In Vitro Inhibiting Effects of Three Fungal Species on Eggs of Donkey Gastrointestinal Strongyles






{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Fungi
2.2. Sampling
2.3. Parasitological Analysis and Recovery, Suspension, and Purification of Gastrointestinal Strongyle (GIS) Eggs
2.4. Egg Hatch Test (EHT)
2.5. Identification of GIS Genera
2.6. Statistical Analysis
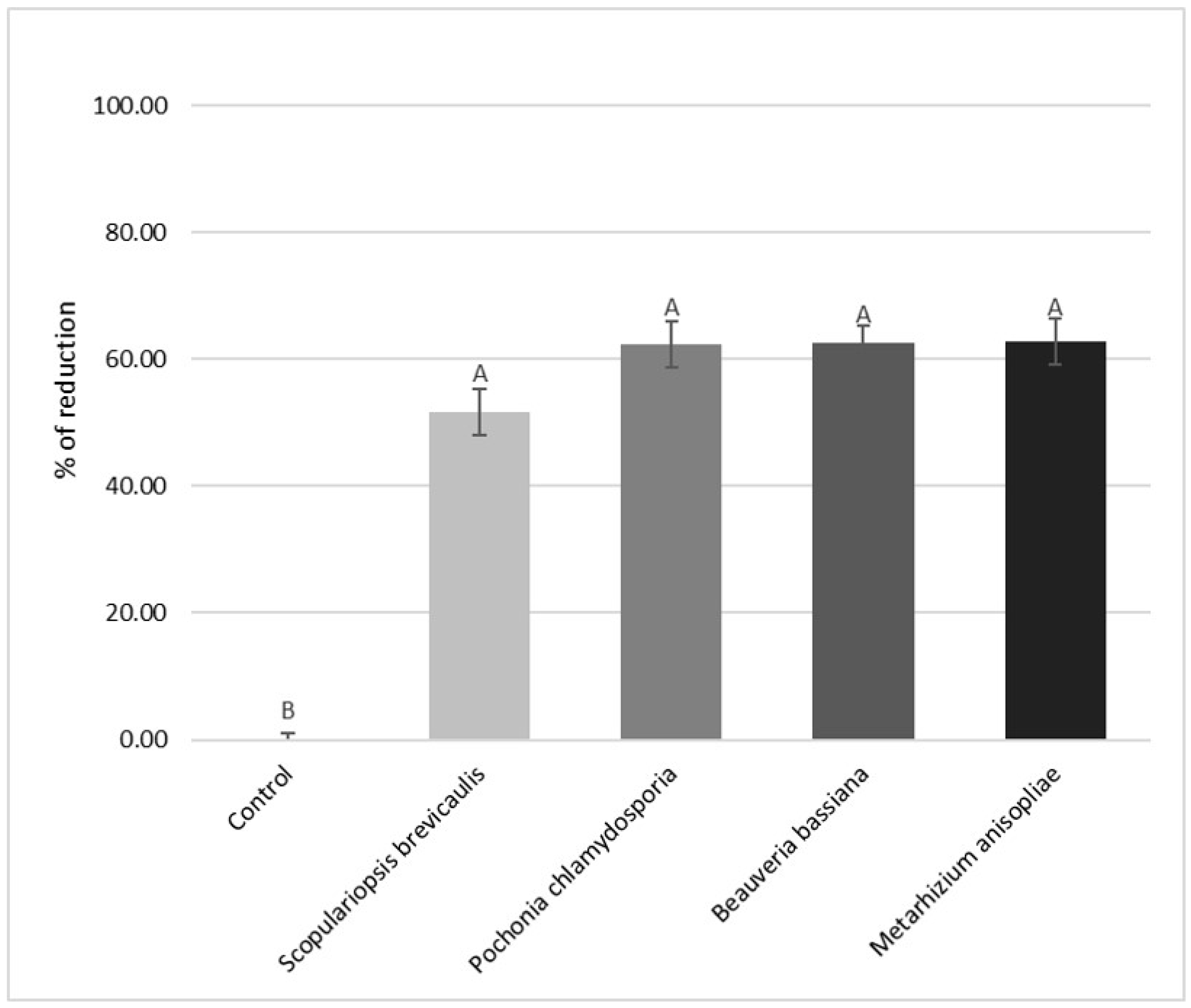
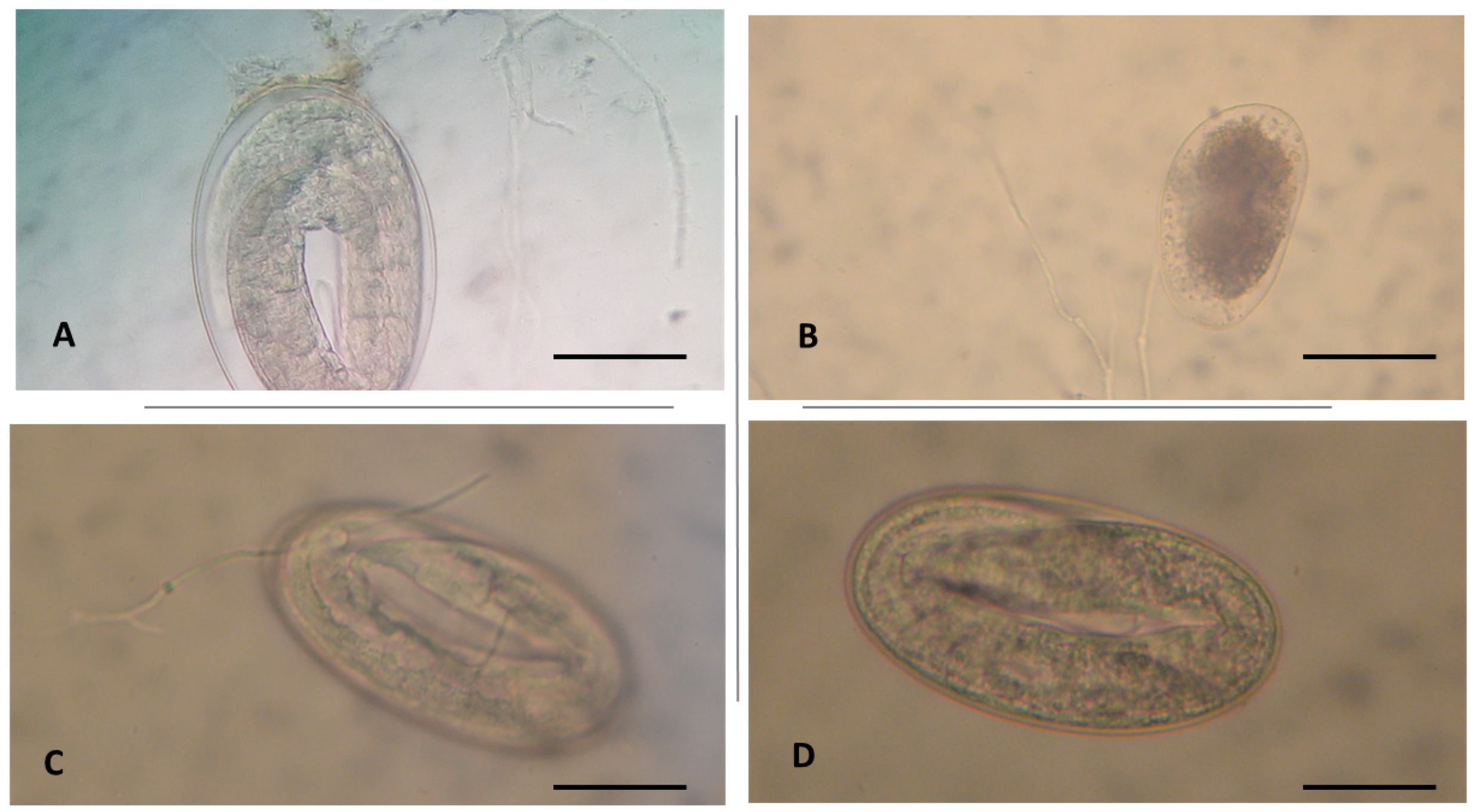
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Camillo, F.; Rota, A.; Biagini, L.; Tesi, M.; Fanelli, D.; Panzani, D. The Current Situation and Trend of Donkey Industry in Europe. Equine Vet. Sci. 2018, 65, 44–49. [Google Scholar] [CrossRef]
- Perinelli, D.R.; Bonacucina, G.; Cespi, M.; Bonazza, F.; Palmieri, G.F.; Pucciarelli, S.; Polzonetti, V.; Attarian, L.; Polidori, P.; Vincenzetti, S. A comparison among β-caseins purified from milk of different species: Selfassembling behaviour and immunogenicity potential. Colloids Surf. B 2019, 173, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Souroullas, K.; Aspri, M.; Papademas, P. Donkey milk as a supplement in infant formula: Benefits and technological challenges. Int. Food Res. J. 2018, 109, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Matthews, J.B.; Burden, F.A. Common helminth infections of donkeys and their control in temperate regions. Equine Vet. Educ. 2013, 25, 461–467. [Google Scholar] [CrossRef]
- Getachew, M.; Trawford, A.; Feseha, G.; Reid, S.W.J. Gastrointestinal parasites of working donkeys of Ethiopia. Trop. Anim. Health 2010, 42, 27–33. [Google Scholar] [CrossRef]
- Molento, M.B.; Canever, R.J. In vitro evaluation of ivermectin, moxidectin, albendazole and pyrantel against cyathostomins of horses. Braz. J. Vet. Parasitol. 2018, 27, 90–93. [Google Scholar] [CrossRef]
- Gokbulut, C.; McKellar, Q.A. Anthelmintic drugs used in equine species. Vet. Parasitol. 2018, 261, 27–52. [Google Scholar] [CrossRef]
- Gokbulut, C.; Aksit, D.; Smaldone, G.; Mariani, U.; Veneziano, V. Plasma pharmacokinetics, faecal excretion and efficacy of pyrantel pamoate paste and granule formulations following per os administration in donkeys naturally infected with intestinal strongilydae. Vet. Parasitol. 2014, 205, 186–192. [Google Scholar] [CrossRef]
- Gokbulut, C.; Aksit, D.; Santoro, M.; Roncoroni, C.; Mariani, U.; Buono, F.; Rufrano, D.; Fagiolo, A.; Veneziano, V. Plasma disposition, milk excretion and parasitological efficacy of mebendazole in donkeys naturally infected by Cyathostominae. Vet. Parasitol. 2016, 217, 95–100. [Google Scholar] [CrossRef]
- Corbett, C.J.; Love, S.; Moore, A.; Burden, F.A.; Matthews, J.B.; Denwood, M.J. The effectiveness of faecal removal methods of pasture management to control the cyathostomin burden of donkeys. Parasite Vectors 2014, 7, 48. [Google Scholar] [CrossRef]
- Hernández, J.Á.; Sánchez-Andrade, R.; Cazapal-Monteiro, C.F.; Arroyo, F.L.; Sanchís, J.M.; Paz-Silva, A.; Arias, M.S. A combined effort to avoid strongyle infection in horses in an oceanic climate region: Rotational grazing and parasiticidal fungi. Parasite Vectors 2018, 11, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Collas, C.; Sallé, G.; Dumont, B.; Cabaret, J.; Cortet, J.; Martin-Rosset, W.; Wimel, L.; Fleurance, G. Are sainfoin or protein supplements alternatives to control small strongyle infection in horses? Animal 2018, 12, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Peachey, L.E.; Pinchbeck, G.L.; Matthews, J.B.; Burden, F.A.; Mulugeta, G.; Scantlebury, C.E.; Hodgkinson, J.E. An evidence-based approach to the evaluation of ethnoveterinary medicines against strongyle nematodes of equids. Vet. Parasitol. 2015, 210, 40–52. [Google Scholar] [CrossRef]
- Silveira, W.F.; Braga, F.R.; Tavela, A.O.; Santos, L.F.; Reis, D.R.; Rocha Aguiar, A.; Magri Ferraz, C.; Carvalho, L.M.; de Hollanda Ayupe, T.; Cola Zanuncio, J.; et al. Nematophagous fungi combinations reduce free-living stages of sheep gastrointestinal nematodes in the field. J. Invertebr. Pathol. 2017, 150, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Braga, F.R.; De Araújo, J.V. Nematophagous fungi for biological control of gastrointestinal nematodes in domestic animals. Appl. Microbiol. Biotechnol. 2014, 98, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Aranda-Martinez, A.; Naranjo Ortiz, M.A.; Abihssira García, I.S.; Zavala-Gonzalez, E.A.; Lopez-Llorca, V.L. Ethanol production from chitosan by the nematophagous fungus Pochonia chlamydosporia and the entomopathogenic fungi Metarhizium anisopliae and Beauveria bassiana. Microbiol. Res. 2017, 204, 30–39. [Google Scholar] [CrossRef]
- Cai, J.; Yang, J.; Du, Y.; Fan, L.; Qiu, Y.; Li, J.; Kennedy, J.F. Purification and characterization of chitin deacetylase from Scopulariopsis brevicaulis. Carbohydr. Polym. 2006, 65, 211–217. [Google Scholar] [CrossRef]
- Mascarin, G.M.; Jaronski, S.T. The production and uses of Beauveria bassiana as a microbial insecticide. World J. Microbiol. Biotechnol. 2016, 32, 177. [Google Scholar] [CrossRef]
- Beys-da-Silva, W.O.; Rosa, R.L.; Berger, M.; Coutinho-Rodrigues, C.J.B.; Vainstein, M.H.; Schrank, A.; Bittencourt, V.R.E.P.; Santi, L. Updating the application of Metarhizium anisopliae to control cattle tick Rhipicephalus microplus (Acari: Ixodidae). Exp. Parasitol. 2020, 208, 107. [Google Scholar] [CrossRef]
- Braga, F.R.; Araújo, J.V.; Carvalho, R.O.; Silva, A.R.; Araujo, J.M.; Feitas Soares, F.E.; André Geniêr, H.L.; Ferreira, S.R.; Queiroz, J.H. Ovicidal action of a crude enzymatic extract of the fungus Pochonia chlamydosporia against cyathostomin eggs. Vet. Parasitol. 2010, 172, 264–268. [Google Scholar] [CrossRef]
- Vieira, I.S.; Oliveira, I.C.; Campos, A.K.; Araújo, J.V. Association and predatory capacity of fungi Pochonia chlamydosporia and Arthrobotrys cladodes in the biological control of parasitic helminths of bovines. Parasitology 2019, 146, 1347–1351. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.A.; Coop, R.L.; Wall, R.L. Veterinary Parasitology, 3rd ed.; Blackwell Publishing: Oxford, UK, 2007; p. 799. [Google Scholar]
- Coles, G.C.; Jackson, F.; Pomroy, W.; Prichard, R.K.; Von Samson-himmelstjerna, G.; Silvestre, A.; Taylor, M.A.; Vercruysse, J. The detection of anthelmintic resistance in nematodes of veterinary importance. Vet. Parasitol. 2006, 136, 167–185. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, C.O.; Chagas, A.C.; Cotinguiba, F.; Furlan, M.; Brito, L.G.; Chaves, F.C.M.; Stephan, M.P.; Bizzo, H.R.; Amarante, A.F.T. The anthelmintic effect of plant extracts on Haemonchus contortus and Strongyloides venezuelensis. Vet. Parasitol. 2012, 183, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Ministry of Agriculture, Fisheries and Food (MAFF). Manual of Veterinary Parasitological Laboratory Techniques; Her Majesty’s Stationary Office (HMSO): London, UK, 1986; pp. 1–152.
- Bevilaqua, C.M.L.; Rodrigues, M.L.; Concordet, D. Identification of infective larvae of some common nematode strongylids of horses. Revue de Méd. Vétérinaire 1993, 144, 989–995. [Google Scholar]
- Braga, F.R.; Araújo, J.V.; Araujo, J.M.; Frassy, L.N.; Tavela, A.O.; Soares, F.E.; Carvalho, R.O.; Queiroz, L.M.; Queiroz, J.H. Pochonia chlamydosporia fungal activity in a solid medium and its crude extract against eggs of Ascaridia galli. J. Helminthol. 2012, 86, 348–352. [Google Scholar] [CrossRef]
- Carvalho, R.O.; Araújo, J.V.; Braga, F.R.; Araujo, J.M.; Alves, C.D.F. Ovicidal activity of Pochonia chlamydosporia and Paecilomyces lilacinus on Toxocara canis eggs. Vet. Parasitol. 2010, 169, 123–127. [Google Scholar] [CrossRef]
- Ferreira, S.R.; Araújo, J.V.; Braga, F.R.; Araujo, J.M.; Carvalho, R.O.; Silva, A.R.; Frassy, L.N.; Freitas, L.G. Ovicidal activity of seven Pochonia chlamydosporia fungal isolates on Ascaris suum eggs. Trop. Anim. Health Prod. 2011, 43, 639–642. [Google Scholar] [CrossRef]
- Silva, A.R.; Araújo, J.V.; Braga, F.R.; Alves, C.D.F.; Frassy, L.N. In vitro ovicidal activity of the nematophagous fungi Duddingtonia flagrans, Monacrosporium thaumasium and Pochonia chlamydosporia on Trichuris vulpis eggs. Vet. Paras. 2010, 172, 76–79. [Google Scholar] [CrossRef]
- Luz, C.; Rocha, L.F.N.; Nery, G.V.; Magalhães, B.P.; Tigano, M.S. Activity of Oil-formulated Beauveria bassiana against Triatoma sordida in Peridomestic Areas in Central Brazil. Mem. Inst. Oswaldo Cruz. 2004, 99, 211–218. [Google Scholar] [CrossRef][Green Version]
- Mannino, M.C.; Huarte-Bonnet, C.; Davyt-Colo, B.; Pedrini, N. Is the Insect Cuticle the only Entry Gate for Fungal Infection? Insights into Alternative Modes of Action of Entomopathogenic Fungi. J. Fungi 2019, 5, 33. [Google Scholar] [CrossRef]
- Rehner, S.A.; Minnis, A.M.; Sung, G.H.; Luangsa-ard, J.J.; Devotto, L.; Humber, R.A. Phylogeny and systematics of the anamorphic, entomopathogenic genus Beauveria. Mycologia 2011, 103, 1055–1073. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.Q.; Zou, W.H.; Li, D.L.; Chen, J.T.; Huang, Q.; Zhou, L.J.; Tian, X.X.; Chen, Y.J.; Peng, H.J. Expression of Bacillus thuringiensis toxin Cyt2Ba in the entomopathogenic fungus Beauveria bassiana increases its virulence towards Aedes mosquitoes. PLoS Neglect. Trop. 2019, 13, 7. [Google Scholar] [CrossRef] [PubMed]
- Tupe, S.G.; Pathan, E.K.; Deshpande, M.V. Development of Metarhizium anisopliae as a Mycoinsecticide: From Isolation to Field Performance. J. Vis. Exp. 2017, 125. [Google Scholar] [CrossRef] [PubMed]
- Briggs, L.L.; Colwell, D.D.; Wall, R. Control of the cattle louse Bovicola bovis with the fungal pathogen Metarhizium anisopliae. Vet. Parasitol. 2006, 142, 3–4. [Google Scholar] [CrossRef] [PubMed]
- Brooks, A.J.; Wall, R. Infection of Psoroptes mites with the fungus Metarhizium anisopliae. Exp. Appl. Acarol. 2010, 25, 869–880. [Google Scholar] [CrossRef] [PubMed]
- Kordalewska, M.; Jagielski, T.; Brillowska-Dąbrowska, A. Rapid Assays for Specific Detection of Fungi of Scopulariopsis and Microascus Genera and Scopulariopsis brevicaulis Species. Mycopathologia 2016, 181, 465–474. [Google Scholar] [CrossRef][Green Version]
- Cawcutt, K.; Baddour, L.M.; Burgess, M.A. Case of Scopulariopsis brevicaulis endocarditis with mycotic aneurysm in an immunocompetent host. Med. Case Rep. 2015, 871–872. [Google Scholar] [CrossRef]
- Gupta, A.K.; Drummond-Main, C.; Cooper, E.A. Systematic review of nondermatophyte mold onychomycosis: Diagnosis, clinical types, epidemiology, and treatment. J. Am. Acad. Dermatol. 2012, 66, 494–502. [Google Scholar] [CrossRef]
- Lotery, A.J.; Kerr, J.R.; Page, B.A. Fungal keratitis caused by Scopulariopsis brevicaulis: Successful treatment with topical amphotericin B and chloramphenicol without the need for surgical debridement. Br. J. Ophthalmol. 1994, 78, 730. [Google Scholar] [CrossRef][Green Version]
- Perrucci, S.; Zini, A.; Donadio, E.; Mancianti, F.; Fichi, G. Isolation of Scopulariopsis spp. fungi from Psoroptes cuniculi body surface and evaluation of their entomopathogenic role. Parasitol. Res. 2008, 102, 957–962. [Google Scholar] [CrossRef]
- Braga, F.R.; Araújo, J.V.; Silva, A.R.; Carvalho, R.O.; Araujo, J.M.; Ferreira, S.R.; Carvalho, G.R. Viability of the nematophagous fungus Pochonia chlamydosporia after passage through the gastrointestinal tract of horses. Vet. Parasitol. 2010b, 168, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, L.M.; Braga, F.R.; Domingues, R.R.; Araujo, J.M.; Lelis, R.T.; Paula, A.T.; Silveira, W.F.; Araújo, J.V. Interaction of the nematophagous fungus Pochonia chlamydosporia and Parascaris equorum eggs in different culture media. J. Basic Microbiol. 2014, 54, 109–114. [Google Scholar] [CrossRef] [PubMed]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maestrini, M.; Nardoni, S.; Mancianti, F.; Mancini, S.; Perrucci, S. In Vitro Inhibiting Effects of Three Fungal Species on Eggs of Donkey Gastrointestinal Strongyles. Vet. Sci. 2020, 7, 53. https://doi.org/10.3390/vetsci7020053
Maestrini M, Nardoni S, Mancianti F, Mancini S, Perrucci S. In Vitro Inhibiting Effects of Three Fungal Species on Eggs of Donkey Gastrointestinal Strongyles. Veterinary Sciences. 2020; 7(2):53. https://doi.org/10.3390/vetsci7020053
Chicago/Turabian StyleMaestrini, Michela, Simona Nardoni, Francesca Mancianti, Simone Mancini, and Stefania Perrucci. 2020. "In Vitro Inhibiting Effects of Three Fungal Species on Eggs of Donkey Gastrointestinal Strongyles" Veterinary Sciences 7, no. 2: 53. https://doi.org/10.3390/vetsci7020053
APA StyleMaestrini, M., Nardoni, S., Mancianti, F., Mancini, S., & Perrucci, S. (2020). In Vitro Inhibiting Effects of Three Fungal Species on Eggs of Donkey Gastrointestinal Strongyles. Veterinary Sciences, 7(2), 53. https://doi.org/10.3390/vetsci7020053