Infectious Bronchitis Virus in Egypt: Genetic Diversity and Vaccination Strategies



{kind=link}
Abstract
1. Introduction
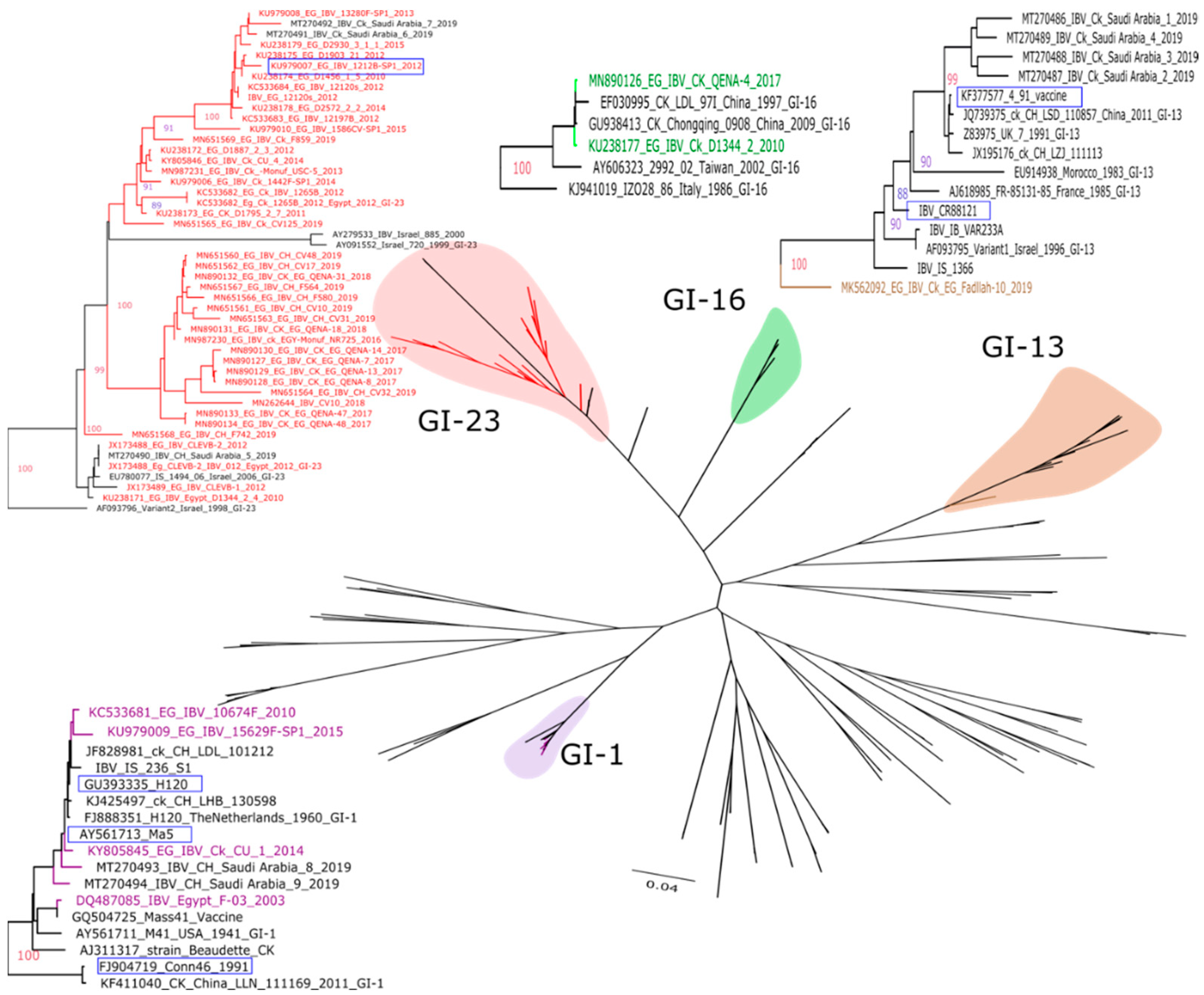
2. Multiple IBV Genotypes Circulating in Egypt
3. Genetic Drift and Recombination of the Egyptian IBV Strains
4. IBV Vaccines in Egypt
5. Perspectives and Recommendations for a Better Control Strategy
Author Contributions
Funding
Conflicts of Interest
References
- De Wit, J.J.S.; Cook, J.K.A. Spotlight on avian pathology: Infectious bronchitis virus. Avian Pathol. 2019, 48, 393–395. [Google Scholar] [CrossRef] [PubMed]
- Miłek, J.; Blicharz-Domańska, K. Coronaviruses in avian species—Review with focus on epidemiology and diagnosis in wild birds. J. Vet. Res. 2018, 62, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Rohaim, M.A.; El Naggar, R.F.; Helal, A.M.; Bayoumi, M.M.; El-Saied, M.A.; Ahmed, K.A.; Shabbir, M.Z.; Munir, M. Genetic diversity and phylodynamics of avian coronaviruses in Egyptian wild birds. Viruses 2019, 11, 57. [Google Scholar] [CrossRef]
- Payne, S. Chapter 17—Family coronaviridae. In Viruses; Payne, S., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 149–158. [Google Scholar] [CrossRef]
- Ennaji, Y.; Khataby, K.; Ennaji, M.M. Infectious bronchitis virus in poultry: Molecular epidemiology and factors leading to the emergence and reemergence of novel strains of infectious bronchitis virus. Emerg. Reemerg. Viral Pathog. 2020, 31–44. [Google Scholar] [CrossRef]
- De Wit, S.J.J.; Cook, J.K.A.; van der Heijden, H.M.J.F. Infectious bronchitis virus variants: A review of the history, current situation and control measures. Avian Pathol. 2011, 40, 223–235. [Google Scholar] [CrossRef]
- Valastro, V.; Holmes, E.C.; Britton, P.; Fusaro, A.; Jackwood, M.W.; Cattoli, G.; Monne, I. S1 gene-based phylogeny of infectious bronchitis virus: An attempt to harmonize virus classification. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2016, 39, 349–364. [Google Scholar] [CrossRef]
- Shan, D.; Fang, S.; Han, Z.; Ai, H.; Zhao, W.; Chen, Y.; Jiang, L.; Liu, S. Effects of hypervariable regions in spike protein on pathogenicity, tropism, and serotypes of infectious bronchitis virus. Virus Res. 2018, 250, 104–113. [Google Scholar] [CrossRef]
- Huang, Y.-P.; Lee, H.-C.; Cheng, M.-C.; Wang, C.-H. S1 and N gene analysis of avian infectious bronchitis viruses in Taiwan. Avian Dis. 2004, 48, 581–589. [Google Scholar] [CrossRef]
- Mase, M.; Tsukamoto, K.; Imai, K.; Yamaguchi, S. Phylogenetic analysis of avian infectious bronchitis virus strains isolated in Japan. Arch. Virol. 2004, 149, 2069–2078. [Google Scholar] [CrossRef]
- Abro, S.H.; Renström, L.H.; Ullman, K.; Isaksson, M.; Zohari, S.; Jansson, D.S.; Belák, S.; Baule, C. Emergence of novel strains of avian infectious bronchitis virus in Sweden. Vet. Microbiol. 2012, 155, 237–246. [Google Scholar] [CrossRef]
- De Wit, J.J.; Nieuwenhuisen-van Wilgen, J.; Hoogkamer, A.; van de Sande, H.; Zuidam, G.J.; Fabri, T.H. Induction of cystic oviducts and protection against early challenge with infectious bronchitis virus serotype D388 (genotype QX) by maternally derived antibodies and by early vaccination. Avian Pathol. 2011, 40, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Toffan, A.; Bonci, M.; Bano, L.; Bano, L.; Valastro, V.; Vascellari, M.; Capua, I.; Terregino, C. Diagnostic and clinical observation on the infectious bronchitis virus strain Q1 in Italy. Vet. Ital. 2013, 49, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Toffan, A.; Monne, I.; Terregino, C.; Cattoli, G.; Hodobo, C.T.; Gadaga, B.; Makaya, P.V.; Mdlongwa, E.; Swiswa, S. QX-like infectious bronchitis virus in Africa. Vet. Rec. 2011, 169, 589. [Google Scholar] [CrossRef] [PubMed]
- Ababneh, M.; Dalab, A.E.; Alsaad, S.; Al-Zghoul, M. Presence of infectious bronchitis virus strain CK/CH/LDL/97I in the Middle East. ISRN Vet. Sci. 2012, 2012, 201721. [Google Scholar] [CrossRef] [PubMed]
- Amin, O.G.M.; Valastro, V.; Salviato, A.; Drago, A.; Cattoli, G.; Monne, I. Circulation of QX-like infectious bronchitis virus in the Middle East. Vet. Rec. 2012, 171, 530. [Google Scholar] [CrossRef]
- Ahmed, H.N. Incidence and Treatment of Some Infectious Viral Respiratory Diseases of Poultry in Egypt. Ph.D. Thesis, Cairo University, Giza, Egypt, 1954. [Google Scholar]
- Davelaar, F.G.; Kouwenhoven, B.; Burger, A.G. Occurrence and significance of infectious bronchitis virus variant strains in egg and broiler production in the Netherlands. Vet. Q. 1984, 6, 114–120. [Google Scholar] [CrossRef]
- Taha, M.; Moustafa, M.M.; Mohamed, S.; Attia, S.A.; Hatem, M.E. Effect of both mycoplasma galliseplicum and infectious bronchitis virus classical and variant strains on the egg production of broiler breeder flocks. Vet. Med. 1991, 39, 169–179. [Google Scholar]
- Rohaim, M.A.; El Naggar, R.F.; Hamoud, M.M.; Bazid, A.I.; Gamal, A.M.; Laban, S.E.; Abdel-Sabour, M.A.; Nasr, S.A.E.; Zaki, M.M.; Shabbir, M.Z.; et al. Emergence and genetic analysis of variant pathogenic 4/91 (serotype 793/B) infectious bronchitis virus in Egypt during 2019. Virus Genes 2019, 55, 720–725. [Google Scholar] [CrossRef]
- Madbouly, H.M.; Abdel-Moneim, A.S.; Gelb, J.J.; Landman, B.S. Molecular characterization of three Egyptian isolates of infectious bronchitis virus. Vet. Med. J. Giza 2002, 50, 1053–1064. [Google Scholar]
- Abdel-Moneim, A.S.; Madbouly, H.M.; Gelb, J., Jr.; Ladman, B.S. Isolation and identification of Egypt/Beni-Suef/01 a novel genotype of infectious bronchitis virus. Vet. Med. J. Giza 2002, 50, 1065–1078. [Google Scholar]
- Meir, R.; Rosenblut, E.; Perl, S.; Kass, N.; Ayali, G.; Perk, S.; Hemsani, E. Identification of a novel nephropathogenic infectious bronchitis virus in Israel. Avian Dis. 2004, 48, 635–641. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Moneim, A.S.; El-Kady, M.F.; Ladman, B.S.; Gelb, J. S1 gene sequence analysis of a nephropathogenic strain of avian infectious bronchitis virus in Egypt. Virol. J. 2006, 3, 78. [Google Scholar] [CrossRef] [PubMed]
- Zanaty, A.; Arafa, A.S.; Hagag, N.; El-Kady, M. Genotyping and pathotyping of diversified strains of infectious bronchitis viruses circulating in Egypt. World J. Virol. 2016, 5, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Abozeid, H.H.; Paldurai, A.; Khattar, S.K.; Afifi, M.A.; El-Kady, M.F.; El-Deeb, A.H.; Samal, S.K. Complete genome sequences of two avian infectious bronchitis viruses isolated in Egypt: Evidence for genetic drift and genetic recombination in the circulating viruses. Infect. Genet. Evol. 2017, 53, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Selim, K.; Arafa, A.S.; Hussein, H.A.; El-Sanousi, A.A. Molecular characterization of infectious bronchitis viruses isolated from broiler and layer chicken farms in Egypt during 2012. Int. J. Vet. Sci. Med. 2013, 1, 102–108. [Google Scholar] [CrossRef]
- Abdel-Moneim, A.S.; Afifi, M.A.; El-Kady, M.F. Emergence of a novel genotype of avian infectious bronchitis virus in Egypt. Arch. Virol. 2012, 157, 2453–2457. [Google Scholar] [CrossRef]
- Zanaty, A.; Naguib, M.M.; El-Husseiny, M.H.; Mady, W.; Hagag, N.; Arafa, A.S. The sequence of the full spike S1 glycoprotein of infectious bronchitis virus circulating in Egypt reveals evidence of intra-genotypic recombination. Arch. Virol. 2016, 161, 3583–3587. [Google Scholar] [CrossRef]
- Moharam, I.; Sultan, H.; Hassan, K.; Ibrahim, M.; Shany, S.; Shehata, A.A.; Abo-ElKhair, M.; Pfaff, F.; Höper, D.; El Kady, M.; et al. Emerging infectious bronchitis virus (IBV) in Egypt: Evidence for an evolutionary advantage of a new S1 variant with a unique gene 3ab constellation. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2020, 85, 104433. [Google Scholar] [CrossRef]
- Sabra, M.; Abdellatif, W.; Ahmed, A.; Osman, N. Molecular characterization and phylogenetic analysis of full-length S1 gene of GI-16 and GI-23 infectious bronchitis virus in Qena, Egypt. J. World’s Poult. Res. 2020, 10, 71–80. [Google Scholar] [CrossRef]
- Abdel-Sabour, M.A.; Al-Ebshahy, E.M.; Khaliel, S.A.; Abdel-Wanis, N.A.; Yanai, T. Isolation and molecular characterization of novel infectious bronchitis virus variants from vaccinated broiler flocks in Egypt. Avian Dis. 2017, 61, 307–310. [Google Scholar] [CrossRef]
- Ghetas, A.M.; Kutkat, M.A.; Amer, M.M.; Awaad, M.H.H. Isolation and molecular identification of IBV isolates in different governorates in Egypt. J. Egypt. Soc. Parasitol. 2016, 46, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Lisowska, A.; Sajewicz-Krukowska, J.; Fusaro, A.; Pikula, A.; Domanska-Blicharz, K. First characterization of a Middle-East GI-23 lineage (Var2-like) of infectious bronchitis virus in Europe. Virus Res. 2017, 242, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Jiang, Y.; Low, S.; Wang, Z.; Nam, S.J.; Liu, W.; Kwang, J. Characterization of three infectious bronchitis virus isolates from China associated with proventriculus in vaccinated chickens. Avian Dis. 2001, 45, 416–424. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.W.; Jackwood, M.W. Evidence of genetic diversity generated by recombination among avian coronavirus IBV. Arch. Virol. 2000, 145, 2135–2148. [Google Scholar] [CrossRef]
- Jenkins, G.M.; Rambaut, A.; Pybus, O.G.; Holmes, E.C. Rates of molecular evolution in RNA viruses: A quantitative phylogenetic analysis. J. Mol. Evol. 2002, 54, 156–165. [Google Scholar] [CrossRef]
- Kusters, J.G.; Jager, E.J.; Lenstra, J.A.; Koch, G.; Posthumus, W.P.; Meloen, R.H.; van der Zeijst, B.A. Analysis of an immunodominant region of infectious bronchitis virus. J. Immunol. 1989, 143, 2692–2698. [Google Scholar]
- Kant, A.; Koch, G.; van Roozelaar, D.J.; Kusters, J.G.; Poelwijk, F.A.; van der Zeijst, B.A. Location of antigenic sites defined by neutralizing monoclonal antibodies on the S1 avian infectious bronchitis virus glycopolypeptide. J. Gen. Virol. 1992, 73 Pt 3, 591–596. [Google Scholar] [CrossRef]
- Cavanagh, D.; Davis, P.J.; Cook, J.K.; Li, D.; Kant, A.; Koch, G. Location of the amino acid differences in the S1 spike glycoprotein subunit of closely related serotypes of infectious bronchitis virus. Avian Pathol. 1992, 21, 33–43. [Google Scholar] [CrossRef]
- Pasternak, A.O.; Spaan, W.J.M.; Snijder, E.J. Nidovirus transcription: How to make sense…? J. Gen. Virol. 2006, 87, 1403–1421. [Google Scholar] [CrossRef]
- Simon-Loriere, E.; Holmes, E.C. Why do RNA viruses recombine? Nat. Rev. Microbiol. 2011, 9, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.M.; Jackwood, M.W. Molecular cloning and sequence comparison of the S1 glycoprotein of the Gray and JMK strains of avian infectious bronchitis virus. Virus Genes 1995, 9, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Thor, S.W.; Hilt, D.A.; Kissinger, J.C.; Paterson, A.H.; Jackwood, M.W. Recombination in avian gamma-coronavirus infectious bronchitis virus. Viruses 2011, 3, 1777–1799. [Google Scholar] [CrossRef]
- Naguib, M.M.; Höper, D.; Arafa, A.S.; Setta, A.M.; Abed, M.; Monne, I.; Beer, M.; Harder, T.C. Full genome sequence analysis of a newly emerged QX-like infectious bronchitis virus from Sudan reveals distinct spots of recombination. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2016, 46, 42–49. [Google Scholar] [CrossRef]
- Kiss, I.; Mató, T.; Homonnay, Z.; Tatár-Kis, T.; Palya, V. Successive occurrence of recombinant infectious bronchitis virus strains in restricted area of Middle East. Virus Evol. 2016, 2. [Google Scholar] [CrossRef]
- Ali, A.; Kilany, W.H.; Zain El-Abideen, M.A.; Sayed, M.E.; Elkady, M. Safety and efficacy of attenuated classic and variant 2 infectious bronchitis virus candidate vaccines. Poult. Sci. 2018, 97, 4238–4244. [Google Scholar] [CrossRef]
- Sultan, H.A.; Ali, A.; El Feil, W.K.; Bazid, A.H.I.; Zain El-Abideen, M.A.; Kilany, W.H. Protective efficacy of different live attenuated infectious bronchitis virus vaccination regimes against challenge with IBV variant-2 circulating in the Middle East. Front. Vet. Sci. 2019, 6, 341. [Google Scholar] [CrossRef]
- Cook, J.K.A.; Orbell, S.J.; Woods, M.A.; Huggins, M.B. Breadth of protection of the respiratory tract provided by different live-attenuated infectious bronchitis vaccines against challenge with infectious bronchitis viruses of heterologous serotypes. Avian Pathol. 1999, 28. [Google Scholar] [CrossRef]
- Cavanagh, D.; Naqi, S.A. Infectious Bronchitis. Dis. Poult. 2003, 11, 101–119. [Google Scholar] [CrossRef]
- Hassan, K.E.; Shany, S.A.S.; Ali, A.; Dahshan, A.-H.M.; El-Sawah, A.A.; El-Kady, M.F. Prevalence of avian respiratory viruses in broiler flocks in Egypt. Poult. Sci. 2016, 95, 1271–1280. [Google Scholar] [CrossRef]
- Abozeid, H.H.; Paldurai, A.; Varghese, B.P.; Khattar, S.K.; Afifi, M.A.; Zouelfakkar, S.; El-Deeb, A.H.; El-Kady, M.F.; Samal, S.K. Development of a recombinant Newcastle disease virus-vectored vaccine for infectious bronchitis virus variant strains circulating in Egypt. Vet. Res. 2019, 50, 12. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.T.; Wu, H.Y.; Wang, C.H. Genetic sequence changes related to the attenuation of avian infectious bronchitis virus strain TW2575/98. Virus Genes 2020, 56, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Jackwood, M.W.; Hall, D.; Handel, A. Molecular evolution and emergence of avian gammacoronaviruses. Infect. Genet. Evol. 2012, 12, 1305. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abozeid, H.H.; Naguib, M.M. Infectious Bronchitis Virus in Egypt: Genetic Diversity and Vaccination Strategies. Vet. Sci. 2020, 7, 204. https://doi.org/10.3390/vetsci7040204
Abozeid HH, Naguib MM. Infectious Bronchitis Virus in Egypt: Genetic Diversity and Vaccination Strategies. Veterinary Sciences. 2020; 7(4):204. https://doi.org/10.3390/vetsci7040204
Chicago/Turabian StyleAbozeid, Hassanein H., and Mahmoud M. Naguib. 2020. "Infectious Bronchitis Virus in Egypt: Genetic Diversity and Vaccination Strategies" Veterinary Sciences 7, no. 4: 204. https://doi.org/10.3390/vetsci7040204
APA StyleAbozeid, H. H., & Naguib, M. M. (2020). Infectious Bronchitis Virus in Egypt: Genetic Diversity and Vaccination Strategies. Veterinary Sciences, 7(4), 204. https://doi.org/10.3390/vetsci7040204