
Protective Effects of Water Extract of Fructus Ligustri Lucidi against Oxidative Stress-Related Osteoporosis In Vivo and In Vitro

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Preparation of Fructus Ligustri Lucidi
2.3. Animals and Animal Ethics
2.4. In Vivo Studies
2.4.1. Experimental Design
2.4.2. Sample Collection
2.4.3. Biochemical Analysis of Serum, Urine, and Fecal Samples
2.4.4. Measurement of Superoxide Dismutase (SOD) and Malondialdehyde (MDA) Level of Tissues in Mice
2.4.5. Analysis of Bone Index and Bone Dry-Wet Ratio
2.4.6. Micro-Computed Tomography Analysis of Mice Femurs
2.5. In Vitro Studies
2.5.1. Cell Culture
2.5.2. Cell Viability Assay
2.5.3. Detection of ALP level in Cell Supernatant
2.5.4. Expression of OPG and RANKL Protein Study
2.6. Statistical Analysis
3. Results
3.1. Effects of FLL on Body Weight, Serum and Urinary Biochemical, and Fecal Ca Study in Mice
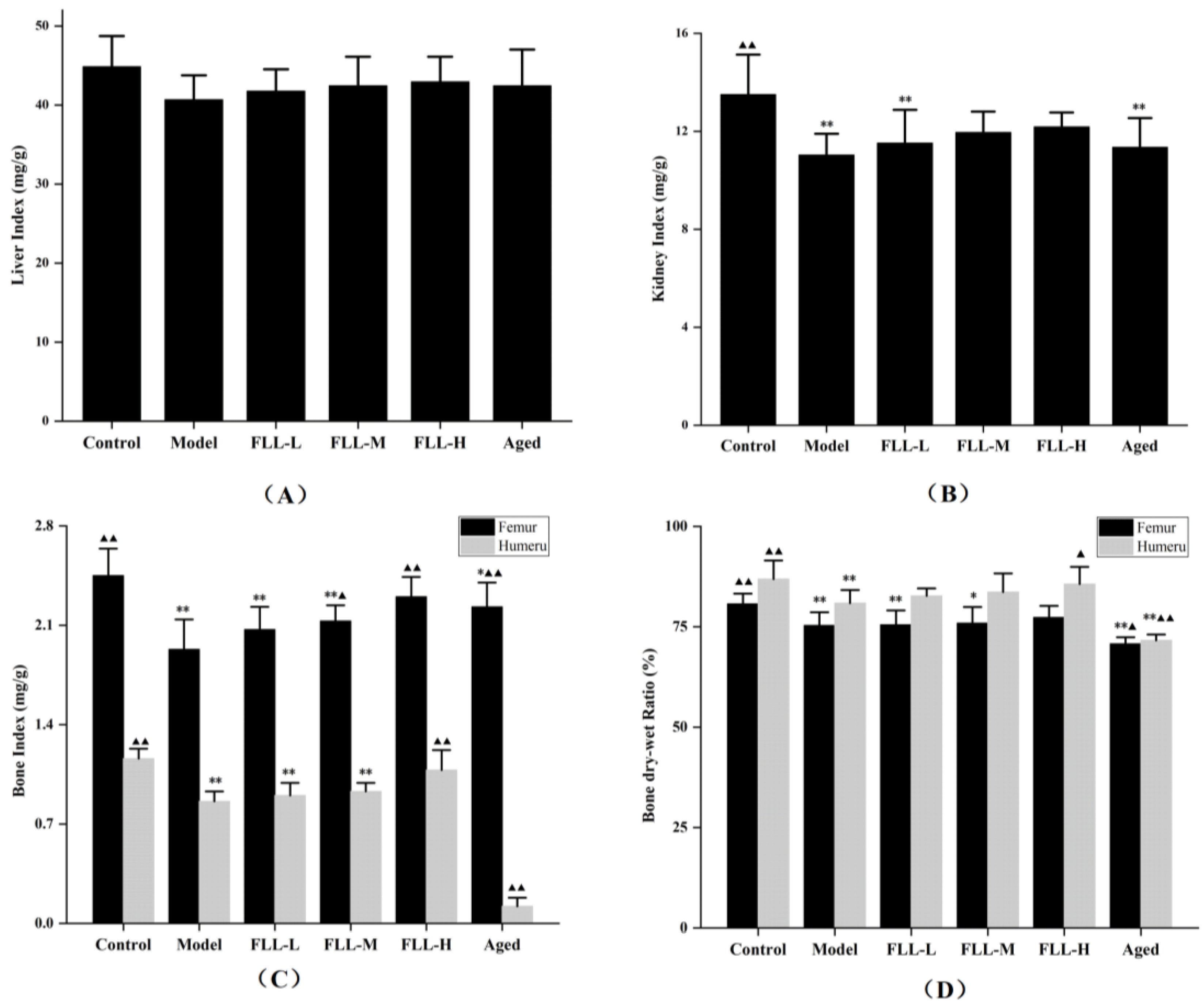
3.2. Effects of FLL on Organ Index, Bone Index, and Bone Dry–Wet Ratio
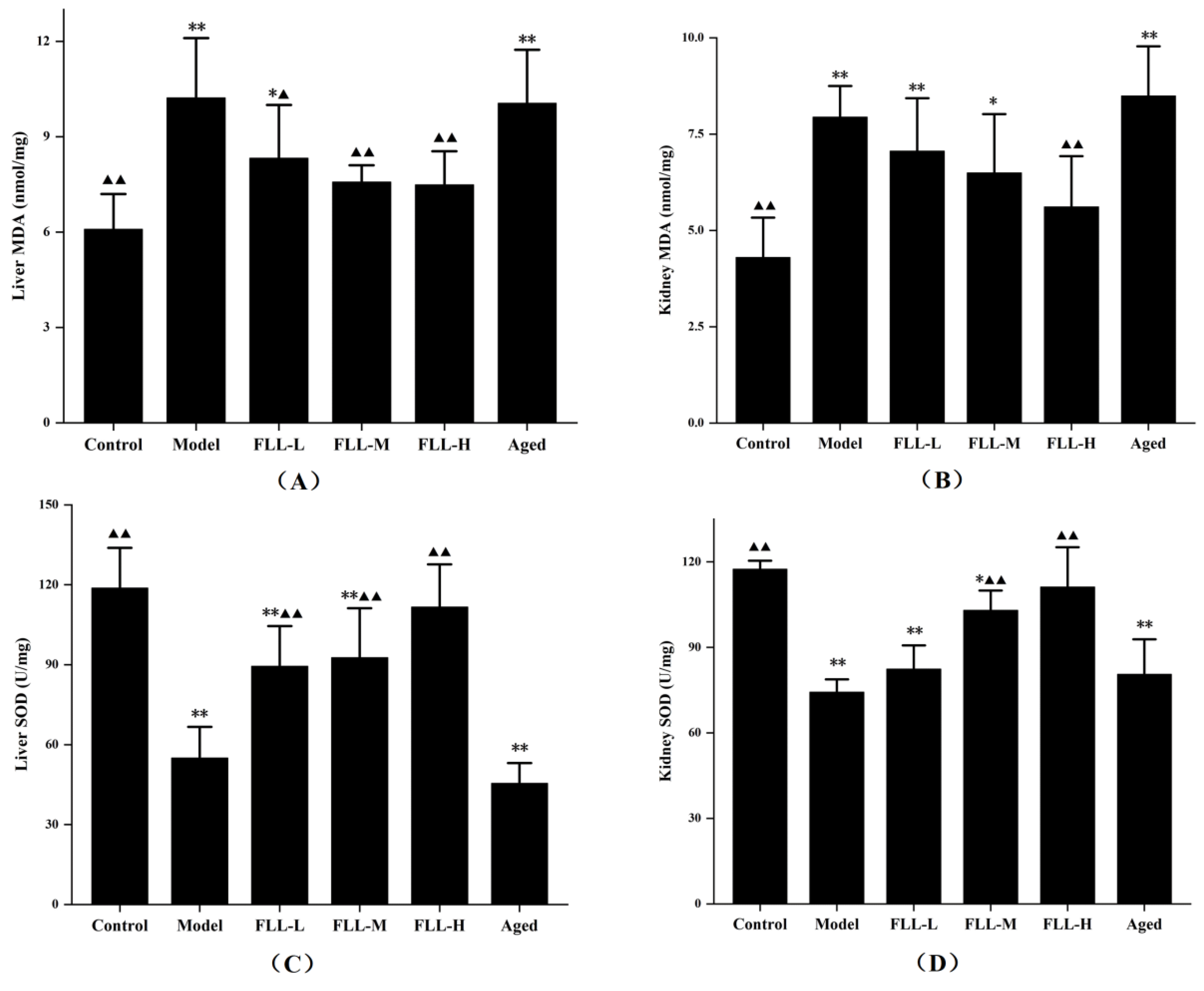
3.3. Effects of FLL on SOD Activity and MDA Content in the Liver and Kidney of Mice
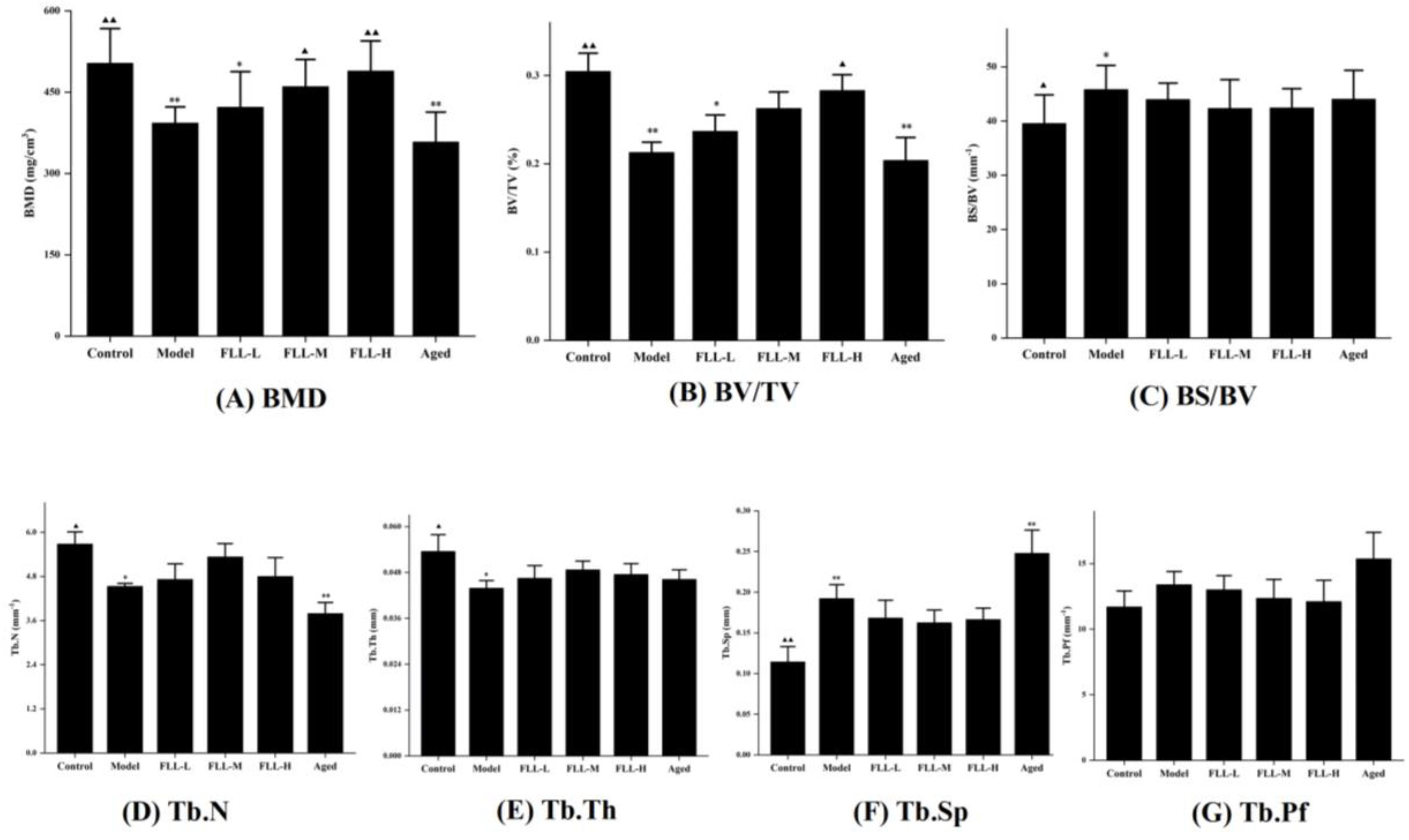
3.4. Effects of FLL on Bone Properties in Mice Analyzed by Micro-CT
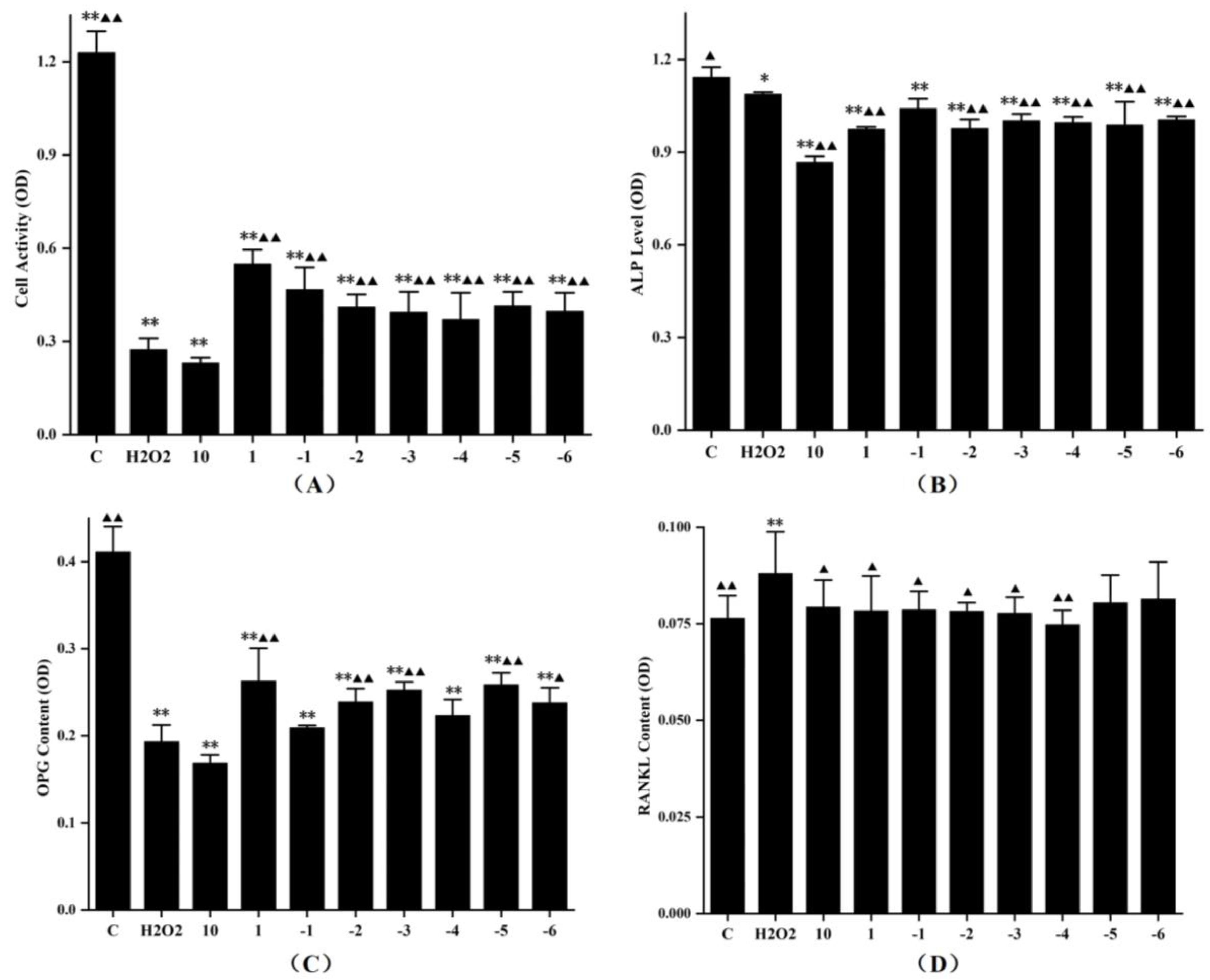
3.5. Effects of FLL on Proliferation, Differentiation and Expression of OPG and RANKL Protein in Osteoblasts under Oxidative Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Compston, J.E.; McClung, M.R.; Leslie, W.D. Osteoporosis. Lancet 2019, 393, 364–376. [Google Scholar] [CrossRef]
- Manolagas, S.C. From estrogen-centric to aging and oxidative stress: Revised perspective of the pathogenesis of osteoporosis. Endocr. Rev. 2010, 31, 266–300. [Google Scholar] [CrossRef] [Green Version]
- Cornelius, C.; Koverech, G.; Crupi, R.; di Paola, R.; Koverech, A.; Lodato, F.; Scuto, M.; Salinaro, A.T.; Cuzzocrea, S.; Calabrese, E.J.; et al. Osteoporosis and alzheimer pathology: Role of cellular stress response and hormetic redox signaling in aging and bone remodeling. Front. Pharmacol. 2014, 5, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Li, B.; Shi, Y.; Wang, C.L.; Ye, L. Targeting reactive oxygen species in stem cells for bone therapy. Drug Discov. Today 2021, 26, 1226–1244. [Google Scholar] [CrossRef]
- Kanzaki, H.; Shinohara, F.; Kajiya, M.; Kodama, T. The Keap1/Nrf2 protein axis plays a role in osteoclast differentiation by regulating intracellular reactive oxygen species signaling. J. Biol. Chem. 2013, 288, 23009–230020. [Google Scholar] [CrossRef] [Green Version]
- Callaway, D.A.; Jiang, J.X. Reactive oxygen species and oxidative stress in osteoclastogenesis, skeletal aging and bone diseases. J. Bone Miner. Metab. 2015, 33, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Veldurthy, V.; Wei, R.; Oz, L.; Dhawan, P.; Jeon, Y.H.; Christakos, S. Vitamin D, calcium homeostasis and aging. Bone Res. 2016, 4, 16041. [Google Scholar] [CrossRef] [Green Version]
- Khosla, S.; Hofbauer, L.C. Osteoporosis treatment: Recent developments and ongoing challenges. Lancet Diabetes Endocrinol. 2017, 5, 898–907. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Huang, G.; Chen, G. Synthesis and biological activities of drugs for the treatment of osteoporosis. Eur. J. Med. Chem. 2020, 197, 112313. [Google Scholar] [CrossRef]
- Rossini, M.; Adami, G.; Adami, S.; Viapiana, O.; Gatti, D. Safety issues and adverse reactions with osteoporosis management. Expert Opin. Drug Saf. 2016, 15, 321–332. [Google Scholar] [CrossRef]
- McClung, M.; Harris, S.T.; Miller, P.D.; Bauer, D.C.; Davison, K.S.; Dian, L.; Hanley, D.A.; Kendler, D.L.; Yuen, C.K.; Lewiecki, E.M. Bisphosphonate Therapy for Osteoporosis: Benefits, Risks, and Drug Holiday. Am. J. Med. 2013, 126, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Cummings, S.R.; Schwartz, A.V.; Black, D.M. Alendronate and atrial fibrillation. N. Engl. J. Med. 2007, 356, 1895–1896. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Shi, J.; Gao, B.; Zhang, H.-Y.; Fan, J.; Li, X.-J.; Fan, J.-Z.; Han, Y.-H.; Zhang, J.-K.; Yang, L.; et al. Gastrodin: An ancient Chinese herbal medicine as a source for antiosteoporosis agents via reducing reactive oxygen species. Bone 2015, 73, 132–144. [Google Scholar] [CrossRef]
- He, J.B.; Chen, M.H.; Lin, D.K. New insights into the tonifying kidney-yin herbs and formulas for the treatment of osteoporosis. Arch. Osteoporos. 2017, 12, 14. [Google Scholar] [CrossRef]
- Gao, L.; Li, C.; Wang, Z.; Liu, X.; You, Y.; Wei, H.; Guo, T. Ligustri Lucidi Fructus as a traditional Chinese medicine: A review of its phytochemistry and pharmacology. Nat. Prod. Res. 2014, 29, 493–510. [Google Scholar] [CrossRef]
- Chen, B.; Wang, L.; Li, L.; Zhu, R.; Liu, H.; Liu, C.; Ma, R.; Jia, Q.; Zhao, D.; Niu, J.; et al. Fructus Ligustri Lucidi in Osteoporosis: A Review of its Pharmacology, Phytochemistry, Pharmacokinetics and Safety. Molecules 2017, 22, 1469. [Google Scholar] [CrossRef]
- Zhang, W.; Feng, J.; Cheng, B.; Lu, Q.; Chen, X.P. Oleanolic acid protects against oxidative stress-induced human umbilical vein endothelial cell injury by activating AKT/eNOS signaling. Mol. Med. Rep. 2018, 18, 3641–3648. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Fan, Y.-S.; Gao, Z.-Q.; Fan, K.; Liu, Z.-J. Effect of Fructus Ligustri Lucidi on osteoblastic like cell-line MC3T3-E1. J. Ethnopharmacol. 2015, 170, 88–95. [Google Scholar] [CrossRef]
- Dong, X.-L.; Zhang, Y.; Favus, M.J.; Che, C.-T.; Wong, M.-S. Ethanol extract of Fructus Ligustri Lucidi increases circulating 1,25-dihydroxyvitamin D3 by inducing renal 25-hydroxyvitamin D-1α hydroxylase activity. Menopause 2010, 17, 1174–1181. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Dong, X.-L.; Leung, P.-C.; Che, C.-T.; Wong, M.-S. Fructus ligustri lucidi extract improves calcium balance and modulates the calciotropic hormone level and vitamin D-dependent gene expression in aged ovariectomized rats. Menopause 2008, 15, 558–565. [Google Scholar] [CrossRef] [PubMed]
- Giorgio, M.; Trinei, M.; Migliaccio, E.; Pelicci, P.G. Hydrogen peroxide: A metabolic by-product or a common mediator of ageing signals. Nat. Rev. Mol. Cell Biol. 2007, 8, 722–728. [Google Scholar] [CrossRef]
- Ren, Y.; Yang, X.; Niu, X.; Liu, S.; Ren, G. Chemical Characterization of the Avenanthramide-Rich Extract from Oat and Its Effect on d-Galactose-Induced Oxidative Stress in Mice. J. Agric. Food Chem. 2011, 59, 206–211. [Google Scholar] [CrossRef]
- Zhang, D.; Han, J.; Li, Y.; Yuan, B.; Zhou, J.; Cheong, L.-Z.; Li, Y.; Lu, C.; Su, X. Tuna Oil Alleviates d-Galactose Induced Aging in Mice Accompanied by Modulating Gut Microbiota and Brain Protein Expression. J. Agric. Food Chem. 2018, 66, 5510–5520. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chen, D.; Wang, Z.K.; Chen, C.Y.; Ning, D.L.; Zhao, S.L. Protective effect of walnut on d-galactose-induced aging mouse model. Food Sci. Nutr. 2019, 7, 969–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casati, L.; Pagani, F.; Limonta, P.; Vanetti, C.; Stancari, G.; Sibilia, V. Beneficial effects of δ-tocotrienol against oxidative stress in osteoblastic cells: Studies on the mechanisms of action. Eur. J. Nutr. 2020, 59, 1975–1987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oftadeh, R.; Perez-Viloria, M.; Villa-Camacho, J.C.; Vaziri, A.; Nazarian, A. Biomechanics and Mechanobiology of Trabecular Bone: A Review. J. Biomech. Eng. 2015, 137, 010802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dempster, D.W.; Compston, J.E.; Drezner, M.K.; Glorieux, F.H.; Kanis, J.A.; Malluche, H.; Meunier, P.J.; Ott, S.M.; Recker, R.R.; Parfitt, A.M. Standardized nomenclature, symbols, and units for bone histomorphometry: A 2012 update of the report of the ASBMR Histomorphometry Nomenclature Committee. J. Bone Miner. Res. 2012, 28, 2–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prentice, A.; Bonjour, J.-P.; Branca, F.; Cooper, C.; Flynn, A.; Pannemans, D.; Weber, P. PASSCLAIM–Bone health and osteoporosis. Eur. J. Nutr. 2003, 42, i28–i49. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Hendriks, M.; Chatzis, A.; Ramasamy, S.K.; Kusumbe, A.P. Bone Vasculature and Bone Marrow Vascular Niches in Health and Disease. J. Bone Miner. Res. 2020, 35, 2103–2120. [Google Scholar] [CrossRef]
- Putnam, S.E.; Scutt, A.M.; Bicknell, K.; Priestley, C.M.; Williamson, E.M. Natural products as alternative treatments for metabolic bone disorders and for maintenance of bone health. Phytother. Res. 2006, 21, 99–102. [Google Scholar] [CrossRef]
- Aizenberg, J.; Weaver, J.C.; Thanawala, M.S.; Sundar, V.C.; Morse, D.E.; Fratzl, P. Skeleton of Euplectella sp.: Structural Hierarchy from the Nanoscale to the Macroscale. Science 2005, 309, 275–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, W.C.; Guntur, A.R.; Long, F.X.; Rosen, C.J. Energy Metabolism of the Osteoblast: Implications for Osteoporosis. Endocr. Rev. 2017, 38, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Lv, C.; Hao, Y.; Han, Y.; Zhang, W.; Cong, L.; Shi, Y.; Tu, G. Role and mechanism of microRNA-21 in H2O2-induced apoptosis in bone marrow mesenchymal stem cells. J. Clin. Neurosci. 2016, 27, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Sims, N.A.; Vrahnas, C. Regulation of cortical and trabecular bone mass by communication between osteoblasts, osteocytes and osteoclasts. Arch. Biochem. Biophys. 2014, 561, 22–28. [Google Scholar] [CrossRef]
- Wang, P.; Cao, Y.; Zhan, D.; Wang, D.; Wang, B.; Liu, Y.; Li, G.; He, W.; Wang, H.; Xu, L. Influence of DNA methylation on the expression of OPG/RANKL in primary osteoporosis. Int. J. Med. Sci. 2018, 15, 1480–1485. [Google Scholar] [CrossRef] [Green Version]
- Sunyecz, J.A.; Weisman, S.M. The role of calcium in osteoporosis drug therapy. J. Womens Health 2005, 2, 180–192. [Google Scholar] [CrossRef] [PubMed]
- Girón-Prieto, M.S.; Cano-García, M.D.C.; Poyatos-Andújar, A.; Arias-Santiago, S.; De Haro-Muñoz, T.; Arrabal-Martín, M.; Arrabal-Polo, M.A. The value of hypercalciuria in patients with osteopenia versus osteoporosis. Urolithiasis 2016, 45, 279–283. [Google Scholar] [CrossRef]
- Iolascon, G.; Frizzi, L.; Di Pietro, G.; Capaldo, A.; Luciano, F.; Gimigliano, F. Bone quality and bone strength: Benefits of the bone-forming approach. Clin. Cases Miner. Bone Metab. 2014, 11, 20–24. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Model | FLL-L | FLL-M | FLL-H | Aged | |
|---|---|---|---|---|---|---|
| Body Weight Change | ||||||
| Weight change rate, (%) | 3.41 ± 2.84 ▲▲ | 9.34 ± 5.38 ** | 8.82 ± 2.16 ** | 6.03 ± 3.36 | 4.10 ± 2.23 ▲ | 1.63 ± 1.20 ▲▲ |
| Serum and Urine Biochemical | ||||||
| S-Ca, mg/dL | 9.81 ± 0.87 | 10.51 ± 0.56 | 10.20 ± 0.90 | 10.09 ± 0.61 | 10.34 ± 0.94 | 9.80 ± 1.01 |
| U-Ca, mg/dL | 7.29 ± 0.90 ▲ | 8.73 ± 0.80 * | 8.50 ± 1.25 * | 8.03 ± 1.01 | 7.69 ± 0.98 | 6.64 ± 1.28 ▲▲ |
| S-Ca/Cr, mg/mg | 11.62 ± 2.95 | 13.92 ± 3.71 | 13.18 ± 4.02 | 12.71 ± 2.39 | 12.20 ± 1.15 | 13.71 ± 2.71 |
| U-Ca/Cr, mg/mg | 0.0936 ± 0.0194 ▲ | 0.1307 ± 0.0375 * | 0.1196 ± 0.0359 | 0.1134 ± 0.0369 | 0.0961 ± 0.0126 ▲ | 0.1382 ± 0.0319 ** |
| Fecal Calcium Level | ||||||
| Fecal Ca, mg/g | 4.77 ± 1.92 ▲▲ | 13.49 ± 1.68 ** | 7.85 ± 2.37 *▲▲ | 5.91 ± 2.41 ▲▲ | 4.61 ± 2.58 ▲▲ | 7.23 ± 2.45 ▲▲ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Hu, Y.; Zhao, Z.; Xu, L.; Chen, Y.; Liu, T.; Li, Q. Protective Effects of Water Extract of Fructus Ligustri Lucidi against Oxidative Stress-Related Osteoporosis In Vivo and In Vitro. Vet. Sci. 2021, 8, 198. https://doi.org/10.3390/vetsci8090198
Wu Y, Hu Y, Zhao Z, Xu L, Chen Y, Liu T, Li Q. Protective Effects of Water Extract of Fructus Ligustri Lucidi against Oxidative Stress-Related Osteoporosis In Vivo and In Vitro. Veterinary Sciences. 2021; 8(9):198. https://doi.org/10.3390/vetsci8090198
Chicago/Turabian StyleWu, Yi, Yusheng Hu, Zeguang Zhao, Lina Xu, Ye Chen, Tongtong Liu, and Qin Li. 2021. "Protective Effects of Water Extract of Fructus Ligustri Lucidi against Oxidative Stress-Related Osteoporosis In Vivo and In Vitro" Veterinary Sciences 8, no. 9: 198. https://doi.org/10.3390/vetsci8090198
APA StyleWu, Y., Hu, Y., Zhao, Z., Xu, L., Chen, Y., Liu, T., & Li, Q. (2021). Protective Effects of Water Extract of Fructus Ligustri Lucidi against Oxidative Stress-Related Osteoporosis In Vivo and In Vitro. Veterinary Sciences, 8(9), 198. https://doi.org/10.3390/vetsci8090198