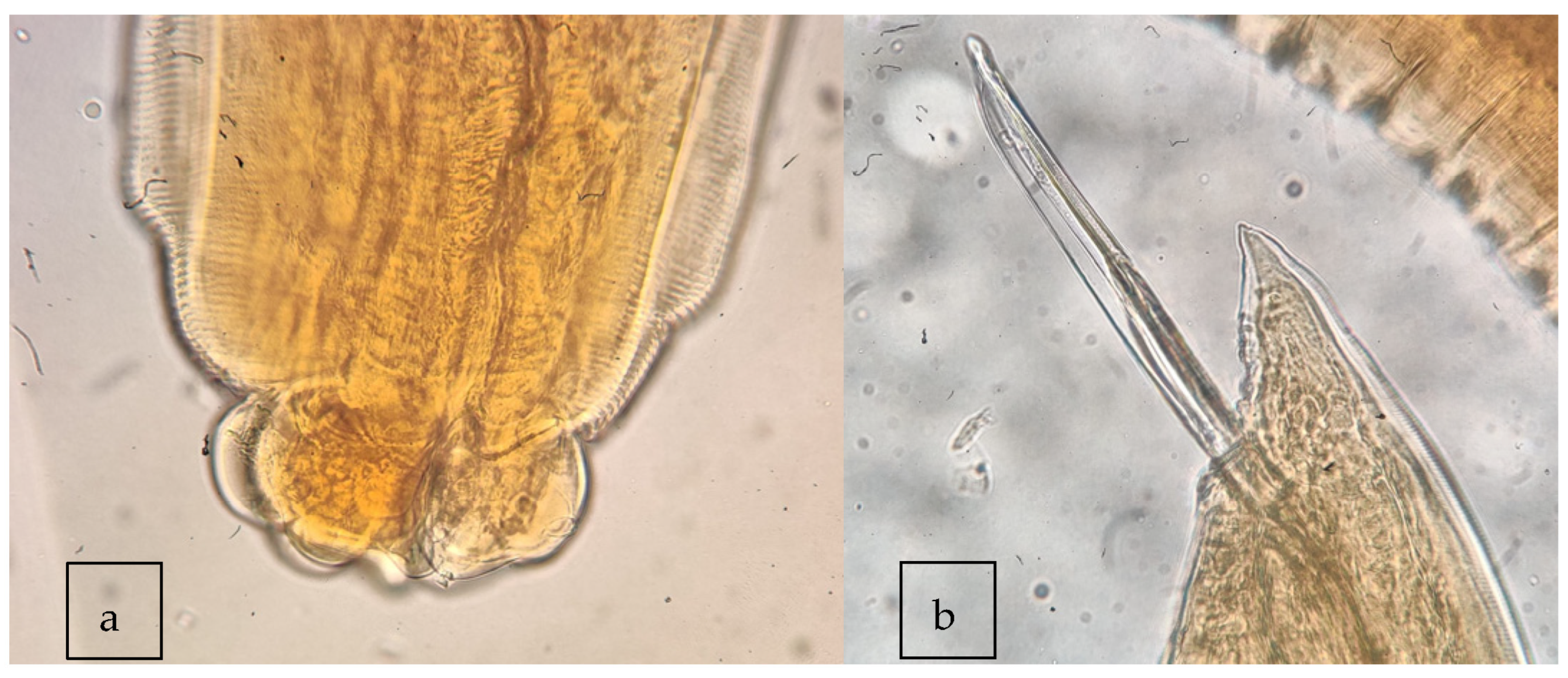
2.2. Visual Inspection and Morphological Analysis
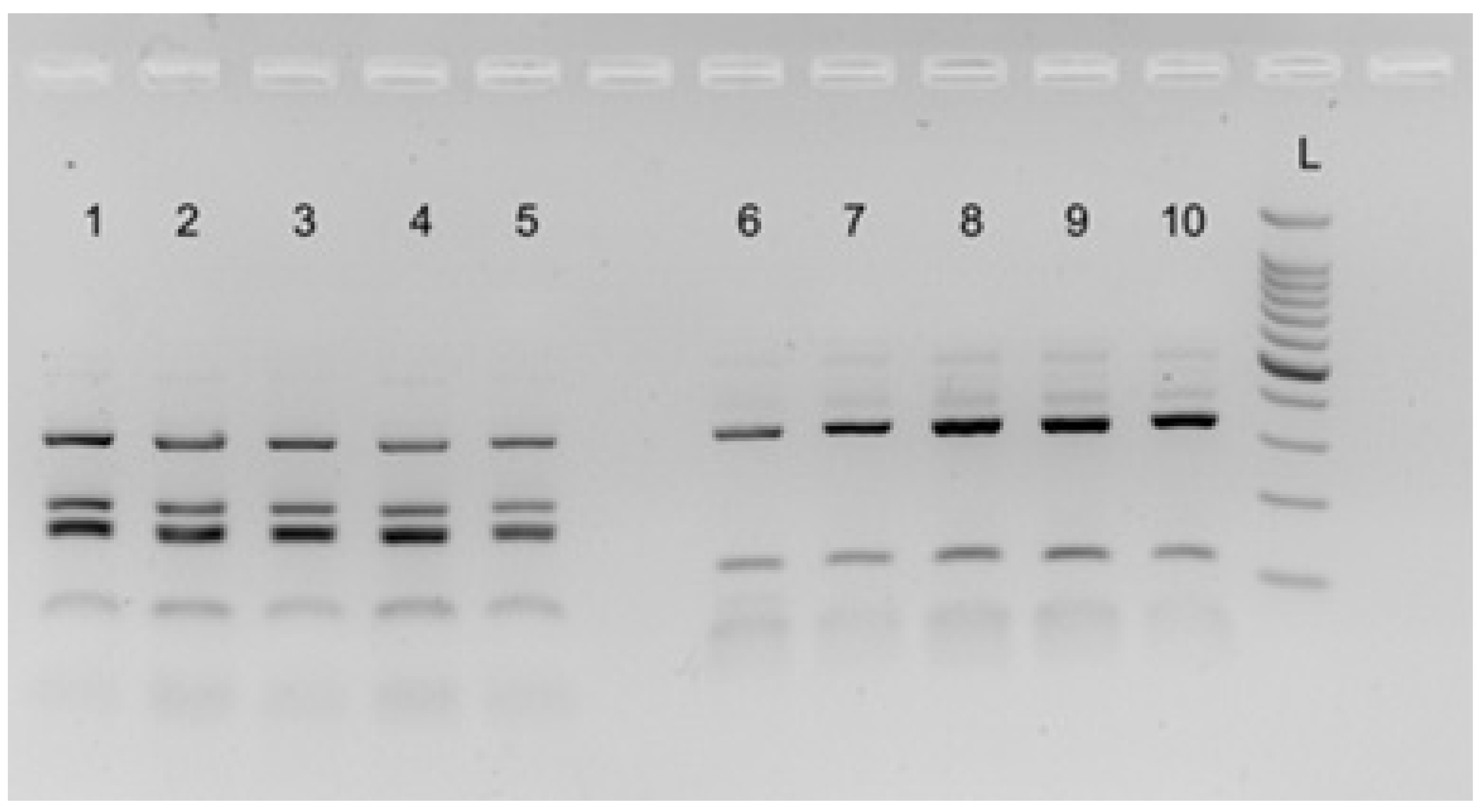
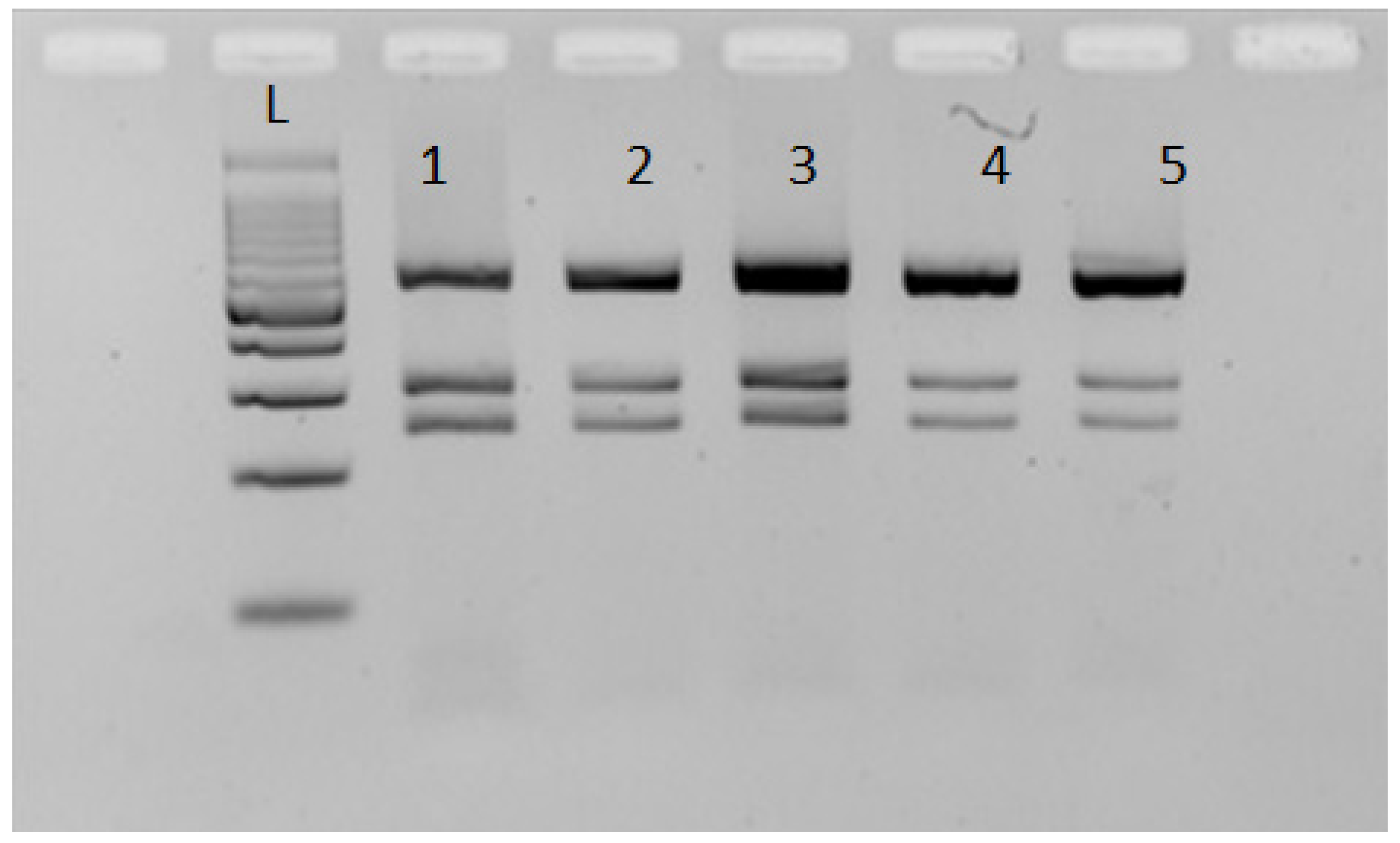
2.3. Molecular Analysis, Data Collection, and Analysis
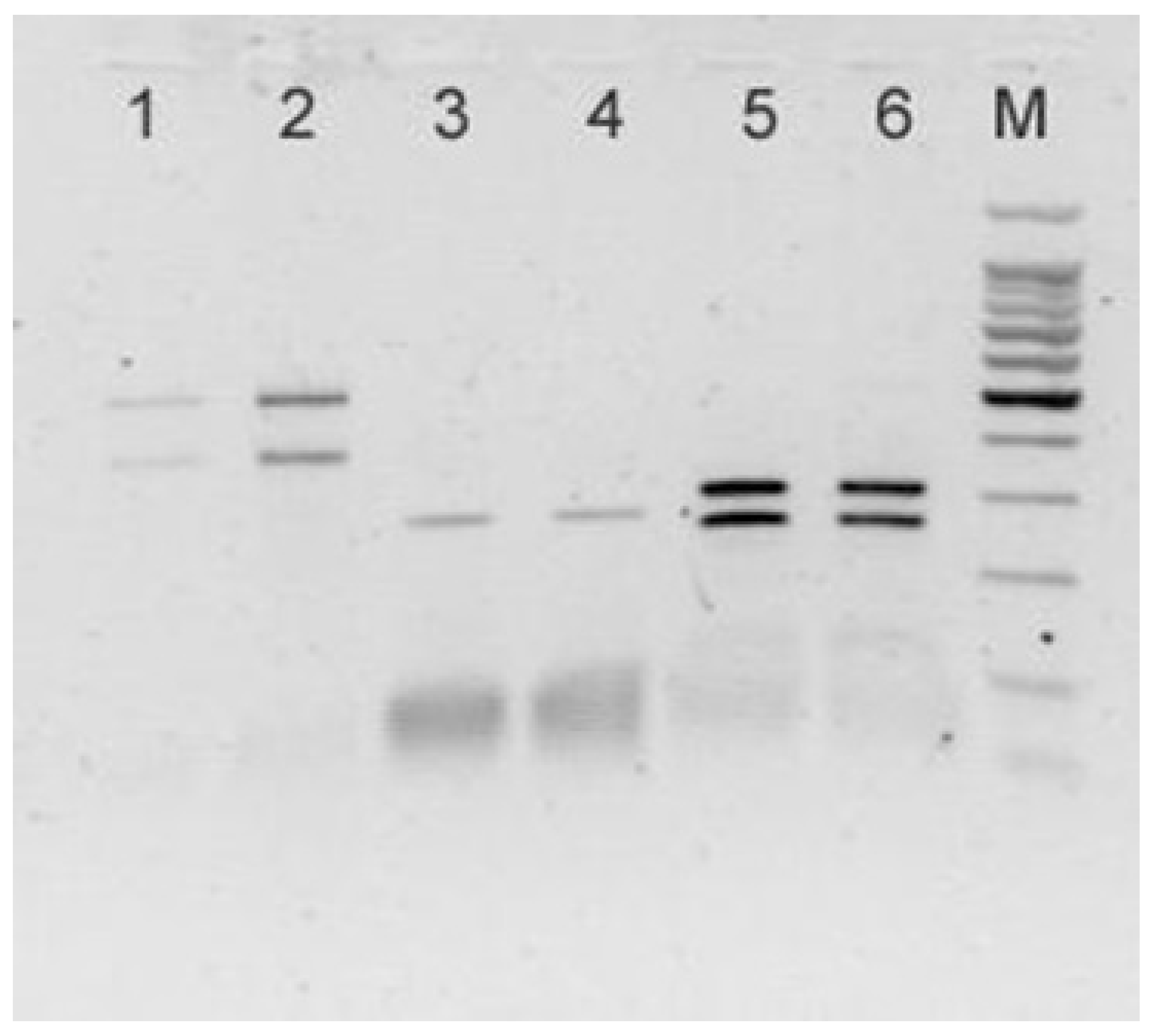
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cammilleri, G.; Pulvirenti, A.; Costa, A.; Graci, S.; Collura, R.; Buscemi, M.D.; Sciortino, S.; Badaco, V.V.; Vazzana, M.; Brunone, M.; et al. Seasonal Trend of Anisakidae Infestation in South Mediterranean Bluefish. Nat. Prod. Res. 2019, 34, 158–161. [Google Scholar] [CrossRef]
- Nalbone, L.; Panebianco, F.; Cammilleri, G.; Ferrantelli, V.; Giarratana, F. Anisakicidal Effects of R (+) Limonene: An Alternative to Freezing Treatment in the Industrial Anchovy Marinating Process. Foods 2022, 11, 1121. [Google Scholar] [CrossRef]
- Mattiucci, S.; Nascetti, G. Molecular Systematics, Phylogeny and Ecology of Anisakid Nematodes of the Genus Anisakis Dujardin, 1845: An Update. Parasite 2006, 13, 99–113. [Google Scholar] [CrossRef] [PubMed]
- Mattiucci, S.; Sbaraglia, G.L.; Palomba, M.; Filippi, S.; Paoletti, M.; Cipriani, P.; Nascetti, G. Genetic Identification and Insights into the Ecology of Contracaecum Rudolphii A and C. Rudolphii B (Nematoda: Anisakidae) from Cormorants and Fish of Aquatic Ecosystems of Central Italy. Parasitol. Res. 2020, 119, 1243–1257. [Google Scholar] [CrossRef]
- Moravec, F. Experimental Studies on the Development of Contracaecum Rudolphii (Nematoda: Anisakidae) in Copepod and Fish Paratenic Hosts. Folia Parasitol. 2009, 56, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Amor, N.; Farjallah, S.; Piras, M.C.; Burreddu, C.; Garippa, G.; Merella, P. New Insights into the Coexistence of Contracaecum rudolphii A and Contracaecum rudolphii B (Nematoda: Anisakidae) in Phalacrocorax carbo sinensis from Sardinia: Genetic Variability and Phylogenetic Analysis. Parasitology 2020, 147, 1538–1551. [Google Scholar] [CrossRef]
- Farjallah, S.; Merella, P.; Ingrosso, S.; Rotta, A.; Slimane, B.B.; Garippa, G.; Said, K.; Busi, M. Molecular Evidence for the Occurrence of Contracaecum rudolphii A (Nematoda: Anisakidae) in Shag Phalacrocorax aristotelis (Linnaeus) (Aves: Phalacrocoracidae) from Sardinia (Western Mediterranean Sea). Parasitol. Int. 2008, 57, 437–440. [Google Scholar] [CrossRef] [PubMed]
- Bregnballe, T.; Engström, H.; Knief, W.; Eerden, M.; van Rijn, S.; Kieckbusch, J.J.; Eskildsen, J. Development of the Breeding Population of Great Cormorants Phalacrocorax carbo sinensis in The Netherlands, Germany, Denmark and Sweden during the 1990s. Vogelwelt 2003, 124, 15–26. [Google Scholar]
- Baccetti, N.; Capizzi, D.; Corbi, F.; Massa, B.; Nissardi, S.; Spano, G.; Sposimo, P. Breeding Shearwaters on Italian Islands: Population Size, Island Selection and Coexistence with Their Main Alien Predator, the Black Rat. Riv. Ital. Orn. 2009, 78, 83–100. [Google Scholar]
- Bregnballe, T.; Lynch, J.; Parz-Gollner, R.; Marion, L.; Volponi, S.; Paquet, J.-Y.; Carss, D.; Eerden, M. Breeding Numbers of Great Cormorants Phalacrocorax carbo in the Western Palearctic, 2012–2013; Aarhus University, DCE–Danish Centre for Environment and Energy: Roskilde, Denmark, 2014; ISBN 978-87-7156-069-5. [Google Scholar]
- Szostakowska, B.; Fagerholm, H.-P. Coexistence and Genetic Variability of Contracaecum rudolphii A and Contracaecum rudolphii B (Nematoda: Anisakidae) in Cormorants, Phalacrocorax carbo sinensis, in the Baltic Region. J. Parasitol. 2012, 98, 472–478. [Google Scholar] [CrossRef]
- Carmeno, C.; Rusconi, A.; Castelli, M.; Prati, P.; Bragoni, R.; Santoro, A.; Postiglione, U.; Sassera, D.; Olivieri, E. Molecular Identification of Contracaecum Rudolphii A and B (Nematoda: Anisakidae) from Cormorants Collected in a Freshwater Ecosystem of the Pre-Alpine Area in Northern Italy. Vet. Parasitol. Reg. Stud. Rep. 2022, 27, 100674. [Google Scholar] [CrossRef] [PubMed]
- Culurgioni, J.; Sabatini, A.; De Murtas, R.; Mattiucci, S.; Figus, V. Helminth Parasites of Fish and Shellfish from the Santa Gilla Lagoon in Southern Sardinia, Italy. J. Helminthol. 2014, 88, 489–498. [Google Scholar] [CrossRef] [PubMed]
- D’Amelio, S.; Barros, N.B.; Ingrosso, S.; Fauquier, D.A.; Russo, R.; Paggi, L. Genetic Characterization of Members of the Genus Contracaecum (Nematoda: Anisakidae) from Fish-Eating Birds from West-Central Florida, USA, with Evidence of New Species. Parasitology 2007, 134, 1041–1051. [Google Scholar] [CrossRef]
- Barus, V.; Sergeeva, T.P.; Sonin, M.D.; Ryzhikov, K.M. Helminths of Fish-Eating Birds of the Palaearctic Region: Nematoda, 1978 ed.; Springer: Berlin/Heidelberg, Germany, 1978; Volume 1, ISBN 978-94-009-9974-9. [Google Scholar]
- Fagerholm, H.P. Systematic Implications of Male Caudal Morphology in Ascaridoid Nematode Parasites. Syst. Parasitol. 1991, 19, 215–229. [Google Scholar] [CrossRef]
- Rózsa, L.; Reiczigel, J.; Majoros, G. Quantifying Parasites in Samples of Hosts. J. Parasitol. 2000, 86, 228–232. [Google Scholar] [CrossRef]
- Torres, P.; Ortega, J.; Schlatter, R. Nematode Parasites of the Digestive Tract in Neotropic Cormorant Chicks (Phalacrocorax Brasilianus) from the River Cruces Ramsar Site in Southern Chile. Parasitol. Res. 2005, 97, 103–107. [Google Scholar] [CrossRef]
- Szostakowska, B.; Fagerholm, H.-P. Molecular Identification of Two Strains of Third-Stage Larvae of Contracaecum Rudolphii Sensu Lato (Nematoda: Anisakidae) From Fish in Poland. J. Parasitol. 2007, 93, 961–964. [Google Scholar] [CrossRef] [PubMed]
- Baruš, V.; Nagasawa, K.; Tenora, F.; Prokeš, M. The Head End Morphology of Contracaecum Rudolphii with Remarks on C. Himeu and C. Umiu (Nematoda, Anisakidae). Acta Univ. Agric. Silvic. Mendel. Brun. 2000, 48, 69–76. [Google Scholar]
- Kanarek, G.; Bohdanowicz, J. Larval Contracaecum sp. (Nematoda: Anisakidae) in the Great Cormorant [Phalacrocorax carbo (L., 1758)] from North-Eastern Poland: A Morphological and Morphometric Analysis. Vet. Parasitol. 2009, 166, 90–97. [Google Scholar] [CrossRef]
- Li, A.-X.; D’Amelio, S.; Paggi, L.; He, F.; Gasser, R.; Lun, Z.-R.; Abollo, E.; Turchetto, M.; Zhu, X.-Q. Genetic Evidence for the Existence of Sibling Species within Contracaecum rudolphii (Hartwich, 1964) and the Validity of Contracaecum septentrionale (Kreis, 1955) (Nematoda: Anisakidae). Parasitol. Res. 2005, 96, 361–366. [Google Scholar] [CrossRef]
- Gagliardi, A.; Gandolla, S.; Lardelli, R.; Tosi, G. Nuove colonie di cormorano (Phalacrocorax carbo) in Lombardia: Dinamica di insediamento e interazioni con l’ittiofauna. (New colonies of cormorants in Lombardy region: Settlement dynamics and interactions with fish species). Alula 2009, 16, 452–454. [Google Scholar]
- Volponi, S. Progetto REDCAFE: Un Approccio Pan-Europeo per la Risoluzione del Conflitto Tra Cormorani e Pescatori; Istituto Nazionale per la Fauna Selvatica: Ozzano Emilia, Italy, 2004. [Google Scholar]
- Molnár, K.; Székely, C.; Baska, F.; Müller, T.; Zuo, S.; Kania, P.W.; Nowak, B.; Buchmann, K. Differential Survival of 3rd Stage Larvae of Contracaecum rudolphii Type B Infecting Common Bream (Abramis brama) and Common Carp (Cyprinus carpio). Parasitol Res 2019, 118, 2811–2817. [Google Scholar] [CrossRef] [PubMed]
- Keller, T. The food of Cormorants (Phalacrocorax carbo sinensis) in Bavaria. J. Ornithol. 1998, 139, 389–400. [Google Scholar] [CrossRef]
- s000507 Avifauna del Lago Salso. Available online: https://lifelagosalso.it/public/documenti/22DOC-769.pdf (accessed on 15 March 2022).
- Ferrantelli, V.; Costa, A.; Graci, S.; Buscemi, M.D.; Giangrosso, G.; Porcarello, C.; Palumbo, S.; Cammilleri, G. Anisakid Nematodes as Possible Markers to Trace Fish Products. Ital. J. Food Saf. 2015, 4, 4090. [Google Scholar] [CrossRef]
- Goffredo, E.; Azzarito, L.; Di Taranto, P.; Mancini, M.E.; Normanno, G.; Didonna, A.; Faleo, S.; Occhiochiuso, G.; D’Attoli, L.; Pedarra, C.; et al. Prevalence of Anisakid Parasites in Fish Collected from Apulia Region (Italy) and Quantification of Nematode Larvae in Flesh. Int. J. Food Microbiol. 2019, 292, 159–170. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Area | N Birds | N. Contracaecum rudolphii (s.l.) | Location (N) | N Larvae | N Adults |
|---|---|---|---|---|---|
| Cattolica Eraclea (Platani River) | 1 | 92 | Stomach (92) | 54 | 38 |
| Pantelleria | 1 | 30 | Stomach (30) | - | 30 |
| Leporano Bay | 1 | 50 | Stomach (35), Intestine (15) | - | 50 |
| Salso Lake | 1 | 9 | Stomach (9) | - | 9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cammilleri, G.; D’Amelio, S.; Ferrantelli, V.; Costa, A.; Buscemi, M.D.; Castello, A.; Bacchi, E.; Goffredo, E.; Mancini, M.E.; Cavallero, S. Identification of Contracaecum rudolphii (Nematoda: Anisakidae) in Great Cormorants Phalacrocorax carbo sinensis (Blumenbach, 1978) from Southern Italy. Vet. Sci. 2023, 10, 194. https://doi.org/10.3390/vetsci10030194
Cammilleri G, D’Amelio S, Ferrantelli V, Costa A, Buscemi MD, Castello A, Bacchi E, Goffredo E, Mancini ME, Cavallero S. Identification of Contracaecum rudolphii (Nematoda: Anisakidae) in Great Cormorants Phalacrocorax carbo sinensis (Blumenbach, 1978) from Southern Italy. Veterinary Sciences. 2023; 10(3):194. https://doi.org/10.3390/vetsci10030194
Chicago/Turabian StyleCammilleri, Gaetano, Stefano D’Amelio, Vincenzo Ferrantelli, Antonella Costa, Maria Drussilla Buscemi, Annamaria Castello, Emanuela Bacchi, Elisa Goffredo, Maria Emanuela Mancini, and Serena Cavallero. 2023. "Identification of Contracaecum rudolphii (Nematoda: Anisakidae) in Great Cormorants Phalacrocorax carbo sinensis (Blumenbach, 1978) from Southern Italy" Veterinary Sciences 10, no. 3: 194. https://doi.org/10.3390/vetsci10030194
APA StyleCammilleri, G., D’Amelio, S., Ferrantelli, V., Costa, A., Buscemi, M. D., Castello, A., Bacchi, E., Goffredo, E., Mancini, M. E., & Cavallero, S. (2023). Identification of Contracaecum rudolphii (Nematoda: Anisakidae) in Great Cormorants Phalacrocorax carbo sinensis (Blumenbach, 1978) from Southern Italy. Veterinary Sciences, 10(3), 194. https://doi.org/10.3390/vetsci10030194