Serological Evidence of Avian Influenza in Captive Wild Birds in a Zoo and Two Safari Parks in Bangladesh

 ,
,  ,
,  ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
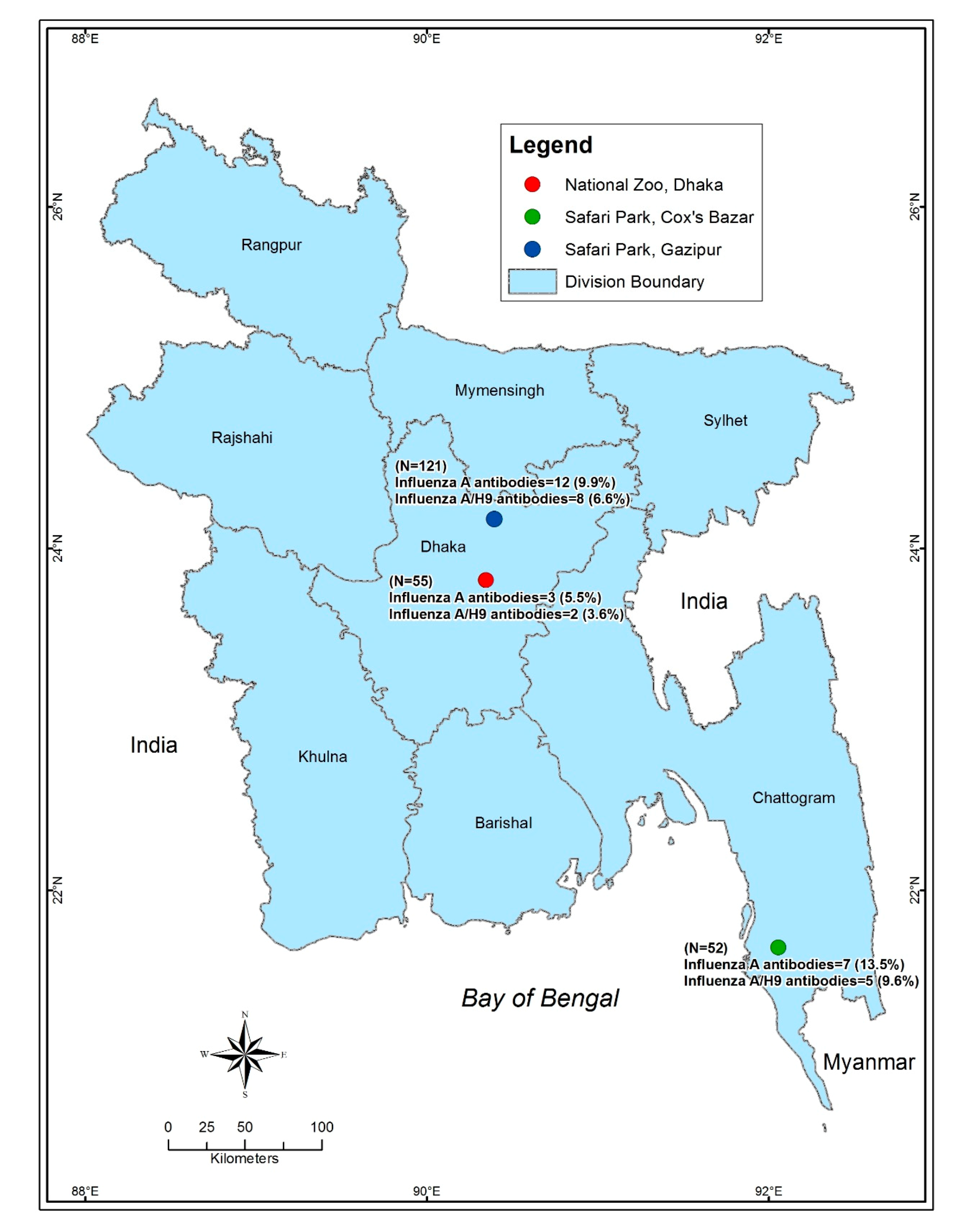
2.1. Study Location
2.2. Ethical Approval
2.3. Sample Collection
2.4. Competitive Enzyme-Linked Immunosorbent Assay (c-ELISA)
2.5. Hemagglutination Inhibition (HI) Test
2.6. Real-Time Reverse Transcription-Polymerase Chain Reaction (rRT-PCR)
2.7. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Swayne, D.; Akey, B. Avian influenza control strategies in the United States of America. Avian Influenza Prev. Control 2005, 8, 113. [Google Scholar]
- Webster, R.G.; Bean, W.J.; Gorman, O.T.; Chambers, T.M.; Kawaoka, Y. Evolution and ecology of influenza A viruses. Microbiol. Mol. Biol. Rev. 1992, 56, 152–179. [Google Scholar] [CrossRef]
- Horimoto, T.; Kawaoka, Y. Pandemic threat posed by avian influenza A viruses. Clin. Microbiol. Rev. 2001, 14, 129–149. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Liu, J. H9N2 influenza virus in China: A cause of concern. Protein Cell 2015, 6, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-H.; Song, C.-S. H9N2 avian influenza virus in Korea: Evolution and vaccination. Clin. Exp. Vaccine Res. 2013, 2, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Naeem, K.; Ullah, A.; Manvell, R.; Alexander, D. Avian Influenza A Subtype H9N2 in Poultry in Pakistan. Vet Rec. 1999, 145, 560. [Google Scholar] [CrossRef]
- Nili, H.; Asasi, K. Avian influenza (H9N2) outbreak in Iran. Avian Dis. 2003, 47, 828–831. [Google Scholar] [CrossRef]
- Gerloff, N.A.; Khan, S.U.; Zanders, N.; Balish, A.; Haider, N.; Islam, A.; Chowdhury, S.; Rahman, M.Z.; Haque, A.; Hosseini, P. Genetically diverse low pathogenicity avian influenza A virus subtypes co-circulate among poultry in Bangladesh. PLoS ONE 2016, 11, e0152131. [Google Scholar] [CrossRef]
- Shanmuganatham, K.; Feeroz, M.M.; Jones-Engel, L.; Walker, D.; Alam, S.; Hasan, M.; McKenzie, P.; Krauss, S.; Webby, R.J.; Webster, R.G. Genesis of avian influenza H9N2 in Bangladesh. Emerg. Microbes Infect. 2014, 3, e88. [Google Scholar] [CrossRef]
- Ansari, W.K.; Parvej, M.S.; El Zowalaty, M.E.; Jackson, S.; Bustin, S.A.; Ibrahim, A.K.; El Zowalaty, A.E.; Rahman, M.T.; Zhang, H.; Khan, M.F.R. Surveillance, epidemiological, and virological detection of highly pathogenic H5N1 avian influenza viruses in duck and poultry from Bangladesh. Vet. Microbiol. 2016, 193, 49–59. [Google Scholar] [CrossRef][Green Version]
- Hassan, M.M.; Hoque, M.A.; Debnath, N.C.; Yamage, M.; Klaassen, M. Are poultry or wild birds the main reservoirs for avian influenza in Bangladesh? Ecohealth 2017, 14, 490–500. [Google Scholar] [CrossRef]
- Hassan, M.M. Who Is the Culprit: Ecology and Epidemiology of Avian Influenza at the Wildlife-Poultry Interface in Bangladesh; Deakin Univeristy: Geelong, VIC, Australia, 2017. [Google Scholar]
- Marinova-Petkova, A.; Shanmuganatham, K.; Feeroz, M.M.; Jones-Engel, L.; Hasan, M.K.; Akhtar, S.; Turner, J.; Walker, D.; Seiler, P.; Franks, J. The continuing evolution of H5N1 and H9N2 influenza viruses in Bangladesh between 2013 and 2014. Avian Dis. 2016, 60, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Hossen, A.; Ahsan, M.; Kamruzzaman, M. Ecotourism Potentiality in the Dulahazra Safari Park, Bangladesh. J. Environ. Sci. Nat. Resour. 2014, 7, 189–196. [Google Scholar] [CrossRef]
- Desvaux, S.; Marx, N.; Ong, S.; Gaidet, N.; Hunt, M.; Manuguerra, J.-C.; Sorn, S.; Peiris, M.; Van der Werf, S.; Reynes, J.-M. Highly pathogenic avian influenza virus (H5N1) outbreak in captive wild birds and cats, Cambodia. Emerg. Infect. Dis. 2009, 15, 475. [Google Scholar] [CrossRef] [PubMed]
- Philippa, J.; Baas, C.; Beyer, W.; Bestebroer, T.; Fouchier, R.; Smith, D.; Schaftenaar, W.; Osterhaus, A. Vaccination against highly pathogenic avian influenza H5N1 virus in zoos using an adjuvanted inactivated H5N2 vaccine. Vaccine 2007, 25, 3800–3808. [Google Scholar] [CrossRef]
- Soilemetzidou, S.-E.; Greenwood, A.D.; Czirják, G.Á. Very low influenza a virus prevalence in cervids in german national parks. J. Zoo Wildl. Med. 2018, 49, 252–254. [Google Scholar] [CrossRef]
- Bertelsen, M.F.; Klausen, J.; Holm, E.; Grøndahl, C.; Jørgensen, P.H. Serological response to vaccination against avian influenza in zoo-birds using an inactivated H5N9 vaccine. Vaccine 2007, 25, 4345–4349. [Google Scholar] [CrossRef]
- Sangkachai, N.; Thongdee, M.; Chaiwattanarungruengpaisan, S.; Buddhirongawatr, R.; Chamsai, T.; Poltep, K.; Wiriyarat, W.; Paungpin, W. Serological evidence of influenza virus infection in captive wild felids, Thailand. J. Vet. Med Sci. 2019, 81, 1341–1347. [Google Scholar] [CrossRef]
- Swayne, D.E. Avian Influenza; John Wiley & Sons: Hoboken, NJ, USA, 2009. [Google Scholar]
- FAO. Biosecurity for Highly Pathogenic Avian Influenza; FAO: Rome, Italy, 2009; p. 17. [Google Scholar]
- Biswas, P.K.; Islam, M.Z.; Debnath, N.C.; Yamage, M. Modeling and Roles of Meteorological Factors in Outbreaks of Highly Pathogenic Avian Influenza H5N1. PLoS ONE 2014, 9, e98471. [Google Scholar] [CrossRef]
- Druce, J.; Garcia, K.; Tran, T.; Papadakis, G.; Birch, C. Evaluation of Swabs, Transport Media, and Specimen Transport Conditions for Optimal Detection of Viruses by PCR. J. Clin. Microbiol. 2012, 50, 1064–1065. [Google Scholar] [CrossRef]
- Basler, C.F.; García-Sastre, A.; Palese, P. Mutation of Neuraminidase Cysteine Residues Yields Temperature-Sensitive Influenza Viruses. J. Virol. 1999, 73, 8095–8103. [Google Scholar] [CrossRef] [PubMed]
- World Organisation For Animal Health (OIE). Manual of Diagnostic Tests and Vaccines for Terrestrial Animals 2018; OIE: Paris, France, 2018. [Google Scholar]
- Selleck, P. Influenza A Virus: A Competitive ELISA for the Detection of Antibodies to Influenza A Viruses in Equine Sera, Equine Influenza c-ELISA Protocol; Australian Animal Health Laboratory, Disease Diagnostic Project CSIRO Animal Health Laboratory: Geelong, Australia, 2007. [Google Scholar]
- AAHL. Molecular Diagnosis at CSIRO Animal, Food and Health Science-Nucleic Acid Detection for Disease Diagnosis & Emergency Disease Investigation-H5, H9 TaqManR Assays-AAHL Regional Programe; CSIRO: Geelong, Victoria, Australia, 2014. [Google Scholar]
- Heine, H.G.; Trinidad, L.; Selleck, P.; Lowther, S. Rapid Detection of Highly Pathogenic Avian Influenza H5N1 Virus by TaqMan Reverse Transcriptase–Polymerase Chain Reaction. Avian Dis. 2007, 51, 370–372. [Google Scholar] [CrossRef] [PubMed]
- Daodu, O.; Jegede, H.; Aiyedun, J.; Oludairo, O.; Olorunshola, I.; Daodu, O.; Ajadi, A.; Ambali, S. Surveillance for avian influenza virus in captive wild birds and indigenous chickens in Nigeria. Trop. Anim. Health Prod. 2020, 14, 1909–1912. [Google Scholar] [CrossRef] [PubMed]
- Nolting, J.M.; Dennis, P.; Long, L.; Holtvoigt, L.; Brown, D.; King, M.J.; Shellbarger, W.; Hanley, C.; Killian, M.L.; Slemons, R.D. Low pathogenic influenza A virus activity at avian interfaces in Ohio zoos, 2006–2009. Avian Dis. 2013, 57, 657–662. [Google Scholar] [CrossRef]
- Li, Y.; Liu, L.; Zhang, Y.; Duan, Z.; Tian, G.; Zeng, X.; Shi, J.; Zhang, L.; Chen, H. New avian influenza virus (H5N1) in wild birds, Qinghai, China. Emerg. Infect. Dis. 2011, 17, 265–267. [Google Scholar] [CrossRef]
- Hassan, M.M.; El Zowalaty, M.E.; Islam, A.; Khan, S.A.; Rahman, M.K.; Järhult, J.D.; Hoque, M.A. Prevalence and Diversity of Avian Influenza Virus Hemagglutinin Sero-Subtypes in Poultry and Wild Birds in Bangladesh. Vet. Sci. 2020, 7, 73. [Google Scholar] [CrossRef]
- Fallah Mehrabadi, M.; Ghalyanchi Langeroudi, A.; Bahonar, A.; Rabiee, M.; Tehrani, F.; Amirhajloo, S.; Steneroden, K.; Salman, M. Prevalence of Avian Influenza in live bird markets, bird gardens, and zoos in Iran in 2015: A cross-sectional study. Arch. Razi Inst. 2019, 74, 243–250. [Google Scholar]
- Nagarajan, S.; Kumar, M.; Murugkar, H.V.; Tripathi, S.; Shukla, S.; Agarwal, S.; Dubey, G.; Nagi, R.S.; Singh, V.P.; Tosh, C. Novel reassortant highly pathogenic avian influenza (H5N8) virus in zoos, India. Emerg. Infect. Dis. 2017, 23, 717. [Google Scholar] [CrossRef]
- Koethe, S.; Ulrich, L.; Ulrich, R.; Amler, S.; Graaf, A.; Harder, T.C.; Grund, C.; Mettenleiter, T.C.; Conraths, F.J.; Beer, M. Modulation of lethal HPAIV H5N8 clade 2.3. 4.4 B infection in AIV pre-exposed mallards. Emerg. Microbes Infect. 2020, 9, 180–193. [Google Scholar] [CrossRef]
- Zaman, R.U.; Alamgir, A.; Rahman, M.; Azziz-Baumgartner, E.; Gurley, E.S.; Sharker, M.A.Y.; Brooks, W.A.; Azim, T.; Fry, A.M.; Lindstrom, S. Influenza in outpatient ILI case-patients in national hospital-based surveillance, Bangladesh, 2007–2008. PLoS ONE 2009, 4, e8452. [Google Scholar] [CrossRef]
- Khan, S.U.; Berman, L.; Haider, N.; Gerloff, N.; Rahman, M.Z.; Shu, B.; Rahman, M.; Dey, T.K.; Davis, T.C.; Das, B.C. Investigating a crow die-off in January–February 2011 during the introduction of a new clade of highly pathogenic avian influenza virus H5N1 into Bangladesh. Arch. Virol. 2014, 159, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.M.; Hoque, M.A.; Ujvari, B.; Klaassen, M. Live bird markets in Bangladesh as a potentially important source for Avian Influenza Virus transmission. Prev. Vet. Med. 2018, 156, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Shinde, P.V.; Koratkar, S.S.; Pawar, S.D.; Kale, S.D.; Rawankar, A.S.; Mishra, A.C. Serologic Evidence of Avian Influenza H9N2 and Paramyxovirus Type 1 Infectionin Emus (Dromaius novaehollandiae) in India. Avian Dis. 2012, 56, 257–260. [Google Scholar] [CrossRef] [PubMed]
- Humberd, J.; Guan, Y.; Webster, R.G. Comparison of the replication of influenza A viruses in Chinese ring-necked pheasants and chukar partridges. J. Virol. 2006, 80, 2151–2161. [Google Scholar] [CrossRef]
- Humberd, J.; Boyd, K.; Webster, R.G. Emergence of influenza A virus variants after prolonged shedding from pheasants. J. Virol. 2007, 81, 4044–4051. [Google Scholar] [CrossRef]


{kind=link}
| Variable | Categories (N) | n (%) | 95% Confidence Interval (CI) | p |
|---|---|---|---|---|
| Year | 2013 (75) | 10 (13.3) | 6.6–23.2 | 0.187 |
| 2014 (153) | 12 (7.8) | 4.1–13.3 | ||
| Month | January (23) | 1 (4.4) | 0.1–21.9 | 0.332 |
| February (130) | 11 (8.5) | 4.3–14.6 | ||
| November (61) | 7 (11.5) | 4.7–22.2 | ||
| December (14) | 3 (21.4) | 4.7–50.8 | ||
| Site | National Zoo, Dhaka (55) | 3 (5.5) | 1.1–15.1 | 0.370 |
| Safari Park, Cox’s Bazar (52) | 7 (13.5) | 5.6–25.8 | ||
| Safari Park, Gazipur (121) | 12 (9.9) | 5.2–16.7 | ||
| Species | Emu (23) | 6 (26.1) | 10.2–48.4 | 0.011 |
| Golden pheasant (44) | 8 (18.2) | 8.2–32.7 | ||
| Green peafowl (32) | 1 (3.1) | 0.07–16.2 | ||
| Guinea fowl (37) | 4 (10.8) | 3.0–25.4 | ||
| Macaw (30) | - | - | ||
| Ostrich (17) | - | - | ||
| Raj dhonesh (22) | 1 (4.6) | 0.1–22.8 | ||
| Red-naped ibis (23) | 2 (8.7) | 1.1–28.1 |
| Variable | Categories (N) | n (%) | 95% Confidence Interval (CI) | p |
|---|---|---|---|---|
| Year | 2013 (75) | 7 (9.3) | 3.8–18.3 | 0.24 |
| 2014 (153) | 8 (5.2) | 2.3–10.0 | ||
| Month | January (23) | 1 (4.4) | 0.1–21.9 | 0.138 |
| February (130) | 7 (5.4) | 2.2–10.8 | ||
| November (61) | 4 (6.6) | 1.8–15.9 | ||
| December (14) | 3 (21.4) | 4.6–50.8 | ||
| Site | National Zoo, Dhaka (55) | 2 (3.6) | 0.4–12.5 | 0.460 |
| Safari Park, Cox’s Bazar (52) | 5 (9.6) | 3.2–21.0 | ||
| Safari Park, Gazipur (121) | 8 (6.6) | 2.9–12.6 | ||
| Species | Emu (23) | 4 (17.4) | 4.9–38.8 | 0.107 |
| Golden pheasant (44) | 5 (11.4) | 3.8–24.6 | ||
| Green peafowl (32) | 1 (3.1) | 0.07–16.2 | ||
| Guinea fowl (37) | 3 (8.1) | 1.7–21.9 | ||
| Macaw (30) | - | - | ||
| Ostrich (17) | - | - | ||
| Raj dhonesh (22) | - | - | ||
| Red-naped ibis (23) | 2 (8.7) | 1.1–28.0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hassan, M.M.; El Zowalaty, M.E.; Islam, A.; Rahman, M.M.; Chowdhury, M.N.U.; Nine, H.S.M.Z.; Rahman, M.K.; Järhult, J.D.; Hoque, M.A. Serological Evidence of Avian Influenza in Captive Wild Birds in a Zoo and Two Safari Parks in Bangladesh. Vet. Sci. 2020, 7, 122. https://doi.org/10.3390/vetsci7030122
Hassan MM, El Zowalaty ME, Islam A, Rahman MM, Chowdhury MNU, Nine HSMZ, Rahman MK, Järhult JD, Hoque MA. Serological Evidence of Avian Influenza in Captive Wild Birds in a Zoo and Two Safari Parks in Bangladesh. Veterinary Sciences. 2020; 7(3):122. https://doi.org/10.3390/vetsci7030122
Chicago/Turabian StyleHassan, Mohammad M., Mohamed E. El Zowalaty, Ariful Islam, Md. M. Rahman, Md. N. U. Chowdhury, Hatem S. M. Z. Nine, Md. K. Rahman, Josef D. Järhult, and Md. A. Hoque. 2020. "Serological Evidence of Avian Influenza in Captive Wild Birds in a Zoo and Two Safari Parks in Bangladesh" Veterinary Sciences 7, no. 3: 122. https://doi.org/10.3390/vetsci7030122
APA StyleHassan, M. M., El Zowalaty, M. E., Islam, A., Rahman, M. M., Chowdhury, M. N. U., Nine, H. S. M. Z., Rahman, M. K., Järhult, J. D., & Hoque, M. A. (2020). Serological Evidence of Avian Influenza in Captive Wild Birds in a Zoo and Two Safari Parks in Bangladesh. Veterinary Sciences, 7(3), 122. https://doi.org/10.3390/vetsci7030122