
Renal Senescence, Telomere Shortening and Nitrosative Stress in Feline Chronic Kidney Disease

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Case Selection
2.2. Histologic Scoring
2.3. Immunohistochemistry
2.4. Assessment of p16 and iNOS IHC
2.5. Telomere FISH Slide Preparation
2.6. FISH Image Capture, Processing and Analysis
2.7. Statistical Analysis
3. Results
3.1. Histopathology
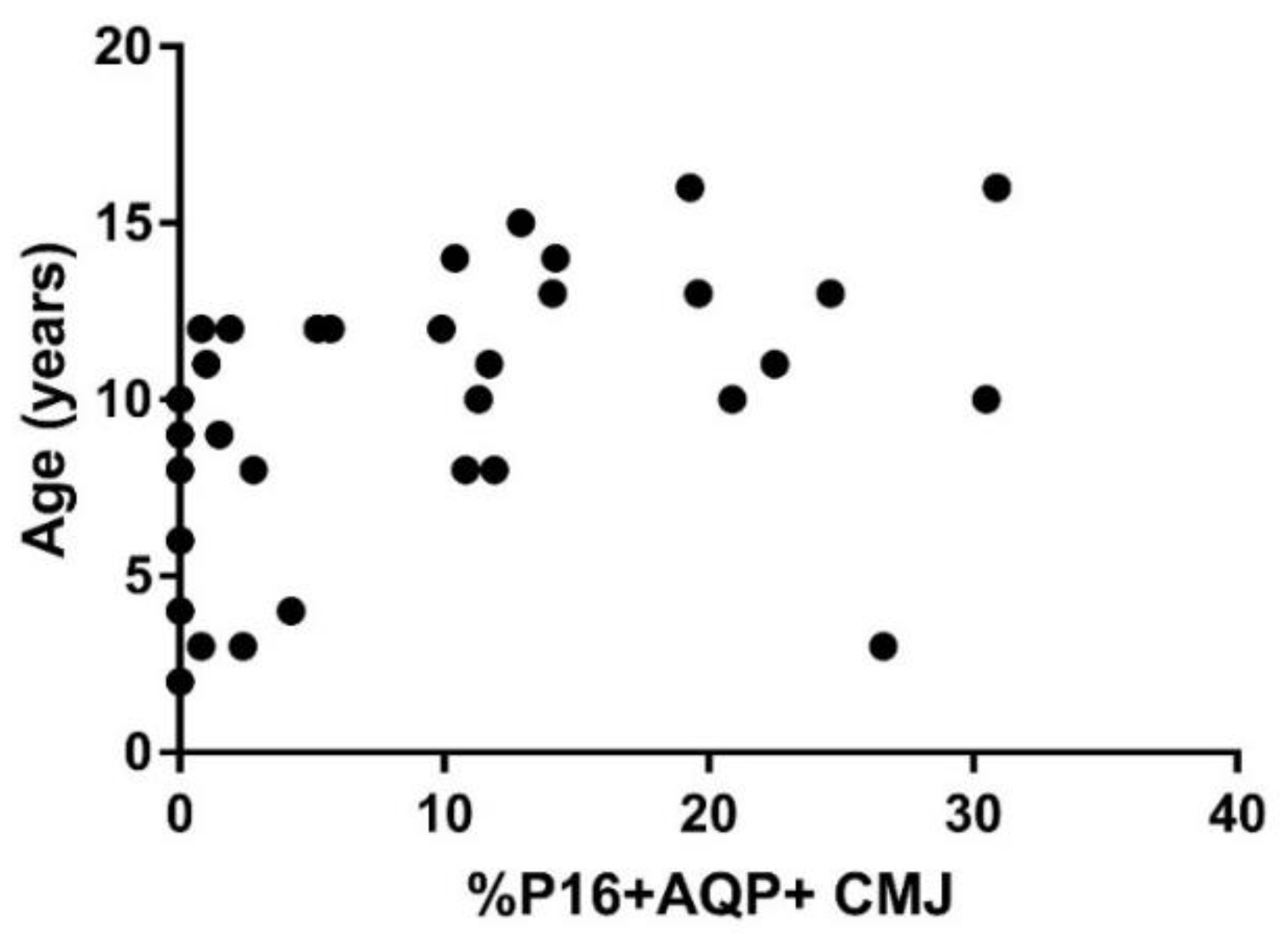
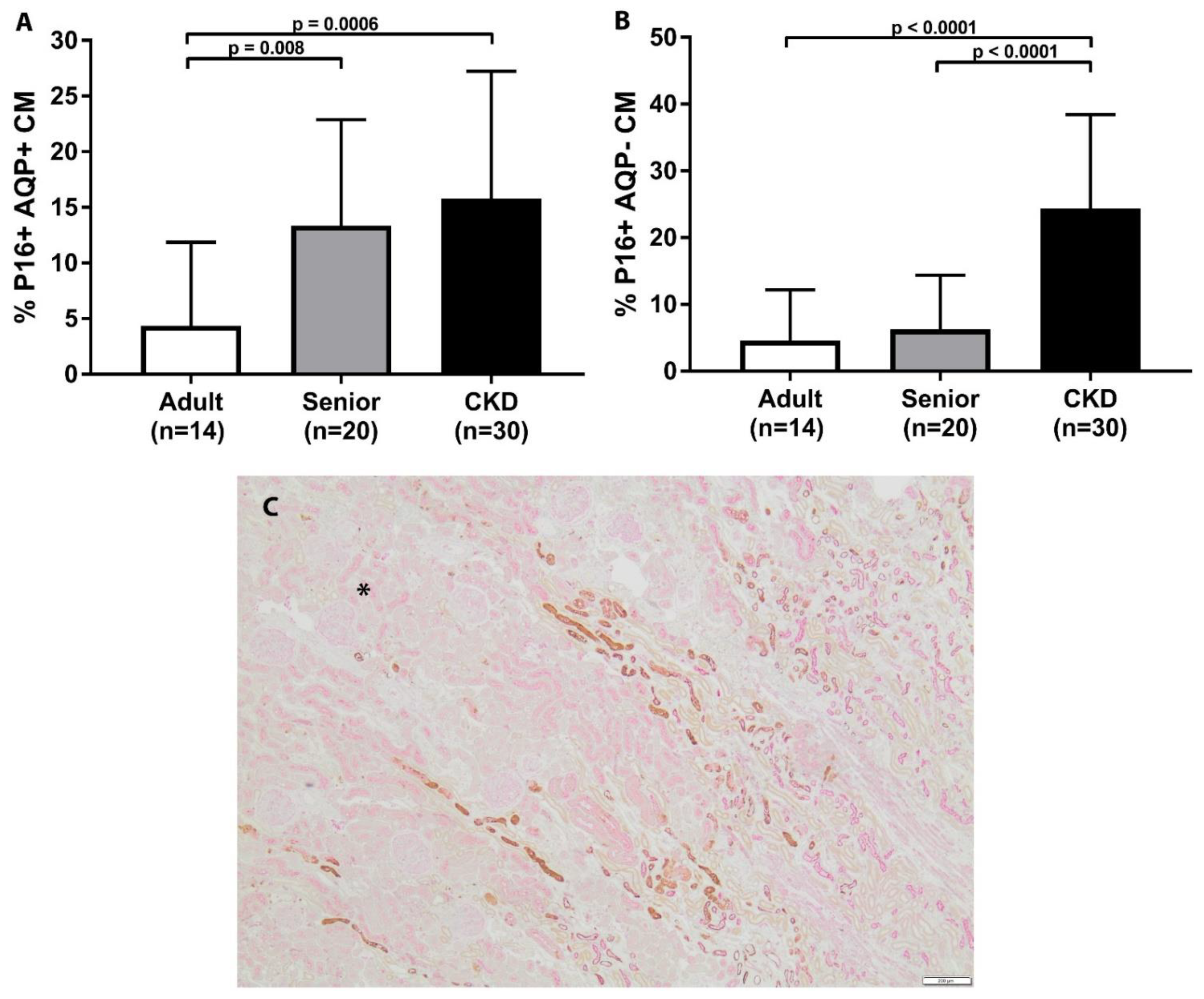
3.2. Cellular Senescence Is Associated with Age and Increased in CKD and Senior Cat Kidneys
3.3. Telomeres Are Significantly Shortened in Kidneys of CKD Cats
3.4. Nitrosative Stress Is Increased in Feline CKD
3.5. Glomerulosclerosis, Inflammation, and Fibrosis Is Correlated with p16 Staining but Not Telomere Length or iNOS
3.6. Telomere Length Is Not Correlated with p16 or iNOS Staining
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marino, C.L.; Lascelles, B.D.; Vaden, S.L.; Gruen, M.E.; Marks, S.L. Prevalence and classification of chronic kidney disease in cats randomly selected from four age groups and in cats recruited for degenerative joint disease studies. J. Feline Med. Surg. 2014, 16, 465–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lulich, J.P.; O’Brien, T.D.; Osborne, C.A.; Polzin, D.J. Feline renal failure:questions, answers, questions. Compend. Contin. Educ. Vet. 1992, 14, 127–152. [Google Scholar]
- McLeland, S.M.; Cianciolo, R.E.; Duncan, C.G.; Quimby, J.M. A comparison of biochemical and histopathologic staging in cats with chronic kidney disease. Vet. Pathol. 2015, 52, 524–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, C.A.; Elliott, J.; Schmiedt, C.W.; Brown, S.A. Chronic Kidney Disease in Aged Cats: Clinical Features, Morphology, and Proposed Pathogeneses. Vet. Pathol. 2016, 53, 309–326. [Google Scholar] [CrossRef] [PubMed]
- Jepson, R.E. Current Understanding of the Pathogenesis of Progressive Chronic Kidney Disease in Cats. Vet. Clin. N. Am. Small Anim. Pract. 2016, 46, 1015–1048. [Google Scholar] [CrossRef] [Green Version]
- Spencer, S.; Wheeler-Jones, C.; Elliott, J. Hypoxia and chronic kidney disease: Possible mechanisms, therapeutic targets, and relevance to cats. Vet. J. 2021, 274, 105714. [Google Scholar] [CrossRef]
- Lawson, J.S.; Liu, H.H.; Syme, H.M.; Purcell, R.; Wh.heeler-Jones, C.P.D.; Elliott, J. The cat as a naturally occurring model of renal interstitial fibrosis: Characterisation of primary feline proximal tubular epithelial cells and comparative pro-fibrotic effects of TGF-beta1. PLoS ONE 2018, 13, e0202577. [Google Scholar] [CrossRef]
- Lawson, J.; Elliott, J.; Wheeler-Jones, C.; Syme, H.; Jepson, R. Renal fibrosis in feline chronic kidney disease: Known mediators and mechanisms of injury. Vet. J. 2015, 203, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.J.; Rakheja, D.; Yu, X.; Saxena, R.; Vaziri, N.D.; Silva, F.G. The aging kidney. Kidney Int. 2008, 74, 710–720. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, R. Senotherapy: Growing old and staying young? Pflugers Arch. 2017, 469, 1051–1059. [Google Scholar] [CrossRef]
- Knoppert, S.N.; Valentijn, F.A.; Nguyen, T.Q.; Goldschmeding, R.; Falke, L.L. Cellular Senescence and the Kidney: Potential Therapeutic Targets and Tools. Front. Pharmacol. 2019, 10, 770. [Google Scholar] [CrossRef] [PubMed]
- Valentijn, F.A.; Falke, L.L.; Nguyen, T.Q.; Goldschmeding, R. Cellular senescence in the aging and diseased kidney. J. Cell Commun. Signal. 2018, 12, 69–82. [Google Scholar] [CrossRef] [Green Version]
- Baar, M.P.; Brandt, R.M.C.; Putavet, D.A.; Klein, J.D.D.; Derks, K.W.J.; Bourgeois, B.R.M.; Stryeck, S.; Rijksen, Y.; van Willigenburg, H.; Feijtel, D.A.; et al. Targeted Apoptosis of Senescent Cells Restores Tissue Homeostasis in Response to Chemotoxicity and Aging. Cell 2017, 169, 132–147.e116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quimby, J.M.; Maranon, D.G.; Battaglia, C.L.; McLeland, S.M.; Brock, W.T.; Bailey, S.M. Feline chronic kidney disease is associated with shortened telomeres and increased cellular senescence. Am. J. Physiol. Renal Physiol. 2013, 305, F295–F303. [Google Scholar] [CrossRef] [Green Version]
- Melk, A.; Schmidt, B.M.; Takeuchi, O.; Sawitzki, B.; Rayner, D.C.; Halloran, P.F. Expression of p16INK4a and other cell cycle regulator and senescence associated genes in aging human kidney. Kidney Int. 2004, 65, 510–520. [Google Scholar] [CrossRef] [Green Version]
- Melk, A.; Schmidt, B.M.; Vongwiwatana, A.; Rayner, D.C.; Halloran, P.F. Increased expression of senescence-associated cell cycle inhibitor p16INK4a in deteriorating renal transplants and diseased native kidney. Am. J. Transplant. 2005, 5, 1375–1382. [Google Scholar] [CrossRef]
- Fernandez-Moreno, M.; Tamayo, M.; Soto-Hermida, A.; Mosquera, A.; Oreiro, N.; Fernandez-Lopez, C.; Fernandez, J.L.; Rego-Perez, I.; Blanco, F.J. mtDNA haplogroup J modulates telomere length and nitric oxide production. BMC Musculoskelet Disord. 2011, 12, 283. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Bae, E.H.; Ma, S.K.; Kim, S.W. Altered Nitric Oxide System in Cardiovascular and Renal Diseases. Chonnam. Med. J. 2016, 52, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Modlinger, P.S.; Wilcox, C.S.; Aslam, S. Nitric oxide, oxidative stress, and progression of chronic renal failure. Semin. Nephrol. 2004, 24, 354–365. [Google Scholar] [CrossRef] [PubMed]
- Nentwig, A.; Schweighauser, A.; Maissen-Villiger, C.; Bruckmaier, R.M.; Zurbriggen, A.; van Dorland, H.A.; Francey, T. Assessment of the expression of biomarkers of uremic inflammation in dogs with renal disease. Am. J. Vet. Res. 2016, 77, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Ratliff, B.B.; Abdulmahdi, W.; Pawar, R.; Wolin, M.S. Oxidant Mechanisms in Renal Injury and Disease. Antioxid. Redox Signal. 2016, 25, 119–146. [Google Scholar] [CrossRef] [Green Version]
- International Renal Interest Society. Available online: http://www.iris-kidney.com/pdf/IRIS_Staging_of_CKD_modified_2019.pdf. (accessed on 6 February 2020).
- Munday, J.S.; Knight, C.G.; French, A.F. Evaluation of feline oral squamous cell carcinomas for p16CDKN2A protein immunoreactivity and the presence of papillomaviral DNA. Res. Vet. Sci. 2011, 90, 280–283. [Google Scholar] [CrossRef]
- Islam, M.S.; Matsumoto, M.; Hidaka, R.; Miyoshi, N.; Yasuda, N. Expression of NOS and VEGF in feline mammary tumours and their correlation with angiogenesis. Vet. J. 2012, 192, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Brandt, L.E.; Bohn, A.A.; Charles, J.B.; Ehrhart, E.J. Localization of canine, feline, and mouse renal membrane proteins. Vet. Pathol. 2012, 49, 693–703. [Google Scholar] [CrossRef]
- Reyes, J.L.; Lamas, M.; Martin, D.; del Carmen Namorado, M.; Islas, S.; Luna, J.; Tauc, M.; Gonzalez-Mariscal, L. The renal segmental distribution of claudins changes with development. Kidney Int. 2002, 62, 476–487. [Google Scholar] [CrossRef]
- Hall, J.A.; Yerramilli, M.; Obare, E.; Jewell, D.E. Comparison of serum concentrations of symmetric dimethylarginine and creatinine as kidney function biomarkers in cats with chronic kidney disease. J. Vet. Intern. Med. 2014, 28, 1676–1683. [Google Scholar] [CrossRef] [Green Version]
- Verzola, D.; Gandolfo, M.T.; Gaetani, G.; Ferraris, A.; Mangerini, R.; Ferrario, F.; Villaggio, B.; Gianiorio, F.; Tosetti, F.; Weiss, U.; et al. Accelerated senescence in the kidneys of patients with type 2 diabetic nephropathy. Am. J. Physiol. Renal Physiol. 2008, 295, F1563–F1573. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Yang, J.R.; He, Y.N.; Cai, G.Y.; Zhang, J.G.; Lin, L.R.; Zhan, J.; Zhang, J.H.; Xiao, H.S. Accelerated senescence of renal tubular epithelial cells is associated with disease progression of patients with immunoglobulin A (IgA) nephropathy. Transl. Res. 2012, 159, 454–463. [Google Scholar] [CrossRef]
- Sis, B.; Tasanarong, A.; Khoshjou, F.; Dadras, F.; Solez, K.; Halloran, P.F. Accelerated expression of senescence associated cell cycle inhibitor p16INK4A in kidneys with glomerular disease. Kidney Int. 2007, 71, 218–226. [Google Scholar] [CrossRef] [Green Version]
- Bland, S.K.; Schmiedt, C.W.; Clark, M.E.; DeLay, J.; Bienzle, D. Expression of Kidney Injury Molecule-1 in Healthy and Diseased Feline Kidney Tissue. Vet. Pathol. 2017, 54, 490–510. [Google Scholar] [CrossRef] [Green Version]
- Cowgill, L.D.; Polzin, D.J.; Elliott, J.; Nabity, M.B.; Segev, G.; Grauer, G.F.; Brown, S.; Langston, C.; van Dongen, A.M. Is Progressive Chronic Kidney Disease a Slow Acute Kidney Injury? Vet. Clin. N. Am. Small Anim. Pract. 2016, 46, 995–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarry-Adkins, J.L.; Ozanne, S.E.; Norden, A.; Cherif, H.; Hales, C.N. Lower antioxidant capacity and elevated p53 and p21 may be a link between gender disparity in renal telomere shortening, albuminuria, and longevity. Am. J. Physiol. Renal Physiol. 2006, 290, F509–F516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joosten, S.A.; van Ham, V.; Nolan, C.E.; Borrias, M.C.; Jardine, A.G.; Shiels, P.G.; van Kooten, C.; Paul, L.C. Telomere shortening and cellular senescence in a model of chronic renal allograft rejection. Am. J. Pathol 2003, 162, 1305–1312. [Google Scholar] [CrossRef] [Green Version]
- Hamet, P.; Thorin-Trescases, N.; Moreau, P.; Dumas, P.; Tea, B.S.; deBlois, D.; Kren, V.; Pravenec, M.; Kunes, J.; Sun, Y.; et al. Workshop: Excess growth and apoptosis: Is hypertension a case of accelerated aging of cardiovascular cells? Hypertension 2001, 37, 760–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nangaku, M. Chronic hypoxia and tubulointerstitial injury: A final common pathway to end-stage renal failure. J. Am. Soc. Nephrol. 2006, 17, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Lourenco, B.N.; Coleman, A.E.; Tarigo, J.L.; Berghaus, R.D.; Brown, C.A.; Rissi, D.R.; Stanton, J.B.; Brown, S.A.; Schmiedt, C.W. Evaluation of profibrotic gene transcription in renal tissues from cats with naturally occurring chronic kidney disease. J. Vet. Intern. Med. 2020, 34, 1476–1487. [Google Scholar] [CrossRef]
- Ameh, O.I.; Okpechi, I.G.; Dandara, C.; Kengne, A.P. Association Between Telomere Length, Chronic Kidney Disease, and Renal Traits: A Systematic Review. OMICS 2017, 21, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Cokan Vujkovac, A.; Novakovic, S.; Vujkovac, B.; Stevanec, M.; Skerl, P.; Sabovic, M. Aging in Fabry Disease: Role of Telomere Length, Telomerase Activity, and Kidney Disease. Nephron 2020, 144, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Fazzini, F.; Lamina, C.; Raschenberger, J.; Schultheiss, U.T.; Kotsis, F.; Schonherr, S.; Weissensteiner, H.; Forer, L.; Steinbrenner, I.; Meiselbach, H.; et al. Results from the German Chronic Kidney Disease (GCKD) study support association of relative telomere length with mortality in a large cohort of patients with moderate chronic kidney disease. Kidney Int. 2020, 98, 488–497. [Google Scholar] [CrossRef]
- Mazidi, M.; Rezaie, P.; Covic, A.; Malyszko, J.; Rysz, J.; Kengne, A.P.; Banach, M. Telomere attrition, kidney function, and prevalent chronic kidney disease in the United States. Oncotarget 2017, 8, 80175–80181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Lee, S.; Kim, Y.; Cho, S.; Kim, K.; Kim, Y.C.; Han, S.S.; Lee, H.; Lee, J.P.; Joo, K.W.; et al. A Mendelian randomization study found causal linkage between telomere attrition and chronic kidney disease. Kidney Int. 2021. [Google Scholar] [CrossRef]
- Harris, R.C.; Neilson, E.G. Toward a unified theory of renal progression. Annu. Rev. Med. 2006, 57, 365–380. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Iturbe, B.; Garcia Garcia, G. The role of tubulointerstitial inflammation in the progression of chronic renal failure. Nephron Clin. Pract. 2010, 116, c81–c88. [Google Scholar] [CrossRef] [PubMed]
- Lourenco, B.N.; Coleman, A.E.; Schmiedt, C.W.; Brown, C.A.; Rissi, D.R.; Stanton, J.B.; Giguere, S.; Berghaus, R.D.; Brown, S.A.; Tarigo, J.L. Profibrotic gene transcription in renal tissues from cats with ischemia-induced chronic kidney disease. Am. J. Vet. Res. 2020, 81, 180–189. [Google Scholar] [CrossRef]
- Houben, J.M.; Moonen, H.J.; van Schooten, F.J.; Hageman, G.J. Telomere length assessment: Biomarker of chronic oxidative stress? Free Radic. Biol. Med. 2008, 44, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Victorelli, S.; Passos, J.F. Telomeres and Cell Senescence-Size Matters Not. EBioMedicine 2017, 21, 14–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Fogo, A.B. Cell senescence in the aging kidney. J. Am. Soc. Nephrol. JASN 2010, 21, 1436–1439. [Google Scholar] [CrossRef] [Green Version]
- Keegan, R.F.; Webb, C.B. Oxidative stress and neutrophil function in cats with chronic renal failure. J. Vet. Intern. Med. 2010, 24, 514–519. [Google Scholar] [CrossRef] [PubMed]
- Whitehouse, W.; Quimby, J.; Wan, S.; Monaghan, K.; Robbins, R.; Trepanier, L.A. Urinary F2-Isoprostanes in Cats with International Renal Interest Society Stage 1-4 Chronic Kidney Disease. J. Vet. Intern. Med. 2017, 31, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Paetau-Robinson, I. Dietary supplements of vitamins E and C and beta-carotene reduce oxidative stress in cats with renal insufficiency. Vet. Res. Commun. 2006, 30, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Valle, E.; Prola, L.; Vergnano, D.; Borghi, R.; Monacelli, F.; Traverso, N.; Bruni, N.; Bovero, A.; Schiavone, A.; Nery, J.; et al. Investigation of hallmarks of carbonyl stress and formation of end products in feline chronic kidney disease as markers of uraemic toxins. J. Feline Med. Surg. 2019, 21, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Melk, A.; Ramassar, V.; Helms, L.M.; Moore, R.; Rayner, D.; Solez, K.; Halloran, P.F. Telomere shortening in kidneys with age. J. Am. Soc. Nephrol. 2000, 11, 444–453. [Google Scholar] [CrossRef]
- Tchakmakjian, L.; Gardner, J.P.; Wilson, P.D.; Kimura, M.; Skurnick, J.; Zielke, H.R.; Aviv, A. Age-dependent telomere attrition as a potential indicator of racial differences in renal growth patterns. Nephron Exp. Nephrol. 2004, 98, e82–e88. [Google Scholar] [CrossRef] [PubMed]
- Birch, J.; Anderson, R.K.; Correia-Melo, C.; Jurk, D.; Hewitt, G.; Marques, F.M.; Green, N.J.; Moisey, E.; Birrell, M.A.; Belvisi, M.G.; et al. DNA damage response at telomeres contributes to lung aging and chronic obstructive pulmonary disease. Am. J. Physiol. Lung Cell Mol. Physiol. 2015, 309, L1124–L1137. [Google Scholar] [CrossRef] [Green Version]
- Syme, H.M.; Barber, P.J.; Markwell, P.J.; Elliott, J. Prevalence of systolic hypertension in cats with chronic renal failure at initial evaluation. J. Am. Vet. Med. Assoc. 2002, 220, 1799–1804. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, D.L.; Peterson, M.E.; Graves, T.K.; Lesser, M.; Nichols, C.E. Hypertension in cats with chronic renal failure or hyperthyroidism. J. Vet. Intern. Med. 1990, 4, 58–62. [Google Scholar] [CrossRef]
- Stiles, J.; Polzin, D.; Bistner, S. The prevalence of retinopathy in cats with systemic hypertension and chronic renal failure or hyperthyroidism. J. Am. Anim. Hosp. Assoc. 1994, 30, 564–572. [Google Scholar]
- Chiu, C.L.; Hearn, N.L.; Paine, D.; Steiner, N.; Lind, J.M. Does Telomere Shortening Precede the Onset of Hypertension in Spontaneously Hypertensive Mice? Twin Res. Hum. Genet. 2016, 19, 422–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Li, R.; Yang, Y.; Chen, Y.; Yang, S.; Li, J.; Wu, C.; Kong, T.; Liu, T.; Cai, J.; et al. Longitudinal Association of Telomere Attrition with the Effects of Antihypertensive Treatment and Blood Pressure Lowering. Aging Dis. 2020, 11, 494–508. [Google Scholar] [CrossRef] [PubMed]
- Bijsmans, E.S.; Jepson, R.E.; Chang, Y.M.; Syme, H.M.; Elliott, J. Changes in Systolic Blood Pressure over Time in Healthy Cats and Cats with Chronic Kidney Disease. J. Vet. Intern. Med. 2015. [Google Scholar] [CrossRef] [PubMed]
- Jepson, R.E.; Brodbelt, D.; Vallance, C.; Syme, H.M.; Elliott, J. Evaluation of predictors of the development of azotemia in cats. J. Vet. Intern. Med. 2009, 23, 806–813. [Google Scholar] [CrossRef] [PubMed]
- Paepe, D.; Verjans, G.; Duchateau, L.; Piron, K.; Ghys, L.; Daminet, S. Routine health screening: Findings in apparently healthy middle-aged and old cats. J. Feline Med. Surg. 2013, 15, 8–19. [Google Scholar] [CrossRef]
- Cianciolo, R.E.; Mohr, F.C.; Aresu, L.; Brown, C.A.; James, C.; Jansen, J.H.; Spangler, W.L.; van der Lugt, J.J.; Kass, P.H.; Brovida, C.; et al. World Small Animal Veterinary Association Renal Pathology Initiative: Classification of Glomerular Diseases in Dogs. Vet. Pathol. 2016, 53, 113–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habenicht, L.M.; Webb, T.L.; Clauss, L.A.; Dow, S.W.; Quimby, J.M. Urinary cytokine levels in apparently healthy cats and cats with chronic kidney disease. J. Feline Med. Surg. 2013, 15, 99–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, J.S.; Syme, H.M.; Wheeler-Jones, C.P.; Elliott, J. Urinary active transforming growth factor beta in feline chronic kidney disease. Vet. J. 2016, 214, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Adult Cats | Senior Cats | CKD Cats | |
|---|---|---|---|
| p16 | (n = 14) | (n = 20) | (n = 30) |
| Age (years) a,b | 5 (2–9) | 12 (10–16) | 15 (3–20) |
| Sex | MN (8) FS (6) | MN (13) FS (7) | MN (20) FS (10) |
| Creatinine (mg/dL) b,c | 0.8 (0.3–1.5) | 1.1 (0.4–1.5) | 3.9 (1.7–10) |
| Telomere | (n = 19) | (n = 23) | (n = 54) |
| Age (years) a,b | 4 (0.6–9) | 12 (10–16) | 15 (3–21) |
| Sex | MN (13) FS (6) | MN (13) FS (10) | MN (35) FS (19) |
| Creatinine (mg/dL) b,c | 1.0 (0.4–1.5) | 1.1 (0.4–1.5) | 4.3 (1.7–13.6) |
| iNOS | (n = 15) | (n = 19) | (n = 43) |
| Age (years) a,b | 4 (0.6–9) | 12.5 (10–16) | 15 (4–21.4) |
| Sex | MN (10) FS (5) | MN (11) FS (8) | MN (28) FS (15) |
| Creatinine (mg/dL) b,c | 1.0 (0.3–1.5) | 1.1 (0.5–1.5) | 3.9 (1.7–10) |
| Marker | Type, Clone, Reference | Supplier | Dilution |
|---|---|---|---|
| AQP1 | Rabbit polyclonal anti-aquaporin-1 AP, AB2219 [14] | Millipore; Billerica, MA, USA | 1:1000 |
| p16 | Mouse monoclonal anti-human p16, Cat#550834 [23] | BD Biosciences; San Jose, CA, USA | 1:25 |
| iNOS | Rabbit polyclonal anti-inducible nitric oxide synthase, NB300-605 [24] | Fisher Scientific, Waltham, MA, USA | 1:100 |
| Adult Cats (n = 14) | Senior Cats (n = 20) | CKD Cats (n = 30) | |
|---|---|---|---|
| % Global GS b,c | 0 (0–14) | 2 (0–40) | 15 (0–92) |
| Inflammatory Infiltrate Score Cortex b,c | 0 (0–1) | 0 (0–0.7) | 0.9 (0–2.9) |
| Fibrosis Score Cortex b,c | 0 (0–1) | 0 (0–0.6) | 0.8 (0–2.7) |
| Inflammatory Infiltrate Score CMJ b,c | 0 (0–1) | 0 (0–0.5) | 1.5 (0.2–2.5) |
| Fibrosis Score CMJ b,c | 0.1 (0–1.4) | 0 (0–1) | 1.5 (0.3–2.5) |
| % p16+AQP1+ Cortex b,c | 0 (0–22.3) | 2.6 (0–20.6) | 10.6 (0–55) |
| % p16+AQP1− Cortex b,c | 3.7 (0–35) | 15.5 (0–42.6) | 28.5 (5.3–65.9) |
| SQ p16+AQP1+ Cortex b,c | 0 (0–2) | 0 (0–1.5) | 1 (0–3) |
| SQ p16+AQP1− Cortex b,c | 0 (0–1) | 0.25 (0–1.5) | 1 (0–3) |
| % p16+AQP1+ CMJ a,b | 1.2 (0–26.6) | 12.3 (0–30.9) | 12.2 (1.2–48.2) |
| % p16+AQP1− CMJ b,c | 1.1 (0–26.5) | 2.7 (0–25.8) | 23.8 (1.7–52) |
| SQ p16+AQP1+ CMJ a | 0 (0–2) | 1 (0–2) | 0.5 (0–2.5) |
| SQ p16+AQP1− CMJ b,c | 0 (0–0.5) | 0 (0–0.5) | 1 (0–3) |
| Adult Cats (n = 19) | Senior Cats (n = 23) | CKD Cats (n = 54) | |
|---|---|---|---|
| % Global GS b,c | 0 (0–16) | 2 (0–40) | 21 (0–92) |
| Inflammatory Infiltrate Score Cortex b,c | 0 (0–1.0) | 0.1 (0–0.7) | 1.1 (0–2.9) |
| Fibrosis Score Cortex b,c | 0 (0–1.0) | 0 (0–0.8) | 0.8 (0–2.7) |
| Mean Telomere TFI b,c | 556 ± 41.9 | 524.2 ± 39.7 | 429.4 ± 31.4 |
| % Short Telomeres d | 7% ± 1% | 8% ± 1% | 10% ± 1% |
| Mean Telomere TFI | p-Value | % Short Telomeres | p-Value | |
|---|---|---|---|---|
| Normal Cats | 0.749 | 0.335 | ||
| Age (years) | ||||
| <5 (n = 10) | 538.4 ± 66.4 | 6% ± 1% | ||
| 5–9 (n = 9) | 600.3 ± 68.5 | 6% ± 2% | ||
| 10–15 (n = 20) | 542.0 ± 50.3 | 7% ± 1% | ||
| >15 (n = 3) | 489.99 ± 117.9 | 11% ± 4% | ||
| CKD IRIS Stage | 0.9313 | 0.9101 | ||
| IRIS Stage 2 (n = 14) | 440.9 ± 47.3 | 10% ± 2% | ||
| IRIS Stage 3 (n = 17) | 452.5 ± 46.1 | 10% ± 2% | ||
| IRIS Stage 4 (n = 23) | 429.0 ± 41.7 | 11% ± 2% | ||
| CKD Proteinuria | 0.572 | 0.770 | ||
| NP (n = 15) | 431.1 ± 50 | 11% ± 2% | ||
| BP (n = 17) | 418 ± 43.3 | 10% ± 2% | ||
| P (n = 20) | 470.2 ± 42.3 | 10% ± 2% | ||
| CKD Hypertension | 0.03 | 0.057 | ||
| Yes (n = 23) | 384.8 ± 38 | 13% ± 2% | ||
| No (n = 31) | 481.2 ± 34.7 | 9% ± 1% |
| Adult Cats (n = 15) | Senior Cats (n = 19) | CKD Cats (n = 43) | |
|---|---|---|---|
| % Global GS b,c | 0 (0–14) | 2 (0–40) | 21 (0–92) |
| Inflammatory Infiltrate Score Cortex b,c | 0 (0–1) | 0 (0–0.7) | 1.1 (0–2.9) |
| Fibrosis Score Cortex b,c | 0 (0–1) | 0 (0–0.8) | 0.7 (0–1.9) |
| Inflammatory Infiltrate Score CMJ b,c | 0 (0–1) | 0 (0–0) | 1.3 (0–2.5) |
| Fibrosis Score CMJ b,c | 0.1 (0–1.4) | 0 (0–1) | 1.5 (0.2–2.1) |
| % iNOS+ AQP1+ Cortex a,b | 4 (0–99) | 82 (0–100) | 90 (0–100) |
| % iNOS+ AQP1− Cortex | 64 (35–98) | 84 (28–100) | 92 (21–100) |
| INT iNOS+ AQP1+ Cortex b | 1 (0–1.2) | 1 (0–1.8) | 1 (0–2) |
| INT iNOS+ AQP1− Cortex | 1.5 (1–2.9) | 1.9 (1–2.9) | 1.6 (1–2.8) |
| % iNOS+ AQP1+ CMJ d,e | 37 (0–100) | 94.5 (0–100) | 89 (0–100) |
| % iNOS+ AQP1− CMJ | 50 (4–100) | 91.5 (1–100) | 90.5 (3–100) |
| INT iNOS+ AQP1+ CMJ | 1 (0–1) | 1 (0–2) | 1 (0–2) |
| INT iNOS+ AQP1− CMJ | 1.3 (1–2) | 1.2 (1–2.1) | 1 (0–2) |
| % GGS | Inflammatory Infiltrate Score | Fibrosis Score | |
|---|---|---|---|
| Cortex | |||
| % p16+AQP1+ | p = 0.003 | p = 0.006 | p = 0.0008 |
| r = 0.37 | r = 0.35 | r = 0.41 | |
| % p16+AQP1− | p = 0.0002 | p < 0.0001 | p < 0.0001 |
| r = 0.46 | r = 0.53 | r = 0.53 | |
| CMJ | |||
| % p16+AQP1+ | NA | NS | p = 0.04 |
| r = 0.26 | |||
| % p16+AQP1− | NA | p < 0.0001 | p < 0.0001 |
| r = 0.66 | r = 0.72 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quimby, J.; Erickson, A.; Mcleland, S.; Cianciolo, R.; Maranon, D.; Lunn, K.; Elliott, J.; Lawson, J.; Hess, A.; Paschall, R.; et al. Renal Senescence, Telomere Shortening and Nitrosative Stress in Feline Chronic Kidney Disease. Vet. Sci. 2021, 8, 314. https://doi.org/10.3390/vetsci8120314
Quimby J, Erickson A, Mcleland S, Cianciolo R, Maranon D, Lunn K, Elliott J, Lawson J, Hess A, Paschall R, et al. Renal Senescence, Telomere Shortening and Nitrosative Stress in Feline Chronic Kidney Disease. Veterinary Sciences. 2021; 8(12):314. https://doi.org/10.3390/vetsci8120314
Chicago/Turabian StyleQuimby, Jessica, Andrea Erickson, Shannon Mcleland, Rachel Cianciolo, David Maranon, Katharine Lunn, Jonathan Elliott, Jack Lawson, Ann Hess, Rene Paschall, and et al. 2021. "Renal Senescence, Telomere Shortening and Nitrosative Stress in Feline Chronic Kidney Disease" Veterinary Sciences 8, no. 12: 314. https://doi.org/10.3390/vetsci8120314
APA StyleQuimby, J., Erickson, A., Mcleland, S., Cianciolo, R., Maranon, D., Lunn, K., Elliott, J., Lawson, J., Hess, A., Paschall, R., & Bailey, S. (2021). Renal Senescence, Telomere Shortening and Nitrosative Stress in Feline Chronic Kidney Disease. Veterinary Sciences, 8(12), 314. https://doi.org/10.3390/vetsci8120314