
Analysis of the B-Type Natriuretic Peptide and the Aminoterminal-Pro-B-Type Natriuretic Peptide in Different Parrot, Raptor and Owl Species

Abstract
:1. Introduction
2. Materials and Methods
2.1. Birds, Samples and Ethical Statement
2.2. RNA Isolation and PCR
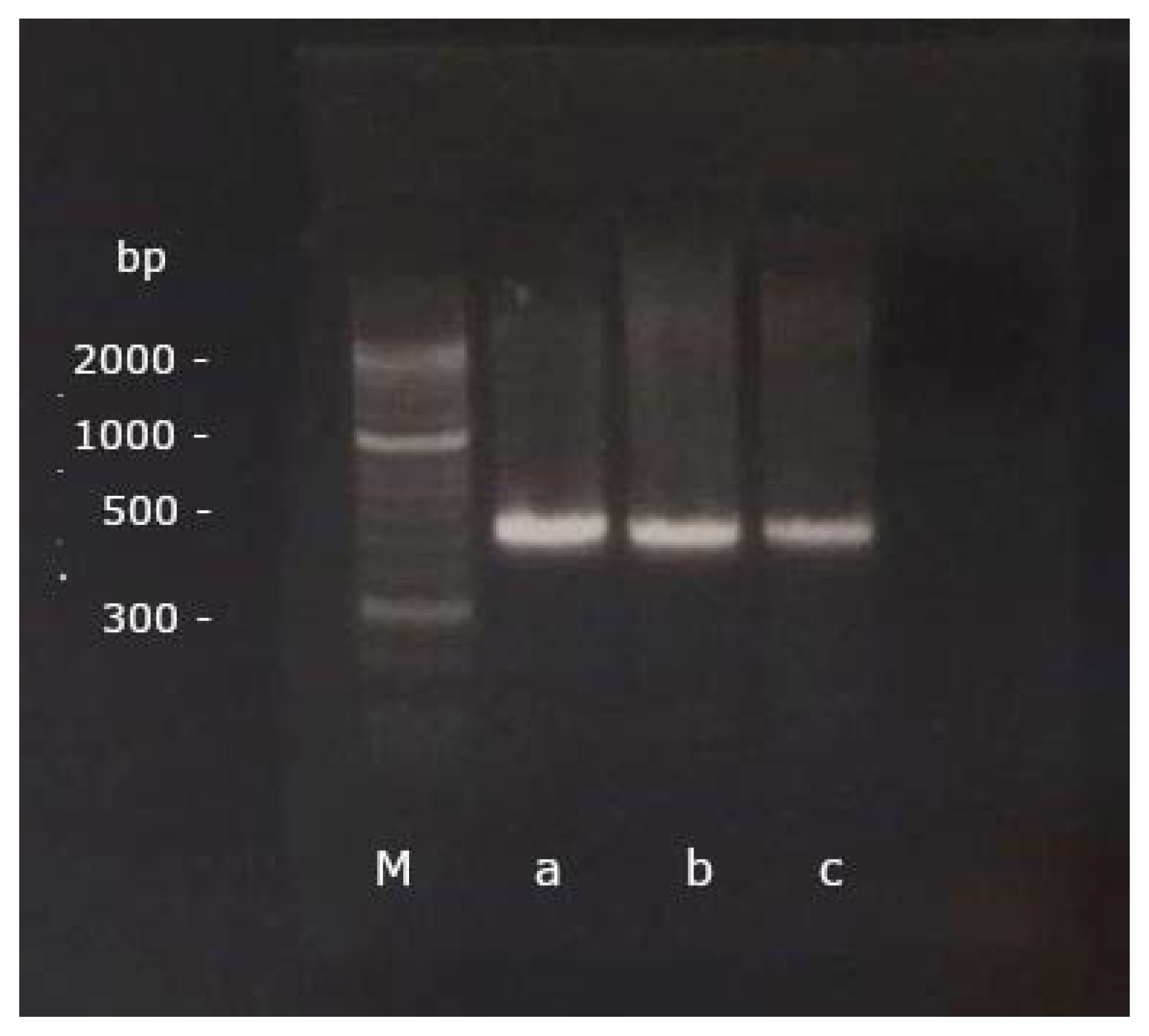
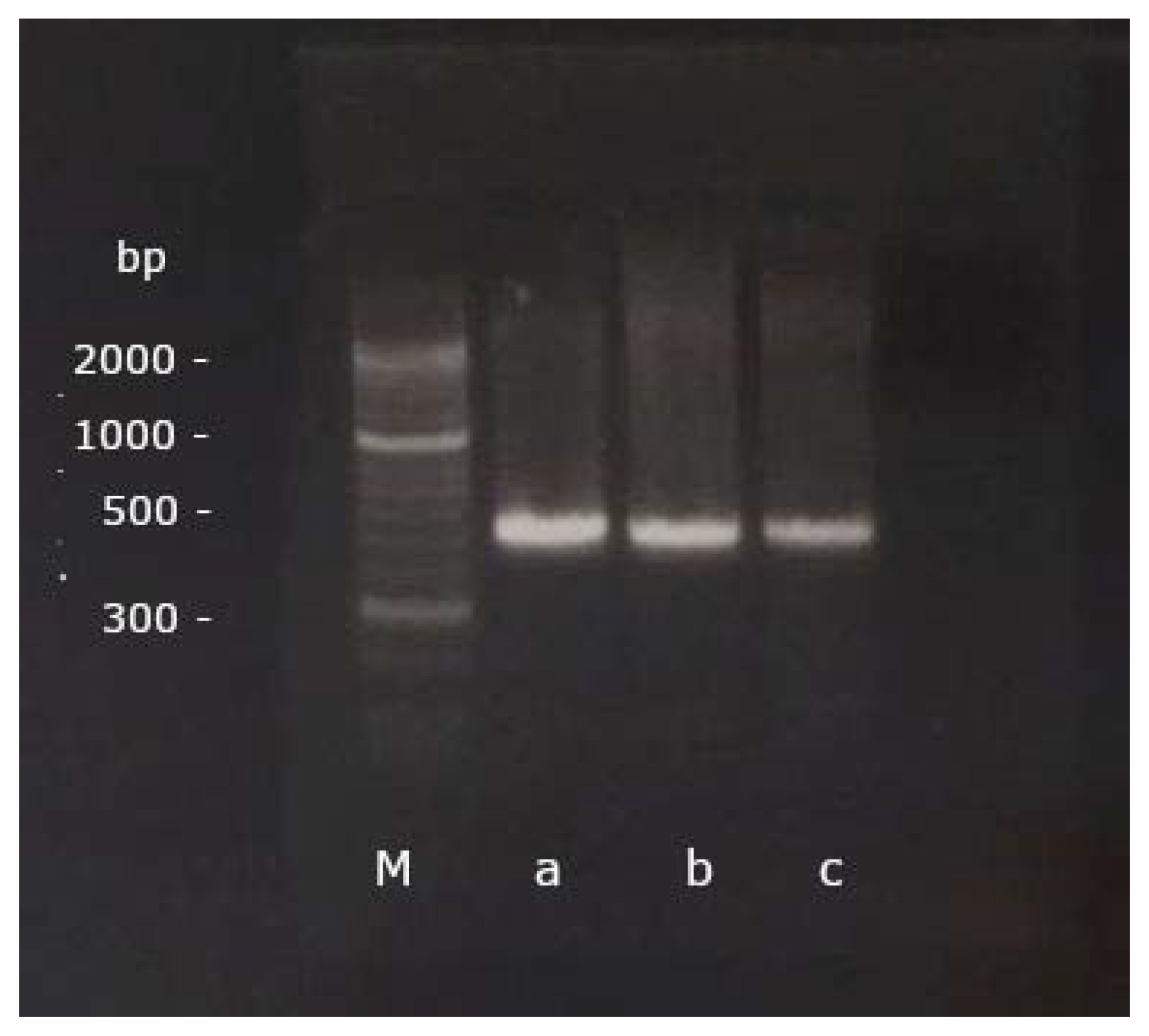
2.3. Gel Electrophoresis and Sequencing
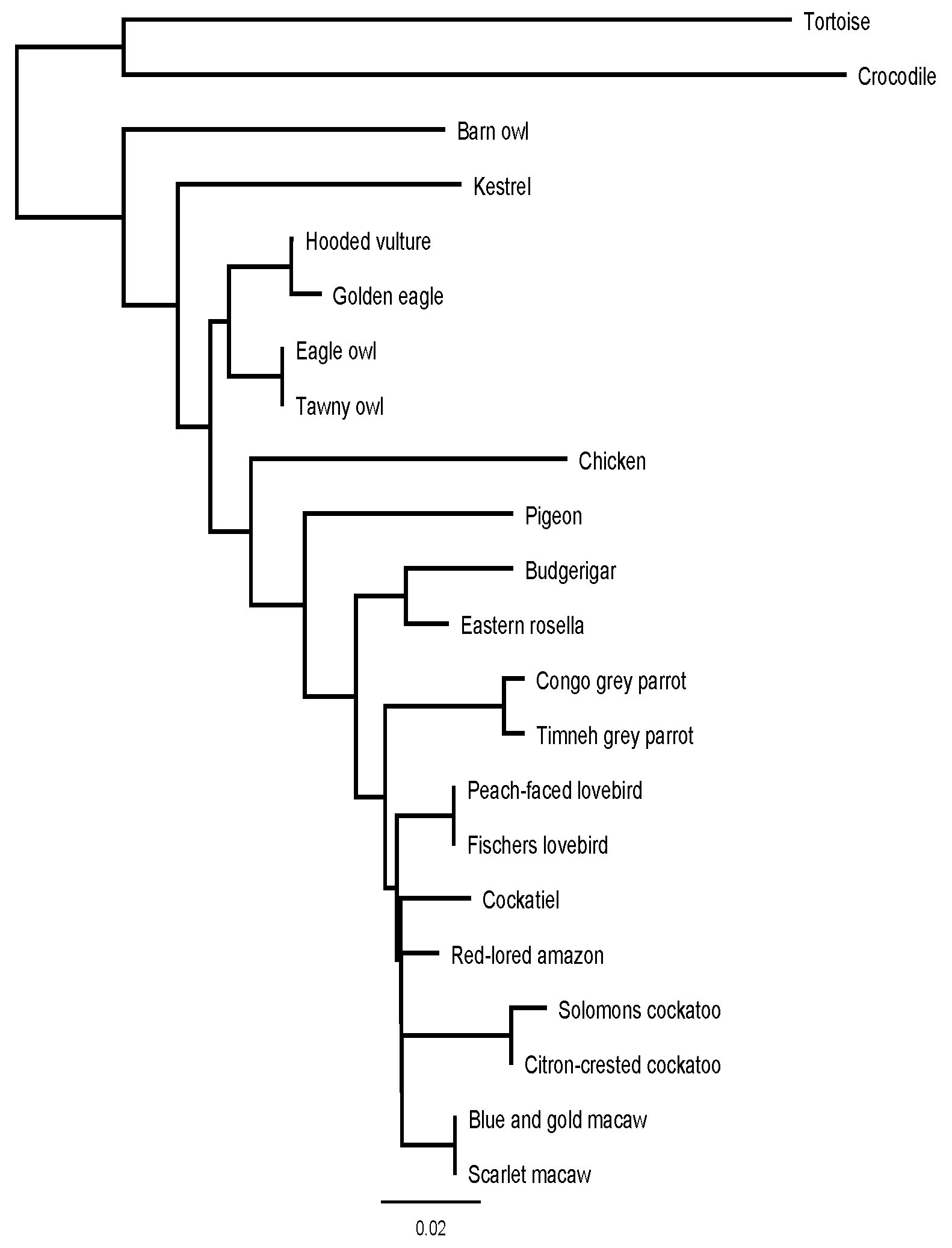
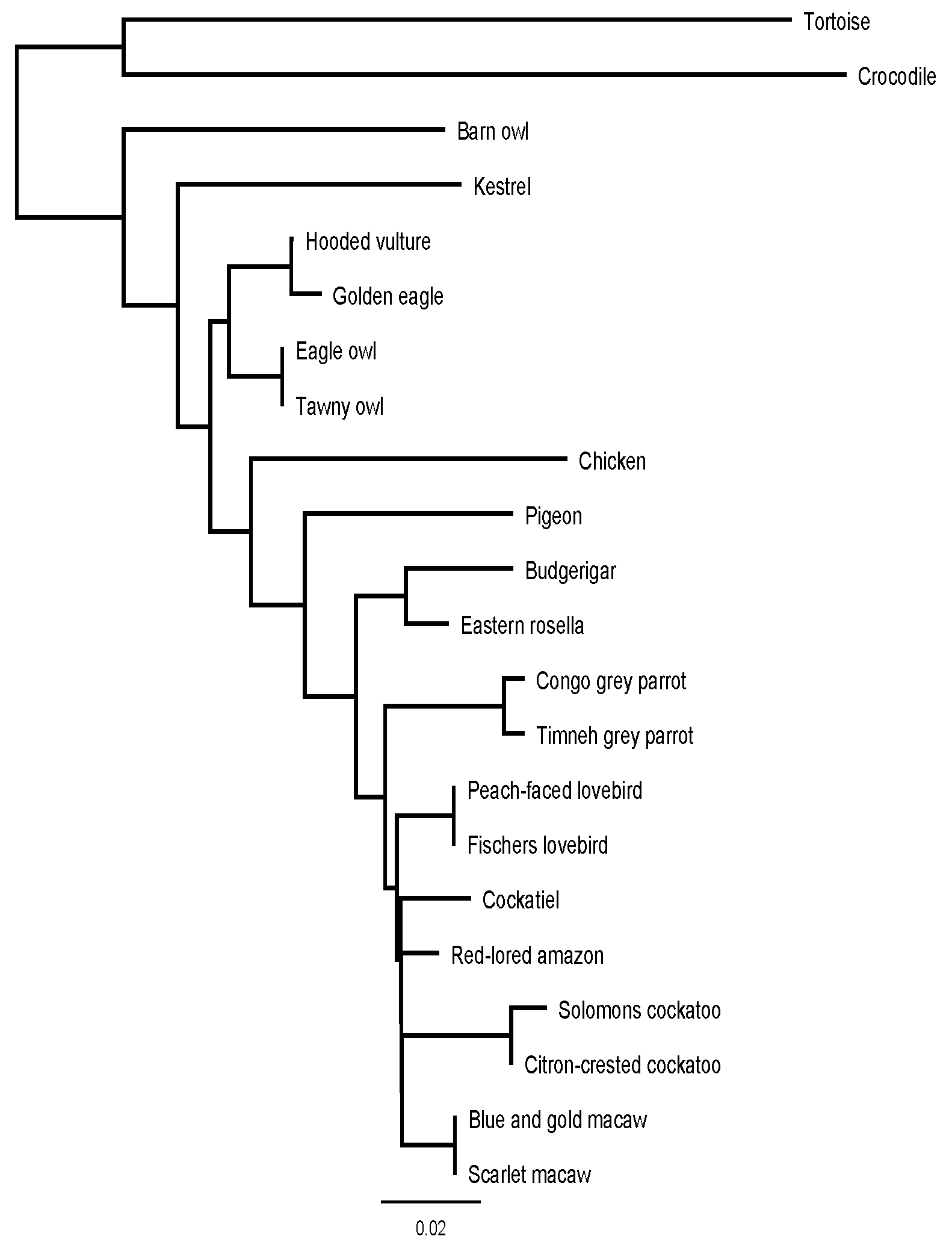
2.4. Analysis
3. Results
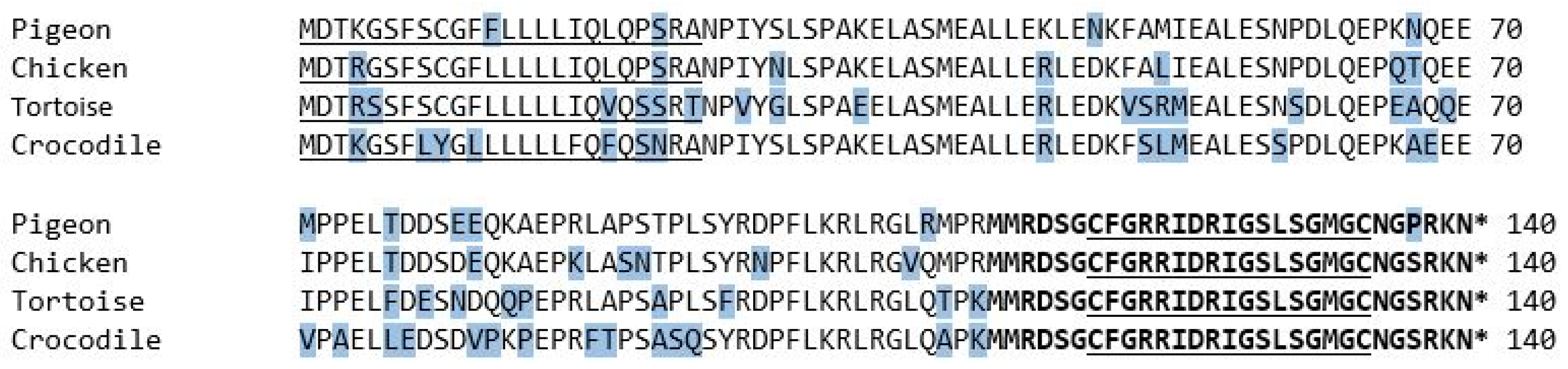
3.1. Mature Peptide Region
3.2. Signal Sequence and NT-pro-BNP
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sudoh, T.; Kangawa, K.; Minamino, N.; Matsuo, H. A new natriuretic peptide in porcine brain. Nature 1988, 332, 78–81. [Google Scholar] [CrossRef] [PubMed]
- Mukoyama, M.; Nakao, K.; Hosoda, K.; Suga, S.; Saito, Y.; Ogawa, Y.; Shirakami, G.; Jougasaki, M.; Obata, K.; Yasue, H. Brain natriuretic peptide as a novel cardiac hormone in humans. Evidence for an exquisite dual natriuretic peptide system, atrial natriuretic peptide and brain natriuretic peptide. J. Clin. Investig. 1991, 87, 1402–1412. [Google Scholar] [CrossRef] [PubMed]
- de Bold, A.J.; Bruneau, B.G.; de Bold, M.L.K. Mechanical and neuroendocrine regulation of the endocrine heart. Cardiovasc. Res. 1996, 31, 7–18. [Google Scholar] [CrossRef] [Green Version]
- Trajanovska, S. Molecular Biology of Natriuretic Peptides in Reptiles and Birds. Doctoral Dissertation, Deakin University, Geelong, Australia, 2008. [Google Scholar]
- Takei, Y.; Inoue, K.; Trajanovska, S.; Donald, J.A. B-type natriuretic peptide (BNP), not ANP, is the principal cardiac natriuretic peptide in vertebrates as revealed by comparative studies. Gen. Comp. Endocrinol. 2011, 171, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Gray, D.A. Plasma atrial natriuretic factor concentrations and renal actions in the domestic fowl. J. Comp. Physiol. B 1993, 163, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Schütz, H.; Gray, D.A.; Gerstberger, R. Modulation of kidney function in conscious Pekin ducks by atrial natriuretic factor. Endocrinology 1992, 130, 678–684. [Google Scholar] [CrossRef] [PubMed]
- Gray, D.A.; Schütz, H.; Gerstberger, R. Interaction of atrial natriuretic factor and osmoregulatory hormones in the Pekin duck. Gen. Comp. Endocrinol. 1991, 81, 246–255. [Google Scholar] [CrossRef]
- Gray, D.A.; Schütz, H.; Gerstberger, R. Plasma atrial natriuretic factor responses to blood volume changes in the Pekin duck. Endocrinology 1991, 128, 1655–1660. [Google Scholar] [CrossRef]
- de Bold, A.J. Atrial natriuretic factor: A hormone produced by the heart. Science 1985, 230, 767. [Google Scholar] [CrossRef]
- Akizuki, N.; Kangawa, K.; Minamino, N.; Matsou, H. Cloning and sequence analysis of complementary DNA encoding a precursor for chicken natriuretic peptide. FEBS Lett. 1991, 280, 357–362. [Google Scholar] [CrossRef] [Green Version]
- Torres-Courchoud, I.; Chen, H.H. B-type natriuretic peptide and acute heart failure: Fluid homeostasis, biomarker and therapeutics. Rev. Clín. Esp. 2016, 216, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Santaguida, P.L.; Don-Wauchope, A.C.; Oremus, M.; McKelvie, R.; Ali, U.; Hill, S.A.; Balion, C.; Booth, R.A.; Brown, J.A.; Bustamam, A.; et al. BNP and NT-proBNP as prognostic markers in persons with acute decompensated heart failure: A systematic review. Heart Fail. Rev. 2014, 19, 453–470. [Google Scholar] [CrossRef] [PubMed]
- Doust, J.A.; Pietrzak, E.; Dobson, A.; Glasziou, P. How well does B-type natriuretic peptide predict death and cardiac events in patients with heart failure: Systematic review. BMJ 2005, 330, 625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, T.; Gegenhuber, A.; Dieplinger, B.; Poelz, W.; Haltmayer, M. Long-term stability of endogenous B-type natriuretic peptide (BNP) and amino terminal proBNP (NT-proBNP) in frozen plasma samples. Clin. Chem. Lab. Med. (CCLM) 2004, 42, 942. [Google Scholar] [CrossRef]
- Weber, M.; Mitrovic, V.; Hamm, C. B-Type Natriuretic Peptide and N-Terminal Pro-B-Type Natriuretic Peptide—Diagnostic Role in Stable Coronary Artery Disease. Exp. Clin. Cardiol. 2006, 11, 99. [Google Scholar]
- Bavelaar, F.J.; Beynen, A.C. Atherosclerosis in parrots. A review. Vet. Q. 2004, 26, 50–60. [Google Scholar] [CrossRef]
- Pees, M.; Krautwald-Junghanns, M.-E.; Straub, J. Evaluating and treating the cardiovaskular system. In Clinical Avian Medicine; Harrison, G.J., Lightfoot, T.L., Eds.; Spix Publishing: Palm Beach, FL, USA, 2006; pp. 379–395. [Google Scholar]
- Miyata, A.; Minamino, N.; Kangawa, K.; Matsuo, H. Identification of a 29-amino acid natriuretic peptide in chicken heart. Biochem. Biophys. Res. Commun. 1988, 155, 1330–1337. [Google Scholar] [CrossRef]
- Pees, M. Echokardiographische Untersuchungen an Klinisch Gesunden Psittaciformes unter Besonderer Berücksichtigung des Kongo-Graupapageis (Psittacus Erithacus Erithacus). Doctoral Dissertation, Universität Leipzig, Leipzig, Germany, 2004. [Google Scholar]
- Hennig, A.; Mohr, L.; Fehr, M.; Legler, M. The B-type natriuretic peptide of the Congo and Timneh grey parrot. Vet. Res. Commun. 2021, 45, 329–333. [Google Scholar] [CrossRef]
- Forshaw, J.M. Parrots of the World; Princeton University Press: Princeton, NJ, USA, 2010. [Google Scholar]
- Hackett, S.J.; Kimball, R.T.; Reddy, S.; Bowie, R.C.K.; Braun, E.L.; Braun, M.J.; Chojnowski, J.L.; Cox, W.A.; Han, K.-L.; Harshman, J.; et al. A Phylogenomic Study of Birds Reveals Their Evolutionary History. Science 2008, 320, 1763–1768. [Google Scholar] [CrossRef]
- Legler, M.; Kummerfeld, N.; Wohlsein, P. Atherosclerosis in birds of prey: A case study and the influence of a one-day-old chicken diet on total plasma cholesterol concentration in different raptor and owl species. Berl. Munch. Tierarztl. Wochenschr. 2017, 130, 353–363. [Google Scholar] [CrossRef]
- Pees, M.; Schmidt, V.; Coles, B.; Krautwald-Junghanns, M.E. Diagnosis and long-term therapy of right-sided heart failure in a yellowcrowned amazon. Vet. Rec. 2006, 158, 445. [Google Scholar] [CrossRef]
- Pees, M.; Straub, J.; Krautwald-Junghanns, M.-E. Insufficiency of the muscular atrioventricular valve in the heart of a bluefronted amazon (Amazona aestiva aestiva). Vet. Rec. 2001, 148, 540–543. [Google Scholar] [CrossRef] [PubMed]
- Rembert, M.S.; Smith, J.A.; Strickland, K.N.; Tully, T.N. Intermittent Bradyarrhythmia in a Hispaniolan Amazon Parrot (Amazona ventralis). J. Avian Med. Surg. 2008, 22, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Beaufrère, H.; Holder, K.A.; Bauer, R.; Schnellbacher, R.; Pariaut, R.; Tully, T.N.; Wakamatsu, N. Intermittent Claudication-like Syndrome Secondary to Atherosclerosis in a Yellow-naped Amazon Parrot. J. Avian Med. Surg. 2011, 25, 266–276. [Google Scholar] [CrossRef]
- Beaufrere, H.; Ammersbach, M.; Reavill, D.R.; Garner, M.M.; Heatley, J.J.; Wakamatsu, N.; Nevarez, J.G.; Tully, T.N. Prevalence of and risk factors associated with atherosclerosis in psittacine birds. J. Am. Vet. Med. Assoc. 2013, 242, 1696–1704. [Google Scholar] [CrossRef]
- Juan-Sallés, C.; Soto, S.; Garner, M.M.; Montesinos, A.; Ardiaca, M. Congestive Heart Failure in 6 African Grey Parrots (Psittacus e erithacus). Vet. Pathol. 2011, 48, 691–697. [Google Scholar] [CrossRef]
- Hennig, A.; Legler, M.; Mohr, L.; Lindenwald, R.; Haase, C.; Fehr, M. The B-Type natriuretic peptide (BNP) of the Congo Grey Parrot. In Proceedings of the International Conference on Avian Herpatological and Exotic Mammal Medicine, London, UK, 28 April–2 May 2019. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Position of Substitution |
|---|---|
| Congo grey parrot | 5, 10, 22, 65 |
| Timneh grey parrot | 5, 10, 22, 65 |
| Blue and gold macaw | 59 |
| Scarlet macaw | 59 |
| Solomons cockatoo | 3, 8, 91, 105 |
| Citron-cr. Cockatoo | 3, 8, 91 |
| Cockatiel | 61, 98 |
| Peach-faced lovebird | 64, 76 |
| Fischer’s lovebird | 64, 76 |
| Budgerigar | 6, 67, 139, 140 |
| Eastern rosella | 22, 71, 139 |
| Red lored amazon | 68 |
| Barn owl | 3, 19, 21, 22, 29, 45, 57, 67, 71, 73, 74, 76, 96, 99, 106 |
| Eagle owl | 21, 22, 29, 45, 67, 76, 79 |
| Tawny owl | 21, 22, 29, 45, 67, 76, 79 |
| Hooded vulture | 21, 22, 45, 67, 76, 92 |
| Golden eagle | 21, 22, 45, 67, 76, 78, 92 |
| Kestrel | 21, 22, 45, 67, 72, 75, 76, 84, 86, 91, 94, 99 |
| Pigeon | 22, 48, 67, 71, 76, 80, 81, 137 |
| Congo Grey Parrot | Timneh Grey Parrot | Blue and Gold Macaw | Scarlet Macaw | Solomons Cockatoo | Citron-cr. Cockatoo | Cockatiel | Peach-Faced Lovebird | Fischer’s Lovebird | Budgerigar | Eastern Rosella | Red-Lored Amazon | Barn Owl | Eagle Owl | Tawny Owl | Hooded Vulture | Golden Eagle | Kestrel | Pigeon | Chicken | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Congo g. parrot Timneh g. parrot B.-and-g. macaw Scarlet macaw Solomons c. Citron-cr. c. Cockatiel Peach-faced l. Fischer’s l. Budgerigar Eastern rosella Red-lored amazon Barn owl Eagle owl Tawny owl Hooded vulture Golden eagle Kestrel Pigeon Chicken | 100.00 | 99.29 100.00 | 96.43 96.43 100.00 | 96.43 96.43 100.00 100.00 | 93.57 93.57 95.71 95.71 100.00 | 95.00 95.00 97.14 97.14 98.57 100.00 | 95.71 95.71 97.86 97.86 95.00 96.43 100.00 | 95.71 95.71 97.86 97.86 95.00 96.43 97.14 100.00 | 95.71 95.71 97.86 97.86 95.00 96.43 97.14 100.00 100.00 | 92.14 92.14 94.29 94.29 91.43 92.86 93.57 93.57 93.57 100.00 | 95.00 95.00 96.43 96.43 93.57 95.00 95.71 95.71 95.71 93.57 100.00 | 95.71 95.71 97.86 97.86 95.00 96.43 97.14 97.14 97.14 95.00 95.71 100.00 | 87.14 87.14 88.57 88.57 87.14 88.57 87.86 88.57 88.57 85.71 89.29 87.86 100.00 | 92.86 92.86 94.29 94.29 91.43 92.86 93.57 95.00 95.00 92.86 93.57 94.29 91.43 100.00 | 92.86 92.86 94.29 94.29 91.43 92.86 93.57 95.00 95.00 92.86 93.57 94.29 91.43 100.00 100.00 | 93.53 93.53 94.96 94.96 92.09 93.53 94.24 95.68 95.68 92.81 94.24 94.24 89.93 97.12 97.12 100.00 | 92.81 92.81 94.24 94.24 91.37 92.81 93.53 94.96 94.96 92.09 93.53 93.53 89.21 96.40 96.40 99.28 100.00 | 88.57 88.57 90.00 90.00 87.86 89.29 89.29 90.71 90.71 87.14 89.29 89.29 87.14 92.14 92.14 92.09 91.37 100.00 | 91.43 91.43 92.86 92.86 90.00 91.43 92.14 92.86 92.86 90.00 93.57 92.14 87.14 92.14 92.14 92.81 92.09 88.57 100.00 | 87.86 87.86 89.29 89.29 87.86 89.29 90.00 89.29 89.29 86.43 88.57 88.57 85.71 91.43 91.43 90.65 89.93 85.71 89.29 100.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hennig, A.; Mohr, L.; Fehr, M.; Legler, M. Analysis of the B-Type Natriuretic Peptide and the Aminoterminal-Pro-B-Type Natriuretic Peptide in Different Parrot, Raptor and Owl Species. Vet. Sci. 2022, 9, 64. https://doi.org/10.3390/vetsci9020064
Hennig A, Mohr L, Fehr M, Legler M. Analysis of the B-Type Natriuretic Peptide and the Aminoterminal-Pro-B-Type Natriuretic Peptide in Different Parrot, Raptor and Owl Species. Veterinary Sciences. 2022; 9(2):64. https://doi.org/10.3390/vetsci9020064
Chicago/Turabian StyleHennig, Anja, Lydia Mohr, Michael Fehr, and Marko Legler. 2022. "Analysis of the B-Type Natriuretic Peptide and the Aminoterminal-Pro-B-Type Natriuretic Peptide in Different Parrot, Raptor and Owl Species" Veterinary Sciences 9, no. 2: 64. https://doi.org/10.3390/vetsci9020064
APA StyleHennig, A., Mohr, L., Fehr, M., & Legler, M. (2022). Analysis of the B-Type Natriuretic Peptide and the Aminoterminal-Pro-B-Type Natriuretic Peptide in Different Parrot, Raptor and Owl Species. Veterinary Sciences, 9(2), 64. https://doi.org/10.3390/vetsci9020064