
Molecular Prevalence of Equine Parvovirus-Hepatitis in the Sera of Clinically Healthy Horses in South Korea



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Size Calculation and Collection
2.2. Sample Processing and PCR
2.3. Phylogenetic Analysis
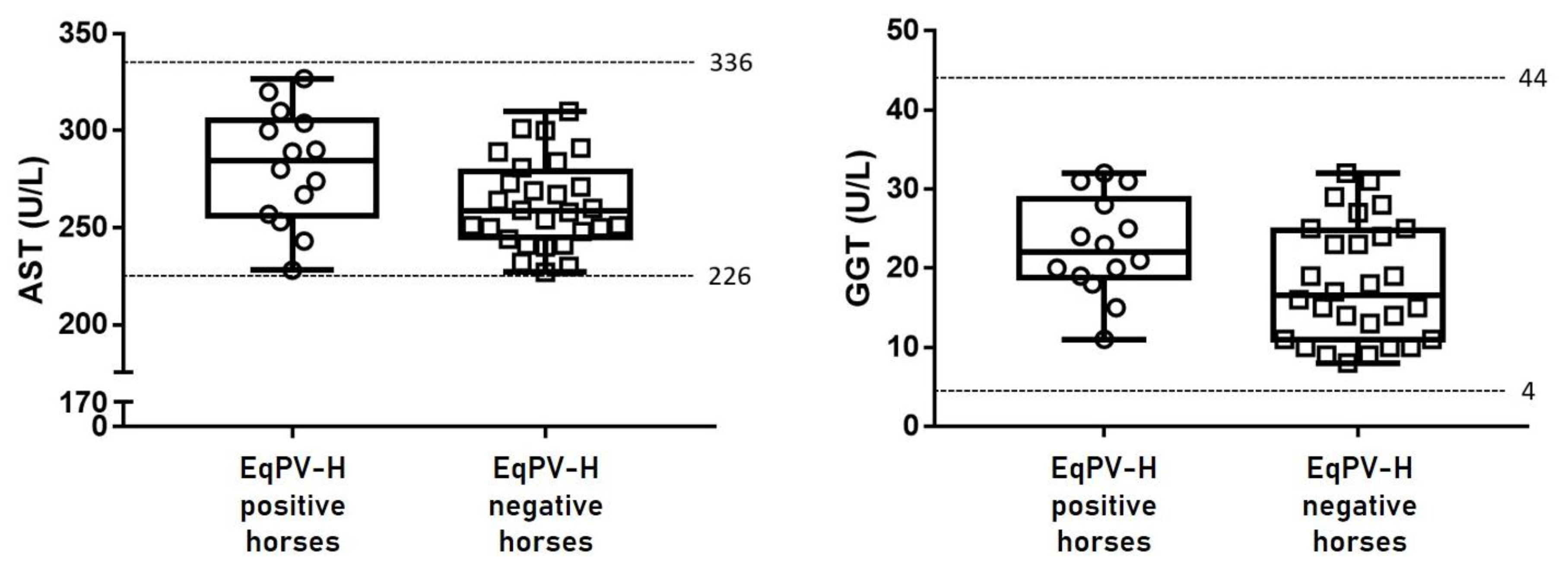
2.4. Serum Biochemistry
2.5. Statistical Analysis
3. Results
3.1. Prevalence of EqPV-H DNA in the Sera of Clinically Normal Horses
3.2. Phylogenetic Analysis
3.3. Liver-Specific Biochemistry
3.4. EqPV-H Infection According to Sex, Age, and Performance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Divers, T.J.; Tennant, B.C.; Kumar, A.; McDonough, S.; Cullen, J.; Bhuva, N.; Jain, K.; Chauhan, L.S.; Scheel, T.K.H.; Lipkin, W.I.; et al. New Parvovirus Associated with Serum Hepatitis in Horses after Inoculation of Common Biological Product. Emerg. Infect. Dis. 2018, 24, 303–310. [Google Scholar] [CrossRef]
- Pénzes, J.J.; Söderlund-Venermo, M.; Canuti, M.; Eis-Hübinger, A.M.; Hughes, J.; Cotmore, S.F.; Harrach, B. Reorganizing the family Parvoviridae: A revised taxonomy independent of the canonical approach based on host association. Arch. Virol. 2020, 165, 2133–2146. [Google Scholar] [CrossRef] [PubMed]
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Adriaenssens, E.M.; Dempsey, D.M.; Dutilh, B.E.; Harrach, B.; Harrison, R.L.; Hendrickson, R.C.; et al. Changes to virus taxonomy and the Statutes ratified by the International Committee on Taxonomy of Viruses (2020). Arch. Virol. 2020, 165, 2737–2748. [Google Scholar] [CrossRef]
- Meister, T.L.; Tegtmeyer, B.; Postel, A.; Cavalleri, J.V.; Todt, D.; Stang, A.; Steinmann, E. Equine Parvovirus-Hepatitis Frequently Detectable in Commercial Equine Serum Pools. Viruses 2019, 11, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotmore, S.F.; Agbandje-McKenna, M.; Chiorini, J.A.; Mukha, D.V.; Pintel, D.J.; Qiu, J.; Soderlund-Venermo, M.; Tattersall, P.; Tijssen, P.; Gatherer, D.; et al. The family Parvoviridae. Arch. Virol. 2014, 159, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Theiler, A. Acute liver-atrophy and parenchymatous hepatitis in horses. In 5th and 6th Repts. of the Director of Veterinary Research; Department of Agriculture, Union of South Africa: Pretoria, South Africa, 1918; pp. 7–165. [Google Scholar]
- Reinecke, B.; Klöhn, M.; Brüggemann, Y.; Kinast, V.; Todt, D.; Stang, A.; Badenhorst, M.; Koeppel, K.; Guthrie, A.; Groner, U.; et al. Clinical Course of Infection and Cross-Species Detection of Equine Parvovirus-Hepatitis. Viruses 2021, 13, 1454. [Google Scholar] [CrossRef] [PubMed]
- Sturgeon, B. Theiler’s disease. Vet. Rec. 2017, 180, 14–15. [Google Scholar] [CrossRef] [PubMed]
- Meister, T.L.; Tegtmeyer, B.; Brüggemann, Y.; Sieme, H.; Feige, K.; Todt, D.; Stang, A.; Cavalleri, J.V.; Steinmann, E. Characterization of Equine Parvovirus in Thoroughbred Breeding Horses from Germany. Viruses 2019, 11, 965. [Google Scholar] [CrossRef] [Green Version]
- Divers, T.J.; Tomlinson, J.E. Theiler’s disease. Equine Vet. Educ. 2020, 32, 63–65. [Google Scholar] [CrossRef]
- Baird, J.; Tegtmeyer, B.; Arroyo, L.; Stang, A.; Brüggemann, Y.; Hazlett, M.; Steinmann, E. The association of Equine Parvovirus-Hepatitis (EqPV-H) with cases of non-biologic-associated Theiler’s disease on a farm in Ontario, Canada. Vet. Microbiol. 2020, 242, 108575. [Google Scholar] [CrossRef]
- Tomlinson, J.E.; Tennant, B.C.; Struzyna, A.; Mrad, D.; Browne, N.; Whelchel, D.; Johnson, P.J.; Jamieson, C.; Löhr, C.V.; Bildfell, R.; et al. Viral testing of 10 cases of Theiler’s disease and 37 in-contact horses in the absence of equine biologic product administration: A prospective study (2014–2018). J. Vet. Intern. Med. 2019, 33, 258–265. [Google Scholar] [CrossRef]
- Tomlinson, J.E.; Kapoor, A.; Kumar, A.; Tennant, B.C.; Laverack, M.A.; Beard, L.; Delph, K.; Davis, E.; Schott Ii, H.; Lascola, K.; et al. Viral testing of 18 consecutive cases of equine serum hepatitis: A prospective study (2014–2018). J. Vet. Intern. Med. 2019, 33, 251–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsh, H. Losses of undetermined cause following an outbreak of equine encephalomyelitis. J. Am. Vet. Med. Assoc. 1937, 91, 88–93. [Google Scholar]
- Thomsett, L.R. Acute hepatic failure in the horse. Equine Vet. J. 1971, 3, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Chandriani, S.; Skewes-Cox, P.; Zhong, W.; Ganem, D.E.; Divers, T.J.; Van Blaricum, A.J.; Tennant, B.C.; Kistler, A.L. Identification of a previously undescribed divergent virus from the Flaviviridae family in an outbreak of equine serum hepatitis. Proc. Natl. Acad. Sci. USA 2013, 110, E1407–E1415. [Google Scholar] [CrossRef] [Green Version]
- Vengust, M.; Jager, M.C.; Zalig, V.; Cociancich, V.; Laverack, M.; Renshaw, R.W.; Dubovi, E.; Tomlinson, J.E.; Van de Walle, G.R.; Divers, T.J. First report of equine parvovirus-hepatitis-associated Theiler’s disease in Europe. Equine Vet. J. 2020, 52, 841–847. [Google Scholar] [CrossRef] [PubMed]
- Badenhorst, M.; de Heus, P.; Auer, A.; Tegtmeyer, B.; Stang, A.; Dimmel, K.; Tichy, A.; Kubacki, J.; Bachofen, C.; Steinmann, E.; et al. Active equine parvovirus-hepatitis infection is most frequently detected in Austrian horses of advanced age. Equine Vet. J. 2021. Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Sun, L.; Ou, J.; Xu, H.; Wu, L.; Li, S. Identification and genetic characterization of a novel parvovirus associated with serum hepatitis in horses in China. Emerg. Microbes Infect. 2018, 7, 170. [Google Scholar] [CrossRef] [Green Version]
- Lu, G.; Wu, L.; Ou, J.; Li, S. Equine Parvovirus-Hepatitis in China: Characterization of Its Genetic Diversity and Evidence for Natural Recombination Events Between the Chinese and American Strains. Front. Vet. Sci. 2020, 7, 121. [Google Scholar] [CrossRef]
- de Moraes, M.V.D.S.; Salgado, C.R.S.; Godoi, T.L.O.S.; de Almeida, F.Q.; Chalhoub, F.L.L.; de Filippis, A.M.B.; de Souza, A.M.; de Oliveira, J.M.; Figueiredo, A.S. Equine parvovirus-hepatitis is detected in South America, Brazil. Transbound Emerg. Dis. 2021. Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Ramsauer, A.S.; Badenhorst, M.; Cavalleri, J.V. Equine parvovirus hepatitis. Equine Vet. J. 2021, 53, 886–894. [Google Scholar] [CrossRef] [PubMed]
- Horse Registry. The current Status of Registered Horses in South Korea. Available online: https://www.horsepia.com/pa/hh/PAHH3000/pHorseRegStateMain.do (accessed on 27 September 2021).
- Sergeant, E.S.G. Epitools Epidemiological Calculators. Available online: http://epitools.ausvet.com.au (accessed on 27 September 2021).
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Waddell, P.J.; Steel, M.A. General Time-Reversible Distances with Unequal Rates across Sites: Mixing Γ and Inverse Gaussian Distributions with Invariant Sites. Mol. Phylogenet. Evol. 1997, 8, 398–414. [Google Scholar] [CrossRef] [Green Version]
- Orsini, J.A.; Divers, T.J. Equine Emergencies, 3rd ed.; W.B. Saunders: Saint Louis, MA, USA, 2008; pp. 755–766. [Google Scholar] [CrossRef]
- Tomlinson, J.E.; Van de Walle, G.R.; Divers, T.J. What Do We Know About Hepatitis Viruses in Horses? Vet. Clin. N. Am. Equine Pract. 2019, 35, 351–362. [Google Scholar] [CrossRef]
- Kopper, J.J.; Schott, H.C., II; Divers, T.J.; Mullaney, T.; Huang, L.; Noland, E.; Smedley, R. Theiler’s disease associated with administration of tetanus antitoxin contaminated with nonprimate (equine) hepacivirus and equine parvovirus-hepatitis virus. Equine Vet. Educ. 2020, 32, e5–e9. [Google Scholar] [CrossRef]
- Animal Disease Control Division. The Current List of Registered Animal Medicine (3rd Quarter of 2021). Available online: http://www.qia.go.kr/viewwebQiaCom.do?id=53739&type=2_23lylyy (accessed on 7 October 2021).
- Tomlinson, J.E.; Jager, M.; Struzyna, A.; Laverack, M.; Fortier, L.A.; Dubovi, E.; Foil, L.D.; Burbelo, P.D.; Divers, T.J.; Van de Walle, G.R. Tropism, pathology, and transmission of equine parvovirus-hepatitis. Emerg. Microbes Infect. 2020, 9, 651–663. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Property | Number (Sample ID) | Breed | Sex | Purpose | EqPV-H-Positive | Age (Years) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TB | P | O | S | G | M | RC | RD | B | O | 2–4 | 5–7 | ≥8 | |||
| KRA Seoul | 153 (KRA1-153) | 9/153 | 0/0 | 0/0 | 7/66 | 2/44 | 0/43 | 9/153 | 0/0 | 0/0 | 0/0 | 9/153 (5.9%) | 4/74 | 5/78 | 0/1 |
| KRA Busan | 80 (KRA242-321) | 3/76 | 1/2 | 1/2 | 0/26 | 5/20 | 0/34 | 2/75 | 3/5 | 0 | 0 | 5/80 (6.3%) | 2/59 | 1/16 | 2/5 |
| KRA Jangsu | 30 (KRA154-183) | 0/16 | 0/2 | 0/12 | 0/13 | 0/6 | 0/11 | 0/0 | 0/0 | 0/16 | 0/14 | 0/30 (0%) | 0/3 | 0/9 | 0/18 |
| KRA Jeju | 58 (KRA184-241) | 0/14 | 0/28 | 0/16 | 0/4 | 0/40 | 0/14 | 0/29 | 0/20 | 0/0 | 0/9 | 0/58 (0%) | 0/6 | 0/17 | 0/35 |
| Total | 321 | 12/259 | 1/32 | 1/30 | 7/109 | 7/110 | 0/102 | 11/257 | 3/25 | 0/16 | 0/23 | 14/321 (4.4%) | 6/142 | 6/120 | 2/59 |
| Comparison | p-Value | OR | 95% CI of OR | |
|---|---|---|---|---|
| Lower Limit | Upper Limit | |||
| Sex | ||||
| Male vs. Female | 0.006 | 0.936 | 0.904 | 0.969 |
| Age | ||||
| 2–4-year-old vs. 5–7-year-old | 0.765 | 0.838 | 0.263 | 2.67 |
| 2–4-year-old vs. >8-year-old | >0.99 | 1.257 | 0.246 | 6.417 |
| 5–7-year-old vs. >8-year-old | >0.99 | 1.5 | 0.293 | 7.668 |
| EqPV-H Status | Number | Performance Levels | ||
|---|---|---|---|---|
| High (≤33.3%) | Moderate (>33.3% to ≤66.7%) | Low (>66.7%) | ||
| Positive | 11 | 2 (18.2%) | 4 (36.4%) | 5 (45.5%) |
| Negative | 11 | 7 (63.6%) | 2 (18.2%) | 2 (18.2%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-K.; Park, D.; Lee, I. Molecular Prevalence of Equine Parvovirus-Hepatitis in the Sera of Clinically Healthy Horses in South Korea. Vet. Sci. 2021, 8, 282. https://doi.org/10.3390/vetsci8110282
Lee S-K, Park D, Lee I. Molecular Prevalence of Equine Parvovirus-Hepatitis in the Sera of Clinically Healthy Horses in South Korea. Veterinary Sciences. 2021; 8(11):282. https://doi.org/10.3390/vetsci8110282
Chicago/Turabian StyleLee, Sang-Kyu, Dongsun Park, and Inhyung Lee. 2021. "Molecular Prevalence of Equine Parvovirus-Hepatitis in the Sera of Clinically Healthy Horses in South Korea" Veterinary Sciences 8, no. 11: 282. https://doi.org/10.3390/vetsci8110282
APA StyleLee, S.-K., Park, D., & Lee, I. (2021). Molecular Prevalence of Equine Parvovirus-Hepatitis in the Sera of Clinically Healthy Horses in South Korea. Veterinary Sciences, 8(11), 282. https://doi.org/10.3390/vetsci8110282