
Multimodal Assessment of Bottlenose Dolphin Auditory Nuclei Using 7-Tesla MRI, Immunohistochemistry and Stereology

 , , ,
, , ,  ,
,  ,
,  ,
,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Specimens
2.2. MRI and DTI
2.3. Western Blot and BLAST
2.4. Immunohistochemistry
2.5. Microscopy and Stereology
2.6. Data Analysis
3. Results
3.1. MRI
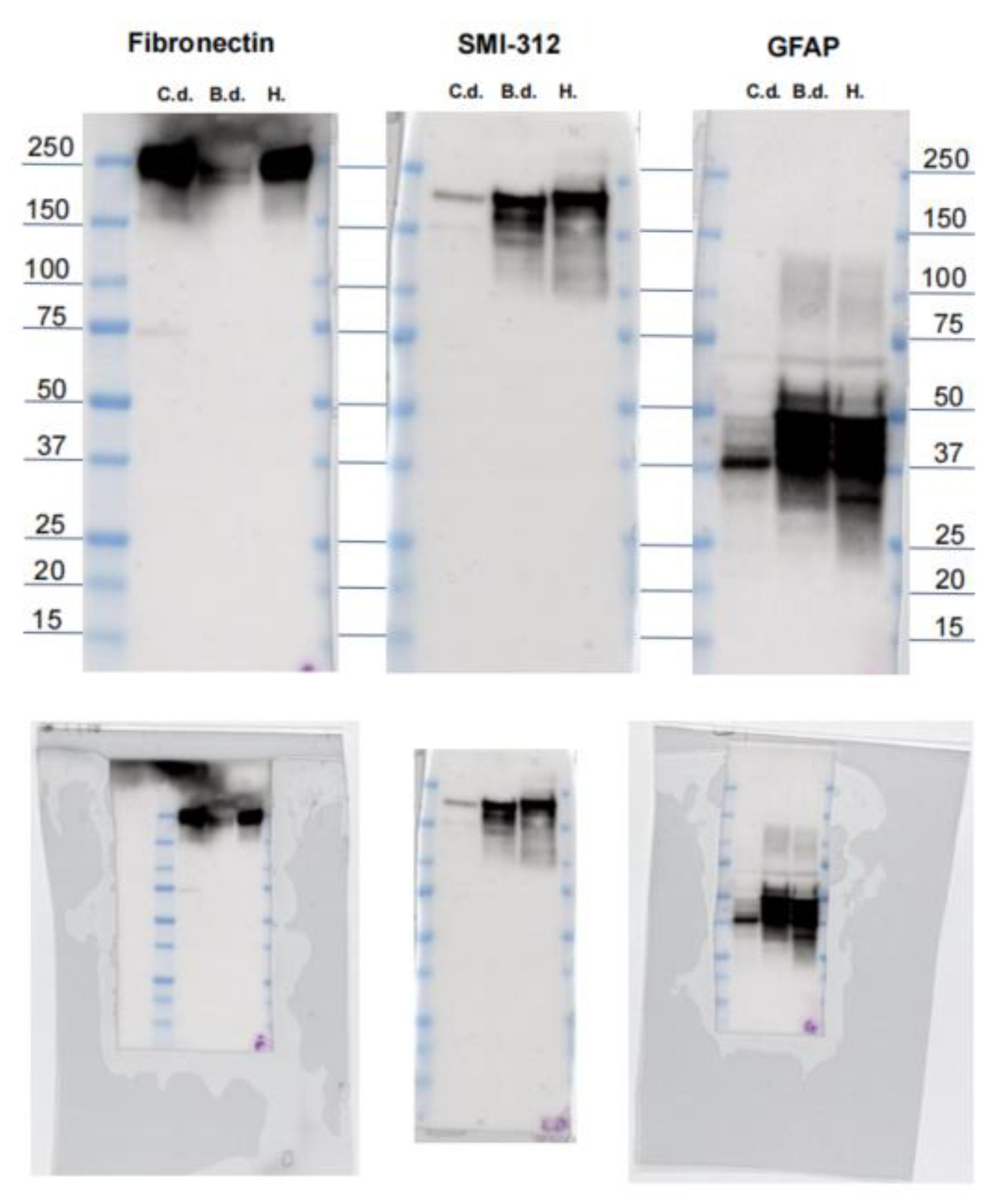
3.2. Western Blot and BLAST
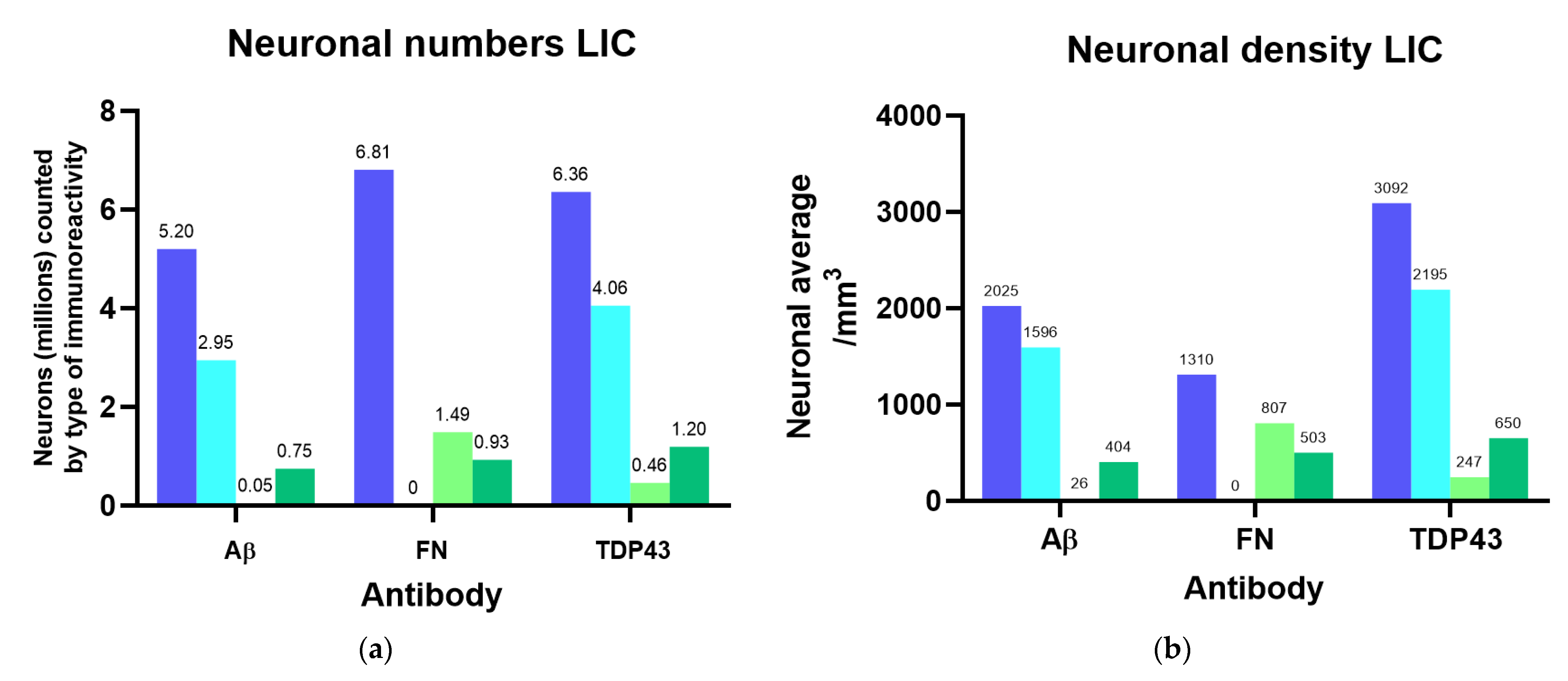
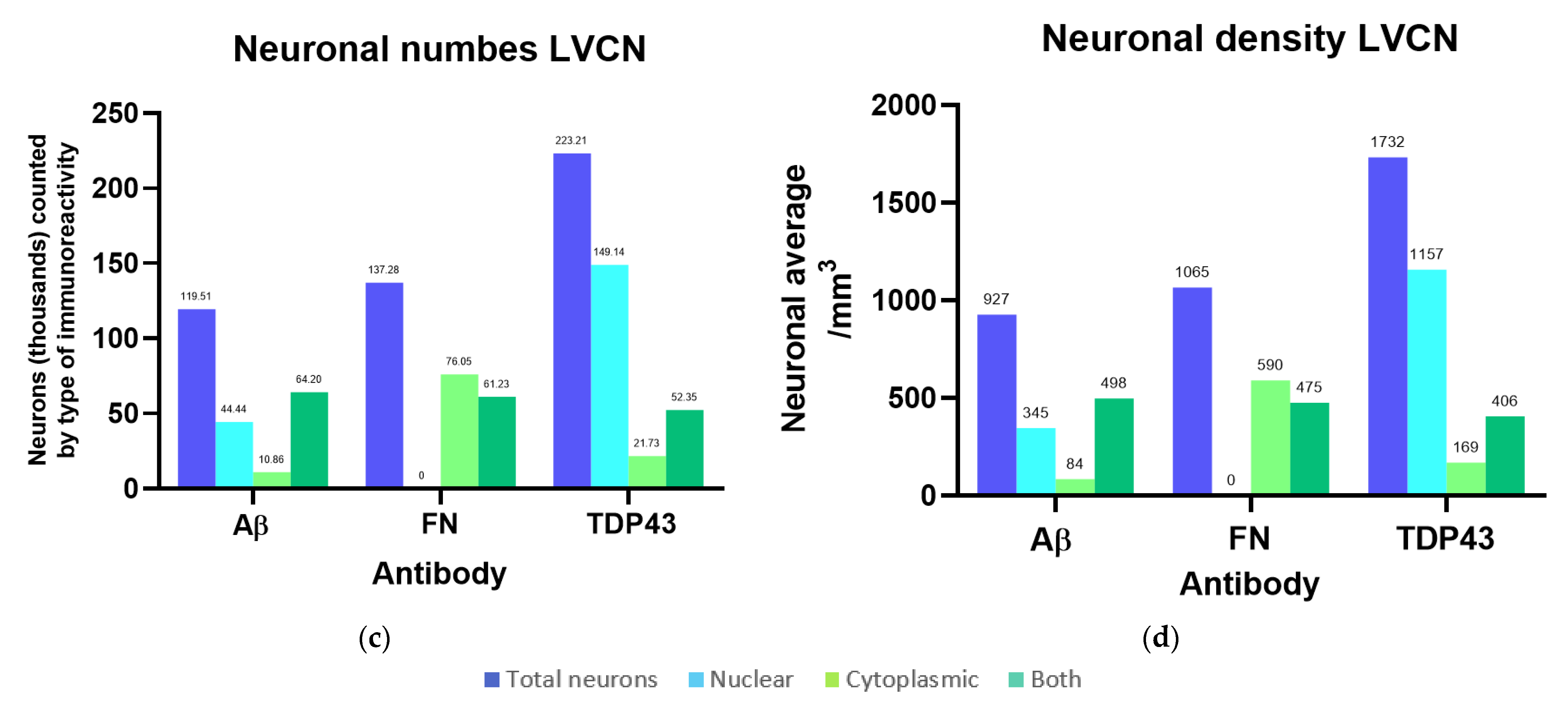
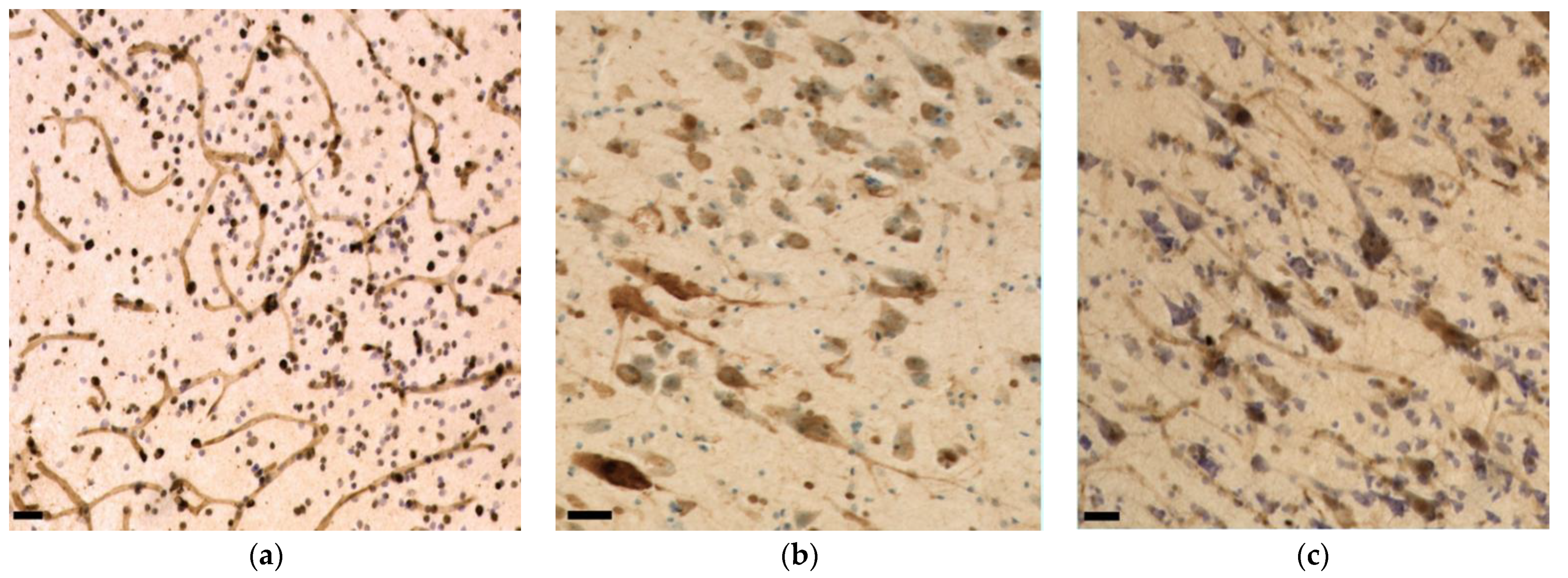
3.3. Immunohistochemistry and Stereology
3.3.1. Aβ
3.3.2. Fibronectin
3.3.3. TDP-43
3.3.4. pNFP
3.3.5. Iba-1
3.3.6. GFAP
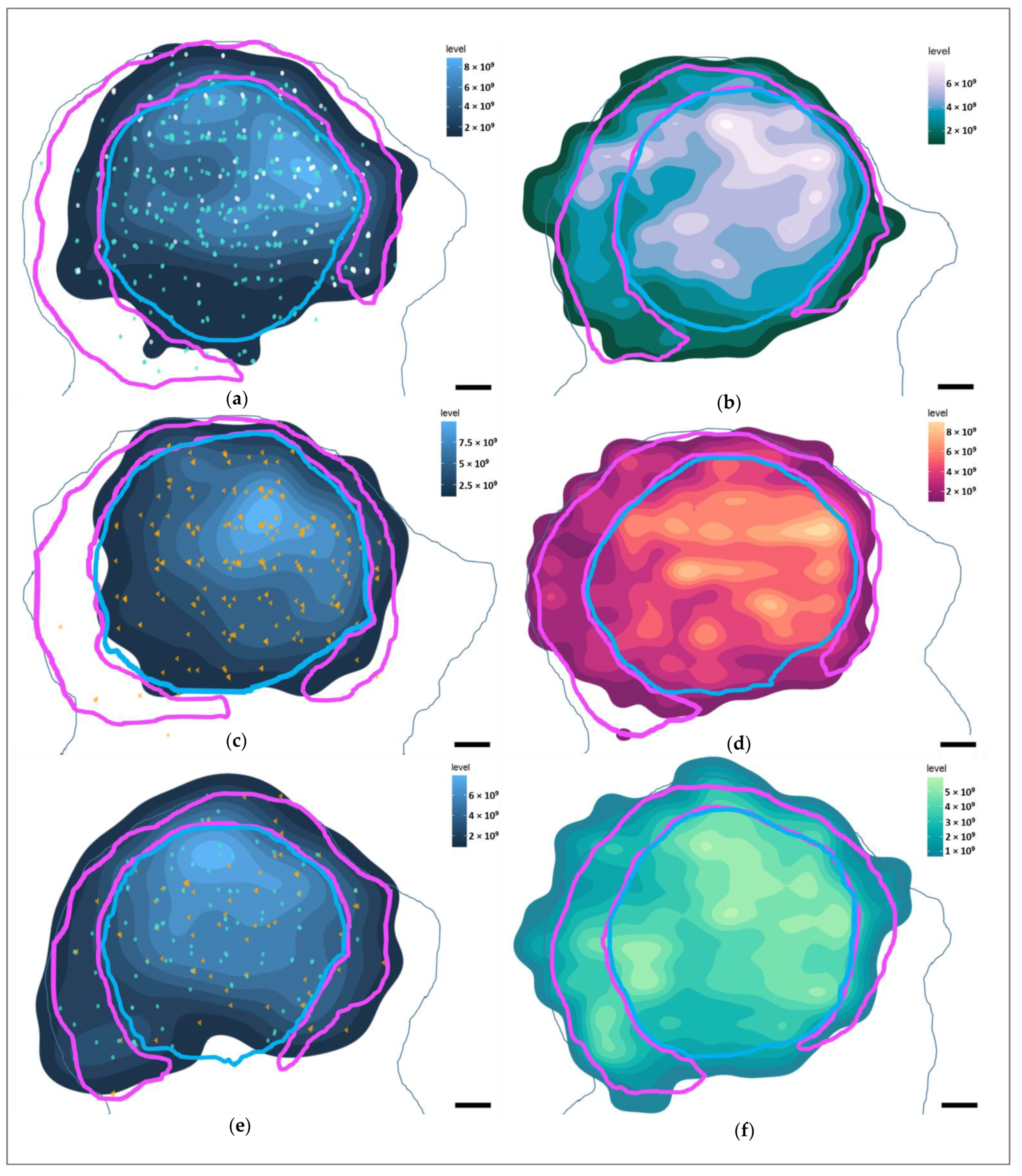
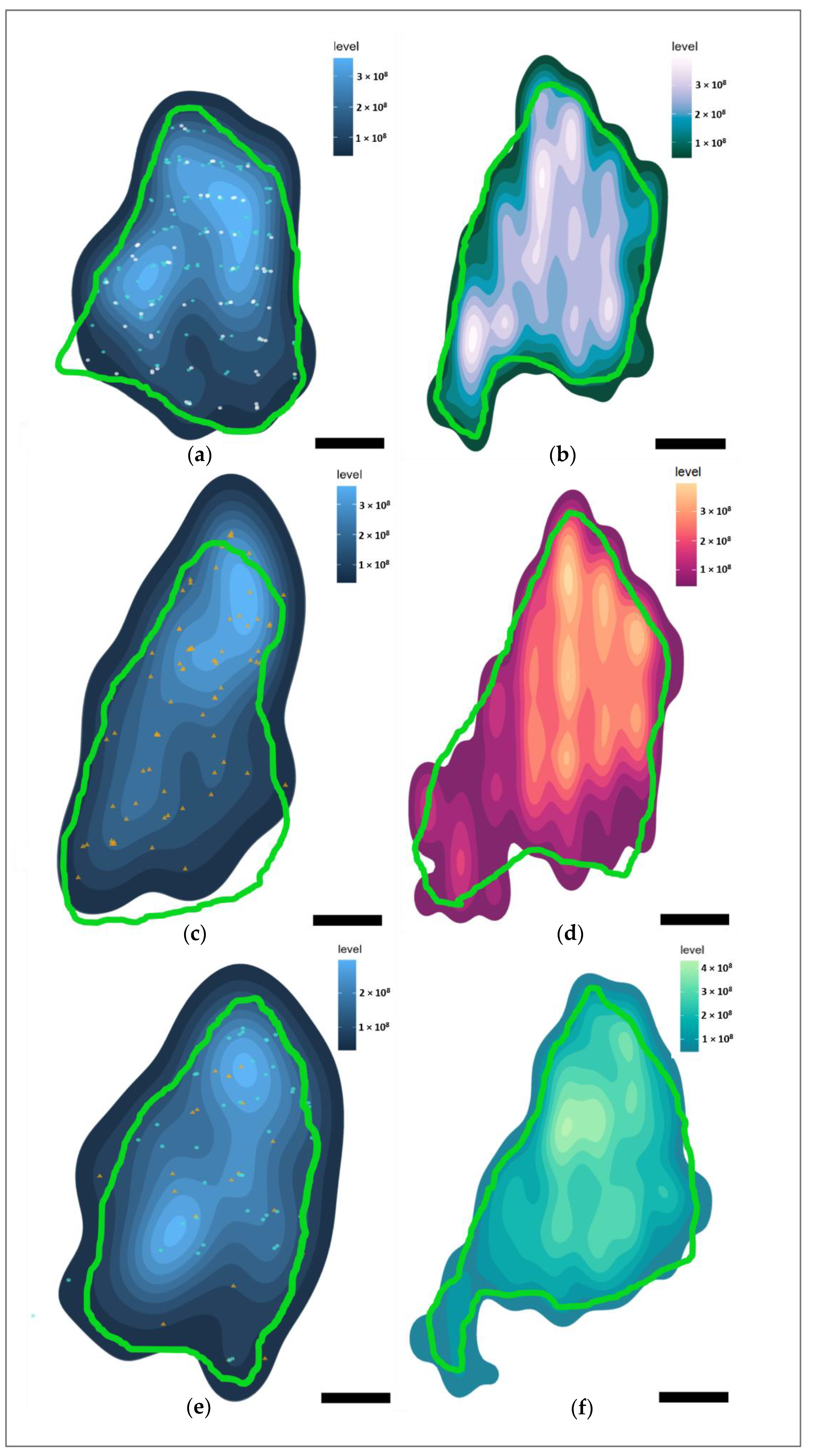
3.4. Marker Distributions
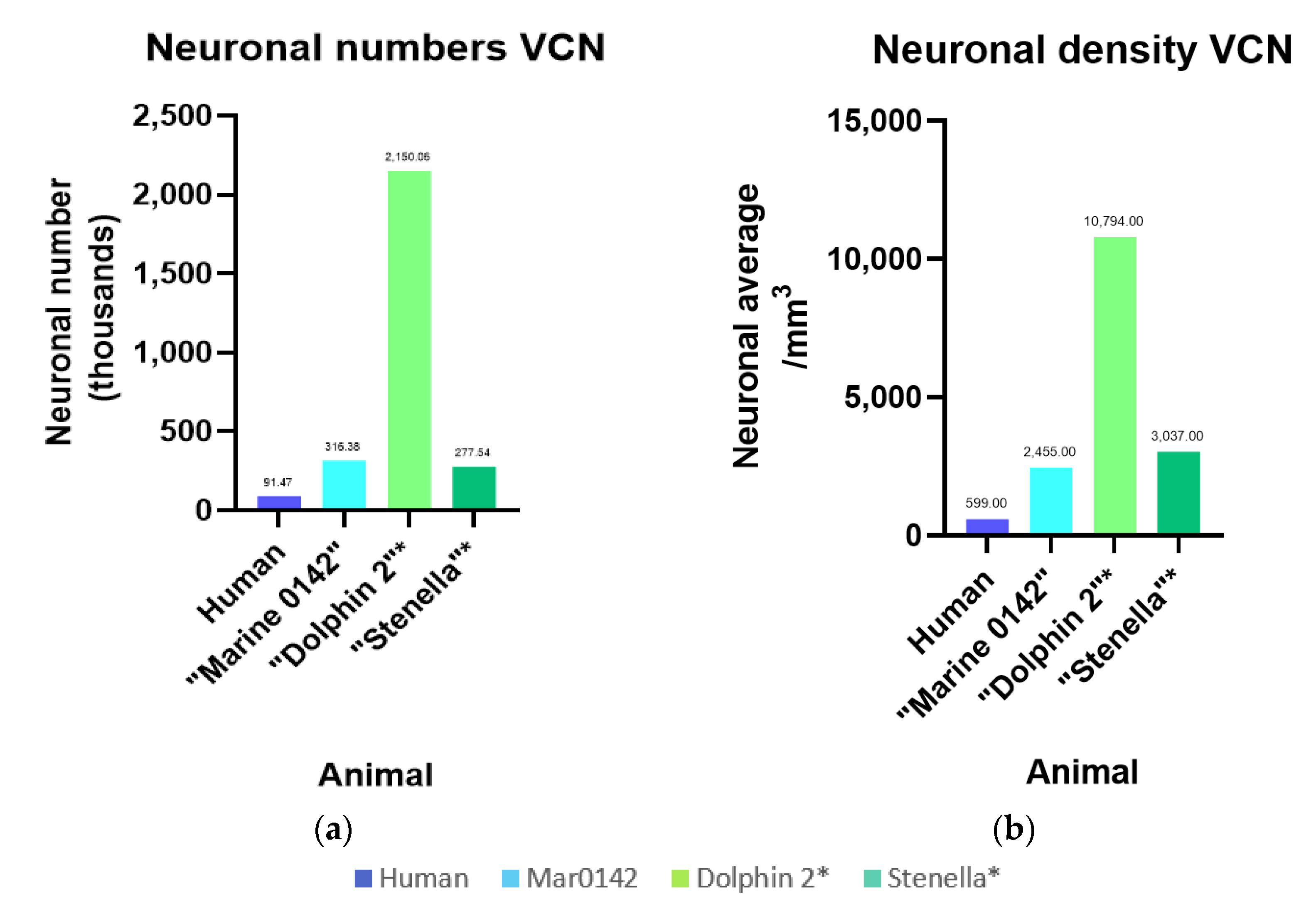
3.5. Intra- and Interspecies Comparisons
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody 1 | Host | Reactivity | Clonality | RRID | Catalogue Number | Lowest Dilution IHC |
|---|---|---|---|---|---|---|
| AT-8 | Mouse | Human, Mouse, Rat (etc.) | Monoclonal | AB_223647 | MN1020 | 1:1000 |
| CD-31 | Rabbit | Human | Polyclonal | AB_726362 | ab28364 | 1:100 |
| Collagen IV | Rabbit | Mouse, Rat, Hamster, Cow, Dog, Human, Pig, Zebrafish, African green monkey, Chinese hamster, Syrian hamster | Polyclonal | AB_445160 | ab6586 | 1:300 |
| Collagen IV | Rabbit | Mouse | Polyclonal | AB_445160 | ab19808 | 1:300 |
| Collagen IV | Rabbit | Human | Monoclonal | AB_2801511 | ab214417 | 1:500 |
| CP-13 | Mouse | Human | Monoclonal | AB_2314223 | Davies Lab | 1:1000 |
| dMBP | Rabbit | Human, Rat | Polyclonal | AB_2140351 | ab5864 | 1:500 |
| GFAP | Rabbit | Mouse, Rat, Human | Monoclonal | AB_880202 | ab68426 | 1:1000 |
| Isolectin B4 2 | Bandeiraea simplicifolia | Non-immune origin | Unspecific binding of glycoproteins | NA | L2140 | 1:40 |
| MAP-2 | Rabbit | Human | Polyclonal | AB_1853945 | HPA008273 | 1:200 |
| MMP-9 | Mouse | Human, Mouse, Rat | Monoclonal | Not available | NBP2-80855 | 1:300 |
| MOAB-2 | Mouse | Human | Monoclonal | AB_2895168 | MABN254 | 1:200 |
| NeuN | Mouse | Avian, Chicken, Ferret, Human, Mouse, Pig, Rat, Salamander | Monoclonal | AB_2298772 | MAB377 | 1:200 |
| PHF1 | Mouse | Human | Monoclonal | AB_2315150 | Davies Lab | 1:500 |
| SMI-32 | Mouse | Human, Mouse, Rat | Monoclonal | AB_2715852 | 801701 | 1:1000 |
| Vimentin | Mouse | Human, Rat | Monoclonal | AB_306239 | ab8069 | 1:200 |
| Data File | Marker | Total Markers Counted | Number of Sampling Sites | Estimated Population Using Mean Section Thickness with Counts | CE (Gundersen), m = 0 | CE (Gundersen), m = 1 | 1st Estimated CE (Schmitz- Hof) | 2nd Estimated CE (Schmitz- Hof) |
|---|---|---|---|---|---|---|---|---|
| Amyloid-β | ||||||||
| LIC CN | Non-ir | 162 | 239 | 960,000.06 | 0.11 | 0.08 | 0.099 | 0.079 |
| LIC CN | Nuclear | 484 | 239 | 2,868,148.5 | 0.06 | 0.05 | 0.057 | 0.045 |
| LIC CN | Cytoplasmic | 7 | 239 | 41481.48 | 0.38 | 0.38 | 0.470 | 0.378 |
| LIC CN | Nuclear + cytoplasmic | 33 | 239 | 195,555.56 | 0.20 | 0.18 | 0.202 | 0.174 |
| LIC CN | All markers | 686 | 239 | 4,065,185.25 | 0.05 | 0.04 | 0.046 | 0.038 |
| LIC EC | Non-ir | 14 | 153 | 82,962.96 | 0.28 | 0.27 | 0.370 | 0.267 |
| LIC EC | Nuclear | 82 | 153 | 485,925.94 | 0.16 | 0.11 | 0.172 | 0.110 |
| LIC EC | Cytoplasmic | 1 | 153 | 5925.93 | 1.00 | 1.00 | 1.000 | 1.000 |
| LIC EC | Nuclear + cytoplasmic | 95 | 153 | 562,963 | 0.12 | 0.10 | 0.157 | 0.103 |
| LIC EC | All markers | 192 | 153 | 1,137,777.88 | 0.1 | 0.07 | 0.109 | 0.072 |
| LVCN | Non-ir | 45 | 137 | 44,444.45 | 0.21 | 0.15 | 0.312 | 0.149 |
| LVCN | Nuclear | 128 | 137 | 126,419.76 | 0.13 | 0.09 | 0.105 | 0.088 |
| LVCN | Cytoplasmic | 11 | 137 | 10,864.20 | 0.34 | 0.30 | 0.343 | 0.302 |
| LVCN | Nuclear + cytoplasmic | 65 | 137 | 64,197.54 | 0.14 | 0.12 | 0.129 | 0.124 |
| LVCN | All markers | 249 | 137 | 245,925.94 | 0.1 | 0.07 | 0.083 | 0.063 |
| Fibronectin | ||||||||
| LIC CN | Non-ir | 594 | 235 | 3,520,000.25 | 0.05 | 0.04 | 0.049 | 0.041 |
| LIC CN | Cytoplasmic | 217 | 235 | 1,285,926.13 | 0.09 | 0.07 | 0.085 | 0.068 |
| LIC CN | Nuclear + cytoplasmic | 129 | 235 | 764,444.5 | 0.12 | 0.09 | 0.100 | 0.088 |
| LIC CN | All markers | 940 | 235 | 5,570,370.5 | 0.04 | 0.03 | 0.045 | 0.033 |
| LIC EC | Non-ir | 147 | 100 | 871,111.19 | 0.10 | 0.08 | 0.123 | 0.082 |
| LIC EC | Cytoplasmic | 35 | 100 | 207,407.41 | 0.19 | 0.17 | 0.192 | 0.169 |
| LIC EC | Nuclear + cytoplasmic | 28 | 100 | 165,925.92 | 0.22 | 0.19 | 0.222 | 0.189 |
| LIC EC | All markers | 210 | 100 | 1,244,444.5 | 0.09 | 0.07 | 0.103 | 0.069 |
| LVCN | Non-ir | 300 | 145 | 296,296.31 | 0.12 | 0.06 | 0.099 | 0.058 |
| LVCN | Cytoplasmic | 77 | 145 | 76,049.38 | 0.12 | 0.11 | 0.133 | 0.114 |
| LVCN | Nuclear + cytoplasmic | 62 | 145 | 61,234.57 | 0.14 | 0.13 | 0.137 | 0.127 |
| LVCN | All markers | 439 | 145 | 433,580.28 | 0.1 | 0.05 | 0.072 | 0.048 |
| TDP-43 | ||||||||
| LIC CN | Non-ir | 555 | 248 | 3,288,889 | 0.07 | 0.04 | 0.051 | 0.042 |
| LIC CN | Nuclear | 74 | 248 | 438,518.59 | 0.12 | 0.12 | 0.118 | 0.116 |
| LIC CN | Cytoplasmic | 60 | 248 | 355,555.56 | 0.16 | 0.13 | 0.170 | 0.129 |
| LIC CN | Nuclear + cytoplasmic | 140 | 248 | 829,629.63 | 0.12 | 0.09 | 0.108 | 0.085 |
| LIC CN | All markers | 829 | 248 | 49,12593 | 0.05 | 0.04 | 0.044 | 0.035 |
| LIC EC | Non-ir | 130 | 106 | 770,370.5 | 0.14 | 0.09 | 0.150 | 0.088 |
| LIC EC | Nuclear | 35 | 106 | 207,407.41 | 0.19 | 0.17 | 0.218 | 0.169 |
| LIC EC | Cytoplasmic | 17 | 106 | 100,740.75 | 0.24 | 0.24 | 0.253 | 0.243 |
| LIC EC | Nuclear + cytoplasmic | 63 | 106 | 373,333.34 | 0.16 | 0.13 | 0.161 | 0.126 |
| LIC EC | All markers | 245 | 106 | 1,451,852 | 0.12 | 0.07 | 0.11 | 0.064 |
| LVCN | Non-ir | 151 | 140 | 149,135.81 | 0.14 | 0.09 | 0.112 | 0.081 |
| LVCN | Nuclear | 47 | 140 | 46,419.76 | 0.19 | 0.15 | 0.150 | 0.146 |
| LVCN | Cytoplasmic | 22 | 140 | 21,728.40 | 0.22 | 0.21 | 0.217 | 0.213 |
| LVCN | Nuclear + cytoplasmic | 53 | 140 | 52,345.68 | 0.29 | 0.15 | 0.148 | 0.137 |
| pNFP | ||||||||
| LIC CN | Axonal | 3970 | 213 | 2,299,468,544 | 0.06 | 0.02 | - | - |
| LIC EC | Axonal | 1795 | 76 | 1,058,085,632 | 0.09 | 0.03 | - | - |
| LVCN | Axonal | 1988 | 115 | 195,308,656 | 0.13 | 0.04 | - | - |
| Iba-1 | ||||||||
| LIC CN | Microglial | 2738 | 227 | 405,629,632 | 0.06 | 0.02 | 0.035 | 0.019 |
| LIC EC | Microglial | 2135 | 122 | 316,296,320 | 0.08 | 0.03 | 0.047 | 0.022 |
| LVCN | Microglial | 1594 | 139 | 39,358,024 | 0.09 | 0.03 | 0.055 | 0.025 |
| GFAP | ||||||||
| LIC CN | Astrocytic | 4840 | 229 | 717,036,992 | 0.05 | 0.02 | 0.03 | 0.014 |
| LIC EC | Astrocytic | 2662 | 96 | 394,370,400 | 0.08 | 0.03 | 0.058 | 0.019 |
| LVCN | Astrocytic | 3128 | 137 | 77,234,568 | 0.08 | 0.02 | 0.043 | 0.018 |

References
- Hof, P.E.; Chanis, R.; Marino, L. Cortical complexity in cetacean brains. Anat. Rec.—Part A Discov. Mol. Cell. Evol. Biol. 2005, 287, 1142–1152. [Google Scholar] [CrossRef]
- Gunn-Moore, D.; Kaidanovich-Beilin, O.; Gallego Iradi, M.C.; Gunn-Moore, F.; Lovestone, S. Alzheimer’s disease in humans and other animals: A consequence of postreproductive life span and longevity rather than aging. Alzheimer’s Dement. 2018, 14, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Di Guardo, G. Cetaceans, models for human disease? Res. Vet. Sci. 2018, 119, 43–44. [Google Scholar] [CrossRef] [PubMed]
- Davis, D.A.; Garamszegi, S.P.; Banack, S.A.; Dooley, P.D.; Coyne, T.M.; McLean, D.W.; Rotstein, D.S.; Mash, D.C.; Cox, P.A. Bmaa, methylmercury, and mechanisms of neurodegeneration in dolphins: A natural model of toxin exposure. Toxins 2021, 13, 697. [Google Scholar] [CrossRef] [PubMed]
- Orekhova, K.; Centelleghe, C.; Di Guardo, G.; Graïc, J.M.; Cozzi, B.; Trez, D.; Verin, R.; Mazzariol, S. Systematic validation and assessment of immunohistochemical markers for central nervous system pathology in cetaceans, with emphasis on auditory pathways. PLoS ONE 2022, 17, e026909. [Google Scholar] [CrossRef] [PubMed]
- Fernández, A.; Edwards, J.F.; Rodríguez, F.; Espinosa De Los Monteros, A.; Herráez, P.; Castro, P.; Jaber, J.R.; Martín, V.; Arbelo, M. “Gas and fat embolic syndrome” involving a mass stranding of beaked whales (Family ziphiidae) exposed to anthropogenic sonar signals. Vet. Pathol. 2005, 42, 446–457. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Sen, N. Traumatic brain injury: A risk factor for neurodegenerative diseases. Rev. Neurosci. 2016, 27, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Lillie, M.A.; Vogl, A.W.; Gerard, S.G.; Raverty, S.; Shadwick, R.E. Retia mirabilia: Protecting the cetacean brain from locomotion-generated blood pressure pulses. Science 2022, 377, 1452–1456. [Google Scholar] [CrossRef] [PubMed]
- Ridgway, S.H.; Brownson, R.H.; Van Alstyne, K.R.; Hauser, R.A. Higher neuron densities in the cerebral cortex and larger cerebellums may limit dive times of delphinids compared to deep-diving toothed whales. PLoS ONE 2019, 14, e0226206. [Google Scholar] [CrossRef]
- Cantú-Medellín, N.; Byrd, B.; Hohn, A.; Vázquez-Medina, J.P.; Zenteno-Savín, T. Differential antioxidant protection in tissues from marine mammals with distinct diving capacities. Shallow/short vs. deep/long divers. Comp. Biochem. Physiol.—Part A Mol. Integr. Physiol. 2011, 158, 438–443. [Google Scholar] [CrossRef]
- Manger, P.R.; Patzke, N.; Spocter, M.A.; Bhagwandin, A.; Karlsson, K.; Bertelsen, M.F.; Alagaili, A.N.; Bennett, N.C.; Mohammed, O.B.; Herculano-Houzel, S.; et al. Amplification of potential thermogenetic mechanisms in cetacean brains compared to artiodactyl brains. Sci. Rep. 2021, 11, 5486. [Google Scholar] [CrossRef]
- Marino, L.; Butti, C.; Connor, R.C.; Fordyce, R.E.; Herman, L.M.; Hof, P.R.; Lefebvre, L.; Lusseau, D.; McCowan, B.; Nimchinsky, E.A.; et al. A claim in search of evidence: Reply to Manger’s thermogenesis hypothesis of cetacean brain structure. Biol. Rev. 2008, 83, 417–440. [Google Scholar] [CrossRef] [PubMed]
- Nieder, C.; Rosene, D.L.; Mortazavi, F.; Oblak, A.L.; Ketten, D.R. Morphology and unbiased stereology of the lateral superior olive in the short-beaked common dolphin, Delphinus delphis (Cetacea, Delphinidae). J. Morphol. 2022, 283, 446–461. [Google Scholar] [CrossRef]
- Simona, S.; Antonio, E.D.L.M.; Yania, P. Amyloid-beta peptide and phosphorylated tau in the frontopolar cerebral cortex and in the cerebellum of toothed whales: Aging vs. hypoxia Summary Statement. Biol. Open 2020, 9, bio054734. [Google Scholar]
- Jo, M.; Lee, S.; Jeon, Y.M.; Kim, S.; Kwon, Y.; Kim, H.J. The role of TDP-43 propagation in neurodegenerative diseases: Integrating insights from clinical and experimental studies. Exp. Mol. Med. 2020, 52, 1652–1662. [Google Scholar] [CrossRef]
- Reed, M.J.; Damodarasamy, M.; Banks, W.A. The extracellular matrix of the blood-brain barrier: Structural and functional roles in health, aging, and Alzheimer’s disease. Tissue Barriers 2019, 7, 1651157. [Google Scholar] [CrossRef]
- Wang, J.; Milner, R. Fibronectin promotes brain capillary endothelial cell survival and proliferation through alpha5beta1 and alphavbeta3 integrins via MAP kinase signalling. J. Neurochem. 2006, 96, 148–159. [Google Scholar] [CrossRef]
- Boero, J.A.; Ascher, J.; Arregui, A.; Rovainen, C.; Woolsey, T.A. Increased brain capillaries in chronic hypoxia. J. Appl. Physiol. 1999, 86, 1211–1219. [Google Scholar] [CrossRef]
- LaManna, J.C.; Chavez, J.C.; Pichiule, P. Structural and functional adaptation to hypoxia in the rat brain. J. Exp. Biol. 2004, 207, 3163–3169. [Google Scholar] [CrossRef]
- Wang, J.T.; Yin, L.; Chen, Z. Neuroprotective role of fibronectin in neuron-glial extrasynaptic transmission. Neural Regen. Res. 2013, 8, 376–382. [Google Scholar] [CrossRef]
- Hof, P.R.; Van Der Gucht, E. Structure of the cerebral cortex of the humpback whale, Megaptera novaeangliae (Cetacea, Mysticeti, Balaenopteridae). Anat. Rec. Part A Discov. Mol. Cell. Evol. Biol. 2006, 31, 175. [Google Scholar] [CrossRef]
- Ackermans, N.L.; Varghese, M.; Williams, T.M.; Grimaldi, N.; Selmanovic, E.; Alipour, A.; Balchandani, P.; Reidenberg, J.S.; Hof, P.R. Evidence of traumatic brain injury in headbutting bovids. Acta Neuropathol. 2022, 144, 5–26. [Google Scholar] [CrossRef]
- Boonstra, J.T.; Michielse, S.; Roebroeck, A.; Temel, Y.; Jahanshahi, A. Dedicated container for postmortem human brain ultra-high field magnetic resonance imaging. Neuroimage 2021, 235, 118010. [Google Scholar] [CrossRef]
- Griswold, M.A.; Jakob, P.M.; Heidemann, R.M.; Nittka, M.; Jellus, V.; Wang, J.; Kiefer, B.; Haase, A. Generalized autocalibrating partially parallel acquisitions (GRAPPA). Magn. Reson. Med. 2002, 47, 1202–1210. [Google Scholar] [CrossRef]
- Jenkinson, M.; Smith, S. A global optimisation method for robust affine registration of brain images. Med. Image Anal. 2001, 5, 143–156. [Google Scholar] [CrossRef]
- Jenkinson, M.; Bannister, P.; Brady, M.; Smith, S. Improved optimization for the robust and accurate linear registration and motion correction of brain images. Neuroimage 2002, 17, 825–841. [Google Scholar] [CrossRef]
- McNab, J.A.; Jbabdi, S.; Deoni, S.C.L.; Douaud, G.; Behrens, T.E.J.; Miller, K.L. High resolution diffusion-weighted imaging in fixed human brain using diffusion-weighted steady state free precession. Neuroimage 2009, 46, 775–785. [Google Scholar] [CrossRef]
- Andersson, J.L.R.; Sotiropoulos, S.N. An integrated approach to correction for off-resonance effects and subject movement in diffusion MR imaging. Neuroimage 2016, 125, 1063–1078. [Google Scholar] [CrossRef] [PubMed]
- Yeh, F.C.; Verstynen, T.D.; Wang, Y.; Fernández-Miranda, J.C.; Tseng, W.Y.I. Deterministic Diffusion Fiber Tracking Improved by Quantitative Anisotropy. PLoS ONE 2013, 8, e80713. [Google Scholar] [CrossRef]
- Yushkevich, P.A.; Piven, J.; Hazlett, H.C.; Smith, R.G.; Ho, S.; Gee, J.C.; Gerig, G. User-guided 3D active contour segmentation of anatomical structures: Significantly improved efficiency and reliability. Neuroimage 2006, 31, 1116–1128. [Google Scholar] [CrossRef]
- Gundersen, H.J.G.; Jensen, E.B.V.; Kiêu, K.; Nielsen, J. The efficiency of systematic sampling in stereology—Reconsidered. J. Microsc. 1999, 193, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Arifuku, S. On the Acoustic System in the Cetacean Brains. Sci. Rep. Whales Res. Inst. 1948, 2, 1–20. [Google Scholar]
- Graaf, A.S.d. Anatomical Aspects of the Cetacean Brain Stem; Van Gorcum: Assen, The Netherlands, 1967. [Google Scholar]
- Zook, J.M.; DiCaprio, R.A. A Potential System of Delay-Lines in the Dolphin Auditory Brainstem. In Sensory Abilities of Cetaceans; Springer: Boston, MA, USA, 1990; pp. 181–193. [Google Scholar] [CrossRef]
- Malmierca, M.S. Anatomy and Physiology of the Mammalian Auditory System. In Encyclopedia of Computational Neuroscience; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1–36. [Google Scholar] [CrossRef]
- Morgane, P.J.; Jacobs, M.S.; McFarland, W.L. The anatomy of the brain of the bottlenose dolphin (Tursiops truncatus). Surface configurations of the telencephalon of the bottlenose dolphin with comparative anatomical observations in four other cetacean species. Brain Res. Bull. 1980, 5, 1–107. [Google Scholar] [CrossRef]
- Hinojosa, R.; Nelson, E.G. Cochlear nucleus neuron analysis in individuals with presbycusis. Laryngoscope 2011, 121, 2641–2648. [Google Scholar] [CrossRef]
- Cozzi, B.; Huggenberger, S.; Oelschläger, H. Anatomy of Dolphins: Insights into Body Structure and Function; Academic Press: Waretown, NJ, USA, 2016; ISBN 9780124072299. [Google Scholar]
- von Mikecz, A. Pathology and function of nuclear amyloid: Protein homeostasis matters. Nucleus 2014, 5, 311–317. [Google Scholar] [CrossRef]
- Hefter, D.; Draguhn, A. APP as a protective factor in acute neuronal insults. Front. Mol. Neurosci. 2017, 10, 22. [Google Scholar] [CrossRef]
- Ridgway, S.H. Diving by cetaceans. In Diving in Animals and Man; Brubakk, A.O., Kanwisher, J.W., Sundnes, G., Eds.; Royal Norwegian Society of Science and Letters: Trondheim, Norway, 1986; pp. 33–62. [Google Scholar]
- Klatsky, L.J.; Wells, R.S.; Sweeney, J.C. Offshore Bottlenose Dolphins (Tursiops truncatus): Movement and Dive Behavior Near the Bermuda Pedestal. J. Mammal. 2007, 88, 59–66. [Google Scholar] [CrossRef]
- Thal, D.R.; Rüb, U.; Orantes, M.; Braak, H. Phases of A beta-deposition in the human brain and its relevance for the development of AD. Neurology 2002, 58, 1791–1800. [Google Scholar] [CrossRef]
- Morris, G.P.; Clark, I.A.; Vissel, B. Questions concerning the role of amyloid-β in the definition, aetiology and diagnosis of Alzheimer’s disease. Acta Neuropathol. 2018, 136, 663–689. [Google Scholar] [CrossRef]
- Ulbrich, P.; Khoshneviszadeh, M.; Jandke, S.; Schreiber, S.; Dityatev, A. Interplay between perivascular and perineuronal extracellular matrix remodelling in neurological and psychiatric diseases. Eur. J. Neurosci. 2021, 53, 3811–3830. [Google Scholar] [CrossRef]
- Labrousse-Arias, D.; Martínez-Ruiz, A.; Calzada, M.J. Hypoxia and Redox Signaling on Extracellular Matrix Remodeling: From Mechanisms to Pathological Implications. Antioxid. Redox Signal. 2017, 27, 802–822. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Lee, Y.J.; Han, H.J. Role of hypoxia-induced fibronectin-integrin β1 expression in embryonic stem cell proliferation and migration: Involvement of PI3K/Akt and FAK. J. Cell. Physiol. 2011, 226, 484–493. [Google Scholar] [CrossRef] [PubMed]
- Michalski, D.; Spielvogel, E.; Puchta, J.; Reimann, W.; Barthel, H.; Nitzsche, B.; Mages, B.; Jäger, C.; Martens, H.; Horn, A.K.E.; et al. Increased Immunosignals of Collagen IV and Fibronectin Indicate Ischemic Consequences for the Neurovascular Matrix Adhesion Zone in Various Animal Models and Human Stroke Tissue. Front. Physiol. 2020, 11, 575598. [Google Scholar] [CrossRef] [PubMed]
- Edwards, D.N.; Bix, G.J. Roles of blood-brain barrier integrins and extracellular matrix in stroke. Am. J. Physiol. Cell Physiol. 2019, 316, C252–C263. [Google Scholar] [CrossRef]
- Lall, R.; Mohammed, R.; Ojha, U. What are the links between hypoxia and Alzheimer’s disease? Neuropsychiatr. Dis. Treat. 2019, 15, 1343–1354. [Google Scholar] [CrossRef]
- Schneuer, M.; Flachsbarth, S.; Czech-Damal, N.U.; Folkow, L.P.; Siebert, U.; Burmester, T. Neuroglobin of seals and whales: Evidence for a divergent role in the diving brain. Neuroscience 2012, 223, 35–44. [Google Scholar] [CrossRef]
- Gama Sosa, M.A.; De Gasperi, R.; Pryor, D.; Perez Garcia, G.S.; Perez, G.M.; Abutarboush, R.; Kawoos, U.; Hogg, S.; Ache, B.; Janssen, W.G.; et al. Low-level blast exposure induces chronic vascular remodeling, perivascular astrocytic degeneration and vascular-associated neuroinflammation. Acta Neuropathol. Commun. 2021, 9, 167. [Google Scholar] [CrossRef]
- Oyanagi, K.; Yamazaki, M.; Hashimoto, T.; Asakawa, M.; Wakabayashi, K.; Takahashi, H. Hippocampal sclerosis in the parkinsonism-dementia complex of Guam: Quantitative examination of neurons, neurofibrillary tangles, and TDP-43 immunoreactivity in CA1. Neuropathology 2015, 35, 224–235. [Google Scholar] [CrossRef]
- Neumann, M.; Sampathu, D.M.; Kwong, L.K.; Truax, A.C.; Micsenyi, M.C.; Chou, T.T.; Bruce, J.; Schuck, T.; Grossman, M.; Clark, C.M.; et al. Ubiquitinated TDP-43 in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Science 2006, 314, 130–133. [Google Scholar] [CrossRef]
- Atkinson, R.; Leung, J.; Bender, J.; Kirkcaldie, M.; Vickers, J.; King, A. TDP-43 mislocalization drives neurofilament changes in a novel model of TDP-43 proteinopathy. Dis. Model. Mech. 2021, 14, dmm047548. [Google Scholar] [CrossRef]
- Hof, P.R.; Glezer, I.I.; Archin, N.; Janssen, W.G.; Morgane, P.J.; Morrison, J.H. The primary auditory cortex in cetacean and human brain: A comparative analysis of neurofilament protein-containing pyramidal neurons. Neurosci. Lett. 1992, 146, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Morell, M.; Raverty, S.A.; Mulsow, J.; Haulena, M.; Barrett-Lennard, L.; Nordstrom, C.A.; Venail, F.; Shadwick, R.E. Combining Cochlear Analysis and Auditory Evoked Potentials in a Beluga Whale With High-Frequency Hearing Loss. Front. Vet. Sci. 2020, 7, 534917. [Google Scholar] [CrossRef] [PubMed]
- Ulfig, N.; Nickel, J.; Bohl, J. Monoclonal antibodies SMI 311 and SMI 312 as tools to investigate the maturation of nerve cells and axonal patterns in human fetal brain. Cell Tissue Res. 1998, 291, 433–443. [Google Scholar] [CrossRef]
- Redondo, J.; Hares, K.; Wilkins, A.; Scolding, N.; Kemp, K. Reductions in kinesin expression are associated with nitric oxide-induced axonal damage. J. Neurosci. Res. 2015, 93, 882–892. [Google Scholar] [CrossRef]
- Schirmer, L.; Antel, J.P.; Brück, W.; Stadelmann, C. Axonal Loss and Neurofilament Phosphorylation Changes Accompany Lesion Development and Clinical Progression in Multiple Sclerosis. Brain Pathol. 2011, 21, 428. [Google Scholar] [CrossRef]
- Nasuhidehnavi, A.; Yap, G.S. Microglia and astrocyte responses to neuropathogenic protozoan parasites. Fac. Rev. 2021, 10, 69. [Google Scholar] [CrossRef]
- Pintore, M.D.; Mignone, W.; Di Guardo, G.; Mazzariol, S.; Ballardini, M.; Florio, C.L.; Goria, M.; Romano, A.; Caracappa, S.; Giorda, F.; et al. Neuropathologic findings in cetaceans stranded in Italy (2002–14). J. Wildl. Dis. 2018, 54, 295–303. [Google Scholar] [CrossRef]
- Yin, H.Z.; Yu, S.; Hsu, C.I.; Liu, J.; Acab, A.; Wu, R.; Tao, A.; Chiang, B.J.; Weiss, J.H. Intrathecal infusion of BMAA induces selective motor neuron damage and astrogliosis in the ventral horn of the spinal cord. Exp. Neurol. 2014, 261, 1–9. [Google Scholar] [CrossRef]
- Finger, C.E.; Moreno-Gonzalez, I.; Gutierrez, A.; Moruno-Manchon, J.F.; McCullough, L.D. Age-related immune alterations and cerebrovascular inflammation. Mol. Psychiatry 2021, 27, 803–818. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, L.S.; Zinsmaier, A.K.; Patterson, G.; Leptich, E.J.; Shoemaker, S.L.; Yatskievych, T.A.; Gibboni, R.; Pace, E.; Luo, H.; et al. Neuroinflammation mediates noise-induced synaptic imbalance and tinnitus in rodent models. PLoS Biol. 2019, 17, e3000307. [Google Scholar] [CrossRef]
- Chang, K.-H.; Jin, S.-Y.; Park, J.-W.; Park, G.-E.; Lee, K.-A.; Jun, B.-C.; Yeo, S.-W. Expression of Glial Fibrillay Acidic Protein in the Central Auditory Pathway of Rats after Loud Noise Exposure. Korean J. Audiol. 2010, 14, 12–15. [Google Scholar]
- Säljö, A.; Bao, F.; Hamberger, A.; Haglid, K.G.; Hansson, H.A. Exposure to short-lasting impulse noise causes microglial and astroglial cell activation in the adult rat brain. Pathophysiology 2001, 8, 105–111. [Google Scholar] [CrossRef]
- Ries, M.; Sastre, M. Mechanisms of Aβ Clearance and Degradation by Glial Cells. Front. Aging Neurosci. 2016, 8, 160. [Google Scholar] [CrossRef]
- Gerrits, E.; Brouwer, N.; Kooistra, S.M.; Woodbury, M.E.; Vermeiren, Y.; Lambourne, M.; Mulder, J.; Kummer, M.; Möller, T.; Biber, K.; et al. Distinct amyloid-β and tau-associated microglia profiles in Alzheimer’s disease. Acta Neuropathol. 2021, 141, 681–696. [Google Scholar] [CrossRef]
- Tom, V.J.; Doller, C.M.; Malouf, A.T.; Silver, J. Astrocyte-Associated Fibronectin Is Critical for Axonal Regeneration in Adult White Matter. J. Neurosci. 2004, 24, 9282. [Google Scholar] [CrossRef]
- Marino, L.; Connor, R.C.; Fordyce, R.E.; Herman, L.M.; Hof, P.R.; Lefebvre, L.; Lusseau, D.; McCowan, B.; Nimchinsky, E.A.; Pack, A.A.; et al. Cetaceans have complex brains for complex cognition. PLoS Biol. 2007, 5, 0966–0972. [Google Scholar] [CrossRef]
- Reiss, D.; Marino, L. Mirror self-recognition in the bottlenose dolphin: A case of cognitive convergence. Proc. Natl. Acad. Sci. USA 2001, 98, 5937–5942. [Google Scholar] [CrossRef]
- Smith, T.G.; Siniff, D.B.; Reichle, R.; Stone, S. Coordinated behavior of killer whales, Orcinus orca, hunting a crabeater seal, Lobodon carcinophagus. Can. J. Zool. 1981, 59, 1185–1189. [Google Scholar] [CrossRef]
- Morlock, G.E.; Ziltener, A.; Geyer, S.; Tersteegen, J.; Mehl, A.; Schreiner, T.; Kamel, T.; Brümmer, F. Evidence that Indo-Pacific bottlenose dolphins self-medicate with invertebrates in coral reefs. iScience 2022, 25, 104271. [Google Scholar] [CrossRef]
- Hall, J.G. Hearing and Primary Auditory Centres of the Whales. Acta Oto-Laryngologica 2009, 63, 244–250. [Google Scholar] [CrossRef]
- Bullock, T.H.; Gurevich, V.S. Soviet literature on the nervous system and psychobiology of Cetacea. Int. Rev. Neurobiol. 1979, 21, 47–127. [Google Scholar] [CrossRef] [PubMed]
- Jansen, J.; Jansen, J.K.S. The nervous system of Cetacea. In Biology of Marine Mammals; Andersen, H.T., Ed.; Academic Press: New York, NY, USA, 1969; pp. 175–252. [Google Scholar]
- Gandolfi, A.; Horoupian, D.S.; De Teresa, R.M. Quantitative and cytometric analysis of the ventral cochlear nucleus in man. J. Neurol. Sci. 1981, 50, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Huggenberger, S.; Oelschläger, H.A.; Cozzi, B.; Centelleghe, C.; Banzato, T.; Demma, M.; Oelschläger, J.; Gorter, U. Atlas of the Anatomy of Dolphins and Whales/Stefan Huggenberger, Helmut Oelschläger, Bruno Cozzi; in Collaboration with Cinzia Centelleghe [and 3 Others]; with the Assistance of Tommaso Banzato [and 8 Others]; Massimo, D., Jutta, O., Eds.; Academic Press: Waretown, NJ, USA, 2019. [Google Scholar]
- Sitek, K.R.; Gulban, O.F.; Calabrese, E.; Johnson, G.A.; Lage-Castellanos, A.; Moerel, M.; Ghosh, S.S.; De Martino, F. Mapping the human subcortical auditory system using histology, postmortem MRI and in vivo MRI at 7T. Elife 2019, 8, e48932. [Google Scholar] [CrossRef] [PubMed]
- Sitek, K.R.; Calabrese, E.; Johnson, G.A.; Ghosh, S.S.; Chandrasekaran, B. Structural Connectivity of Human Inferior Colliculus Subdivisions Using in vivo and post mortem Diffusion MRI Tractography. Front. Neurosci. 2022, 16, 751595. [Google Scholar] [CrossRef]
- Malkemper, E.P.; Oelschläger, H.H.A.; Huggenberger, S. The dolphin cochlear nucleus: Topography, histology and functional implications. J. Morphol. 2012, 273, 173–185. [Google Scholar] [CrossRef]
- Ijsseldijk, L.L.; Brownlow, A.C.; Mazzariol, S. Best Practice on Cetacean Post Mortem Investigation and Tissue Sampling; Joint ACCOBAMS and ASCOBANS document. 2019. Available online: osf.io/zh4ra/ (accessed on 15 April 2022).
- Obusan, M.C.M.; Caras, J.A.A.; Lumang, L.S.L.; Calderon, E.J.S.; Villanueva, R.M.D.; Salibay, C.C.; Siringan, M.A.T.; Rivera, W.L.; Masangkay, J.S.; Aragones, L.V. Bacteriological and histopathological findings in cetaceans that stranded in the Philippines from 2017 to 2018. bioRxiv 2020. [Google Scholar] [CrossRef]
- Morell, M.; Brownlow, A.; McGovern, B.; Raverty, S.A.; Shadwick, R.E.; André, M. Implementation of a method to visualize noise-induced hearing loss in mass stranded cetaceans. Sci. Rep. 2017, 7, srep41848. [Google Scholar] [CrossRef]












| Antibody | Host | Reactivity | Clonality | RRID | Catalogue Number | Dilution IHC | Dilution WB |
|---|---|---|---|---|---|---|---|
| Aβ | rabbit | rat, human, mouse, bottlenose dolphin | monoclonal | AB_2532306 | 700254 | 1:500 | 1:500 1 |
| fibronectin | rabbit | human | polyclonal | AB_476976 | F3648 | 1:200 | 1:1000 |
| TDP-43 | rat | human, Rat | monoclonal | AB_2750118 | 829901 | 1:200 | 1:1000 |
| SMI-312 | mouse | human, mouse, rat | monoclonal | AB_2566782 | 837904 | 1:1000 | 1:1000 |
| Iba-1 | rabbit | human, mouse, rat, other | polyclonal | AB_839504 | 019-1974 | 1:500 | NA |
| GFAP | rabbit | cat, cow, dog, human, mouse, rat, sheep | polyclonal | AB_10013382 | Z0334 | 1:500 | 1:1000 |
| Specimen | Auditory Nucleus | Volume MRI (mm3) | Volume Cavalieri (mm3) |
|---|---|---|---|
| Marine 0142 | LIC | 1775 | 1362 |
| LVCN | 435 | 129 * | |
| Marine 0116 | LIC | 1345 | - |
| LVCN | 387 | - | |
| RIC | 1451 | 921 | |
| RVCN | 378 | 285 | |
| Dolphin 2 ** | RIC | - | 571 |
| LVCN | - | 199 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orekhova, K.; Selmanovic, E.; De Gasperi, R.; Gama Sosa, M.A.; Wicinski, B.; Maloney, B.; Seifert, A.; Alipour, A.; Balchandani, P.; Gerussi, T.; et al. Multimodal Assessment of Bottlenose Dolphin Auditory Nuclei Using 7-Tesla MRI, Immunohistochemistry and Stereology. Vet. Sci. 2022, 9, 692. https://doi.org/10.3390/vetsci9120692
Orekhova K, Selmanovic E, De Gasperi R, Gama Sosa MA, Wicinski B, Maloney B, Seifert A, Alipour A, Balchandani P, Gerussi T, et al. Multimodal Assessment of Bottlenose Dolphin Auditory Nuclei Using 7-Tesla MRI, Immunohistochemistry and Stereology. Veterinary Sciences. 2022; 9(12):692. https://doi.org/10.3390/vetsci9120692
Chicago/Turabian StyleOrekhova, Ksenia, Enna Selmanovic, Rita De Gasperi, Miguel A. Gama Sosa, Bridget Wicinski, Brigid Maloney, Alan Seifert, Akbar Alipour, Priti Balchandani, Tommaso Gerussi, and et al. 2022. "Multimodal Assessment of Bottlenose Dolphin Auditory Nuclei Using 7-Tesla MRI, Immunohistochemistry and Stereology" Veterinary Sciences 9, no. 12: 692. https://doi.org/10.3390/vetsci9120692
APA StyleOrekhova, K., Selmanovic, E., De Gasperi, R., Gama Sosa, M. A., Wicinski, B., Maloney, B., Seifert, A., Alipour, A., Balchandani, P., Gerussi, T., Graïc, J.-M., Centelleghe, C., Di Guardo, G., Mazzariol, S., & Hof, P. R. (2022). Multimodal Assessment of Bottlenose Dolphin Auditory Nuclei Using 7-Tesla MRI, Immunohistochemistry and Stereology. Veterinary Sciences, 9(12), 692. https://doi.org/10.3390/vetsci9120692