
Pathobiology of Hemangiosarcoma in Dogs: Research Advances and Future Perspectives



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Historical Perspective of Canine HSA
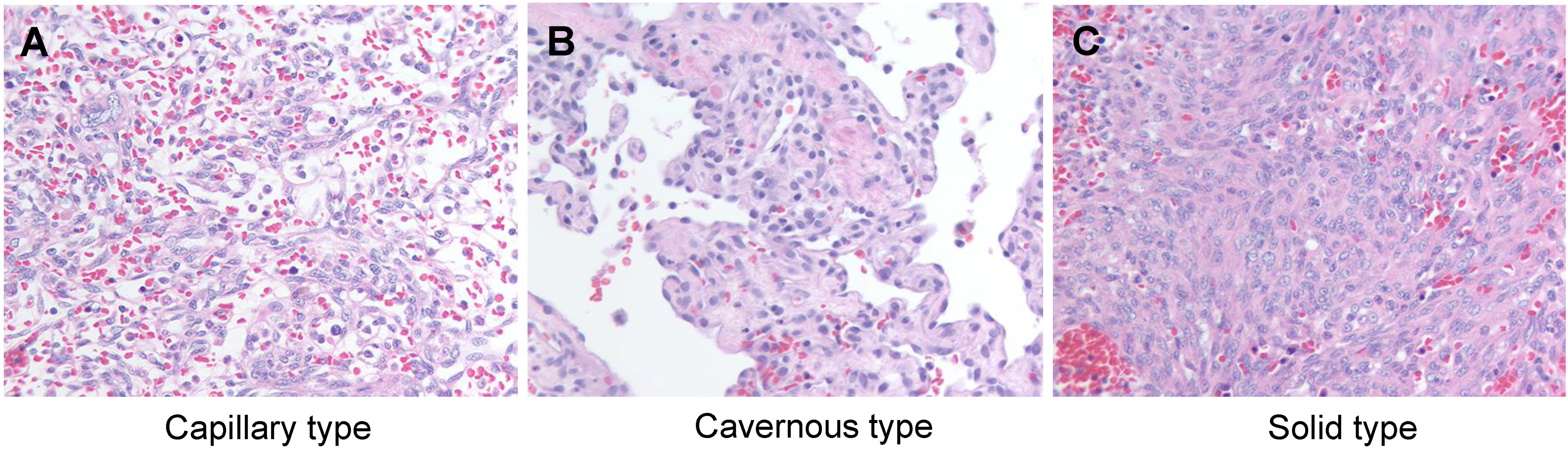
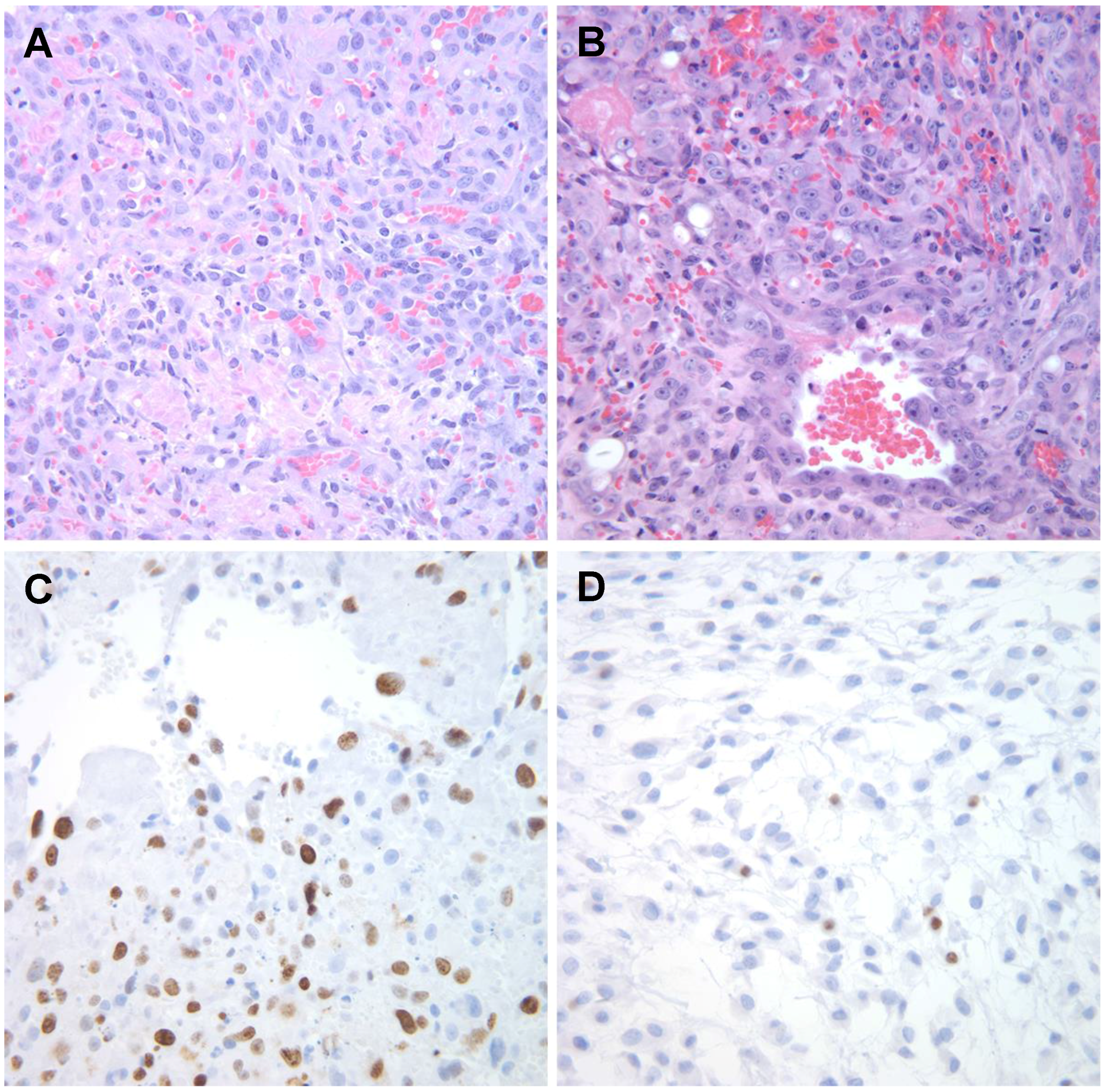
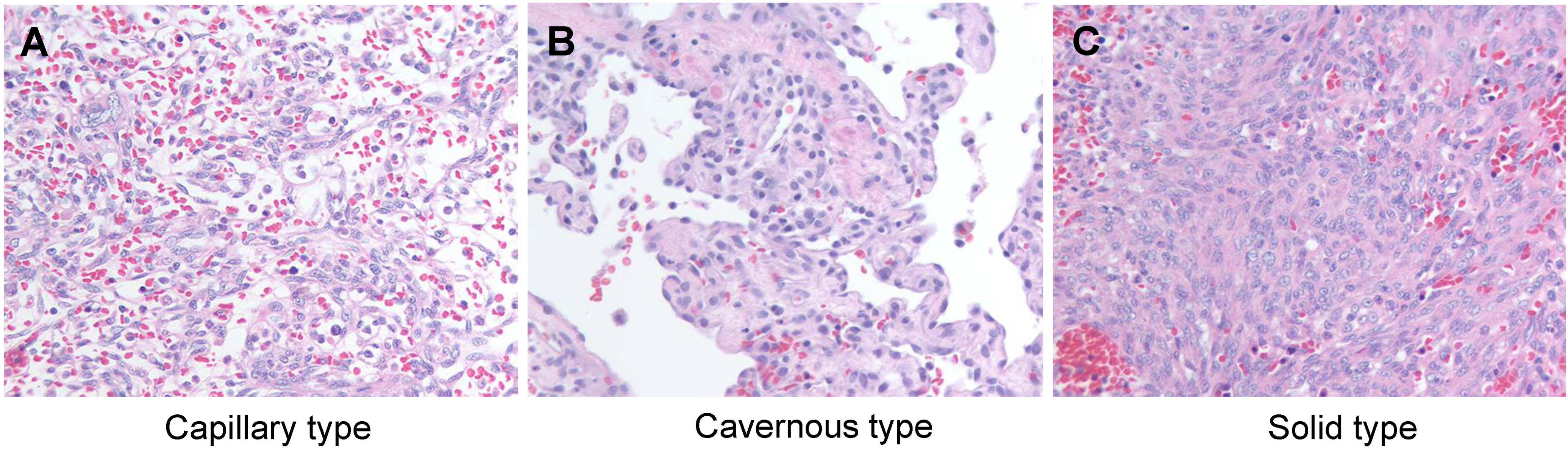
3. Classical Pathology of Canine HSA
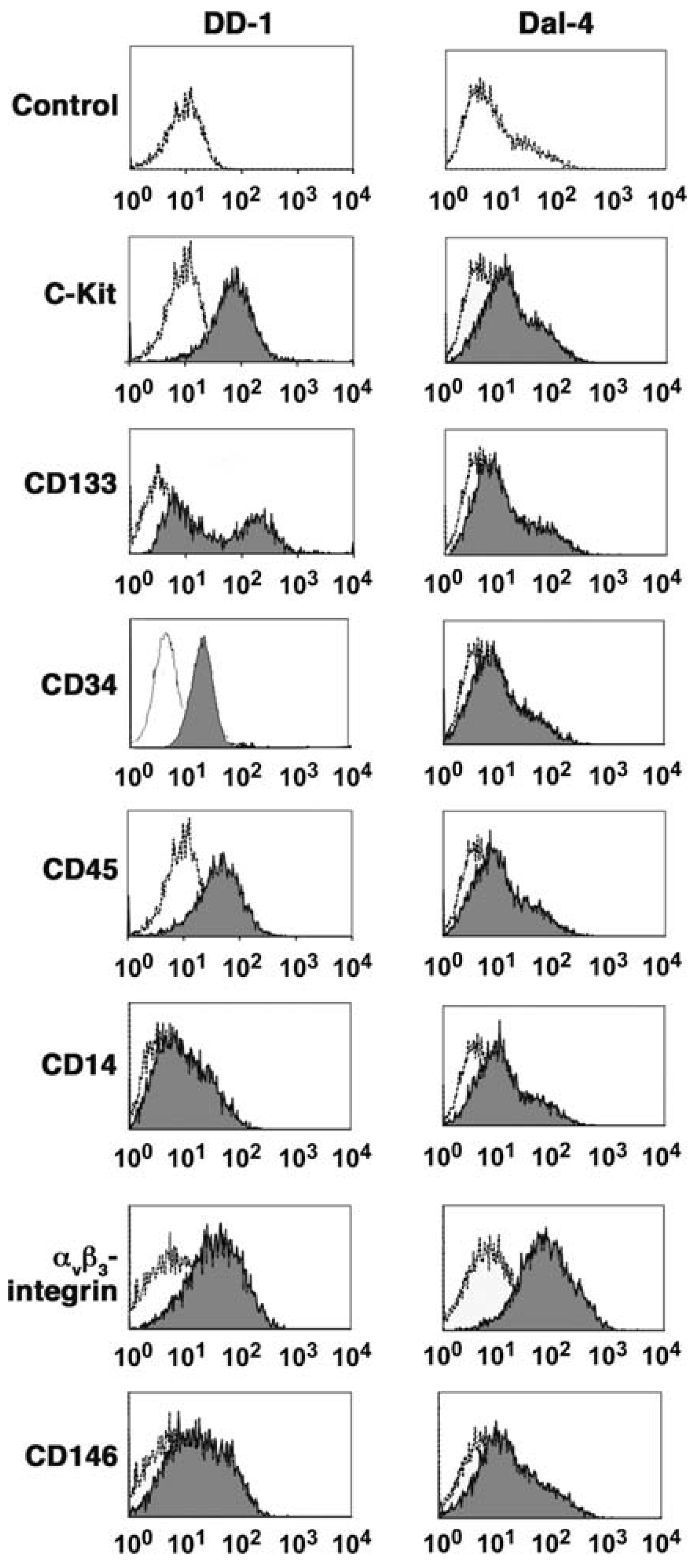
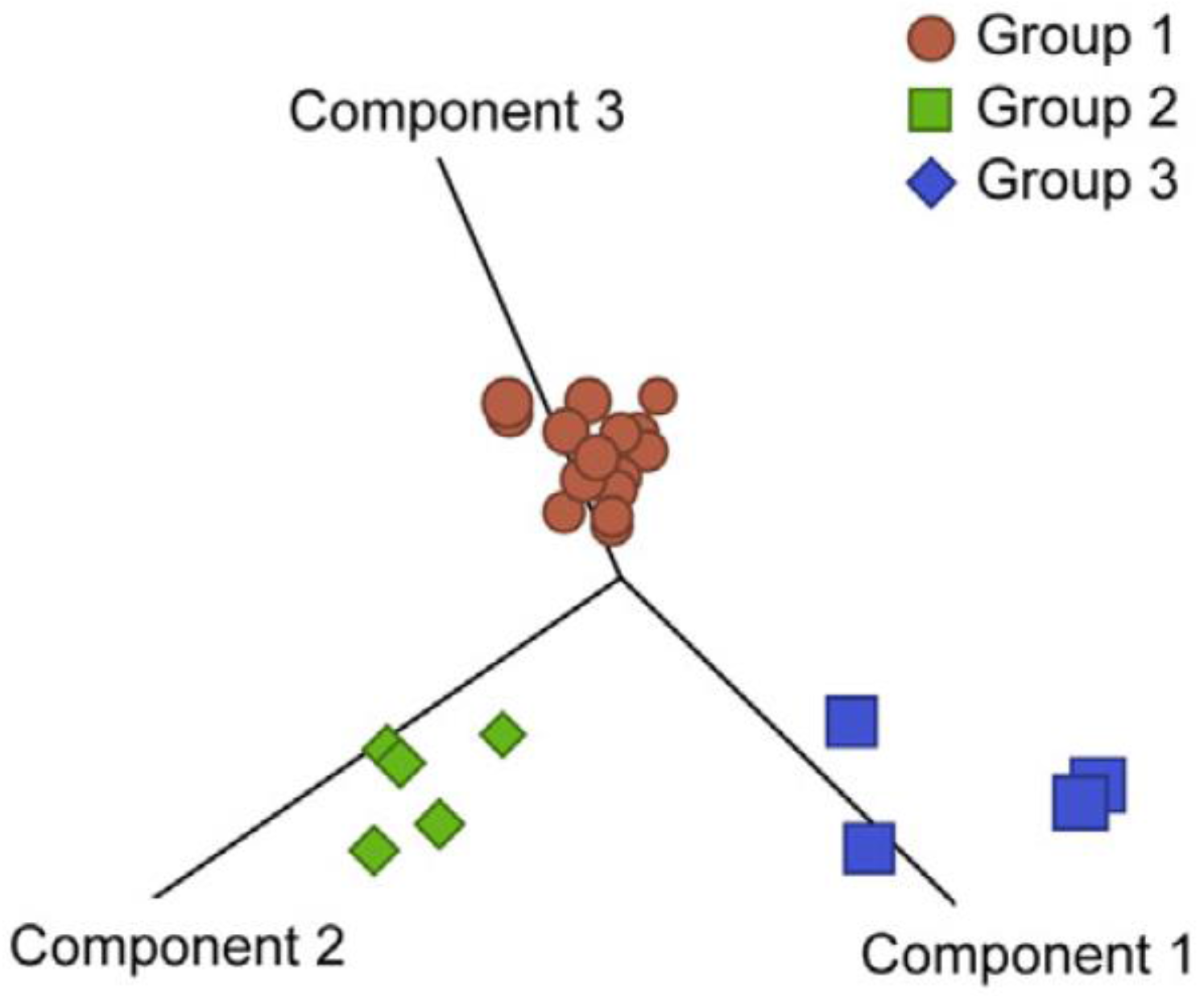
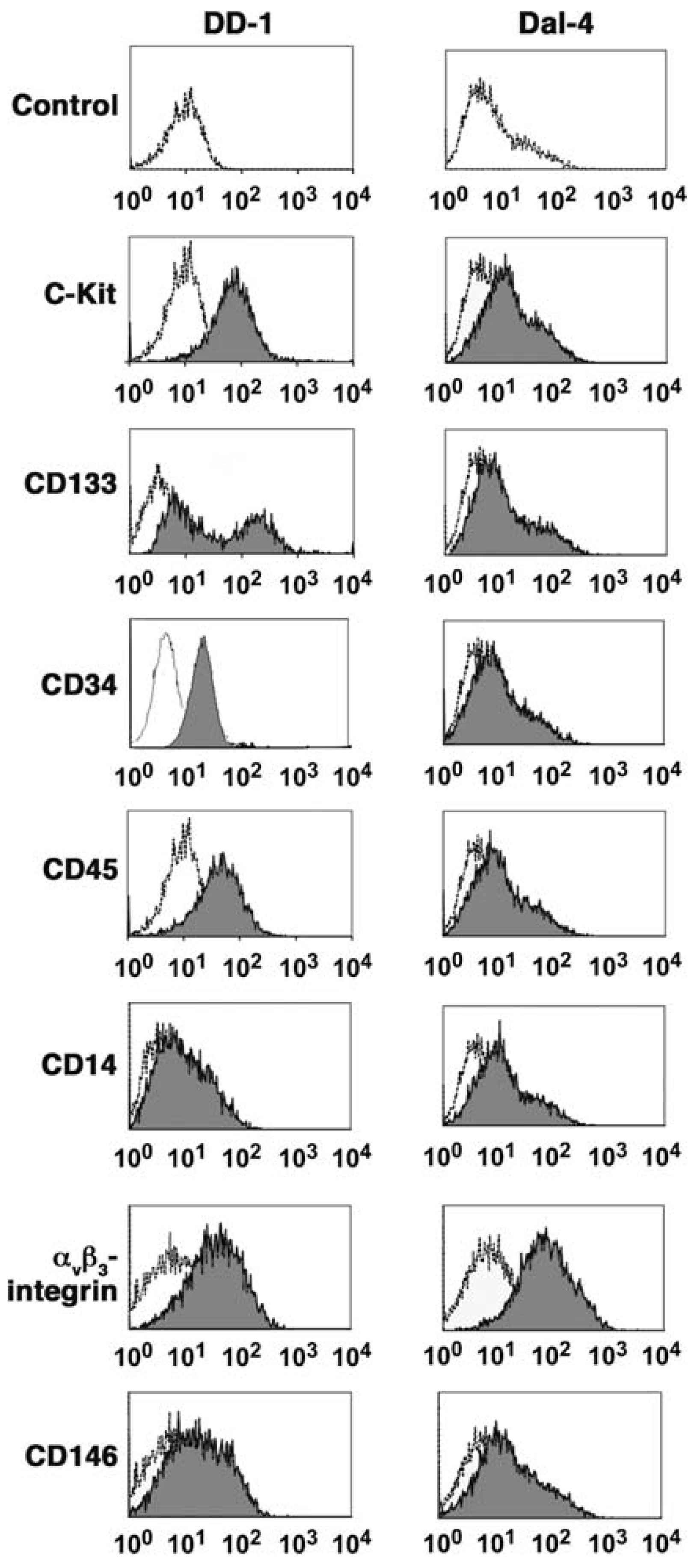
4. Ontogeny of Canine HSA
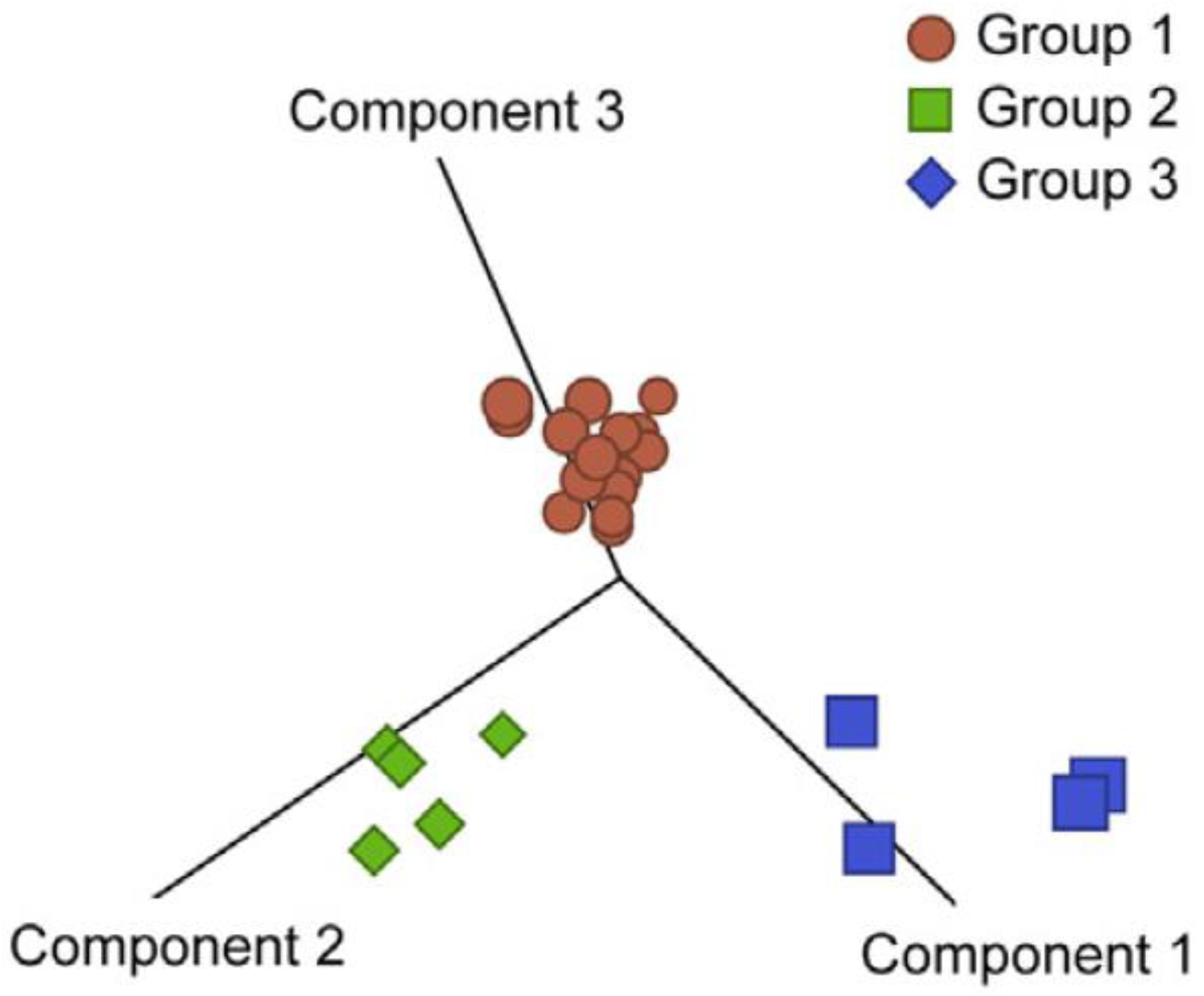
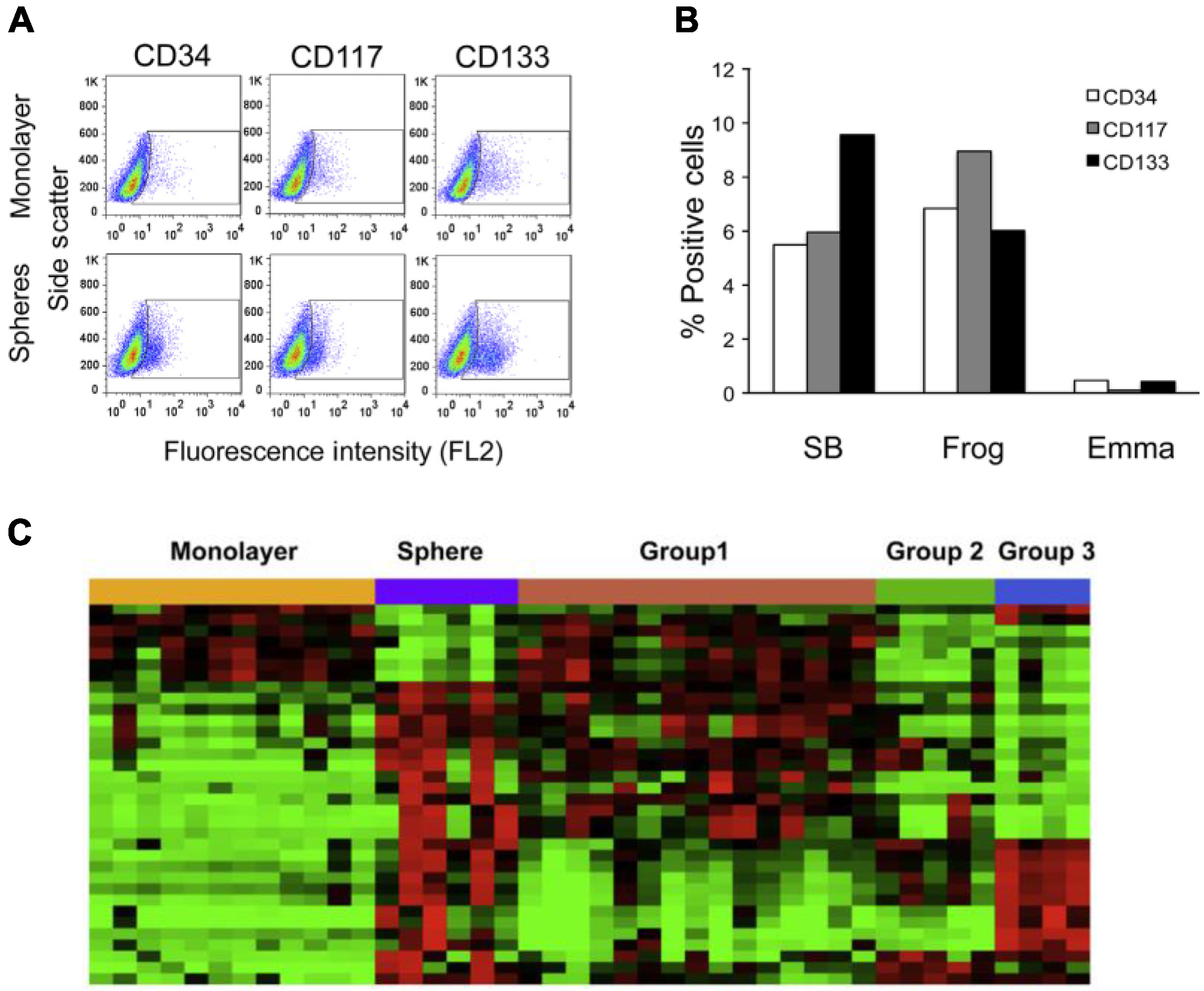
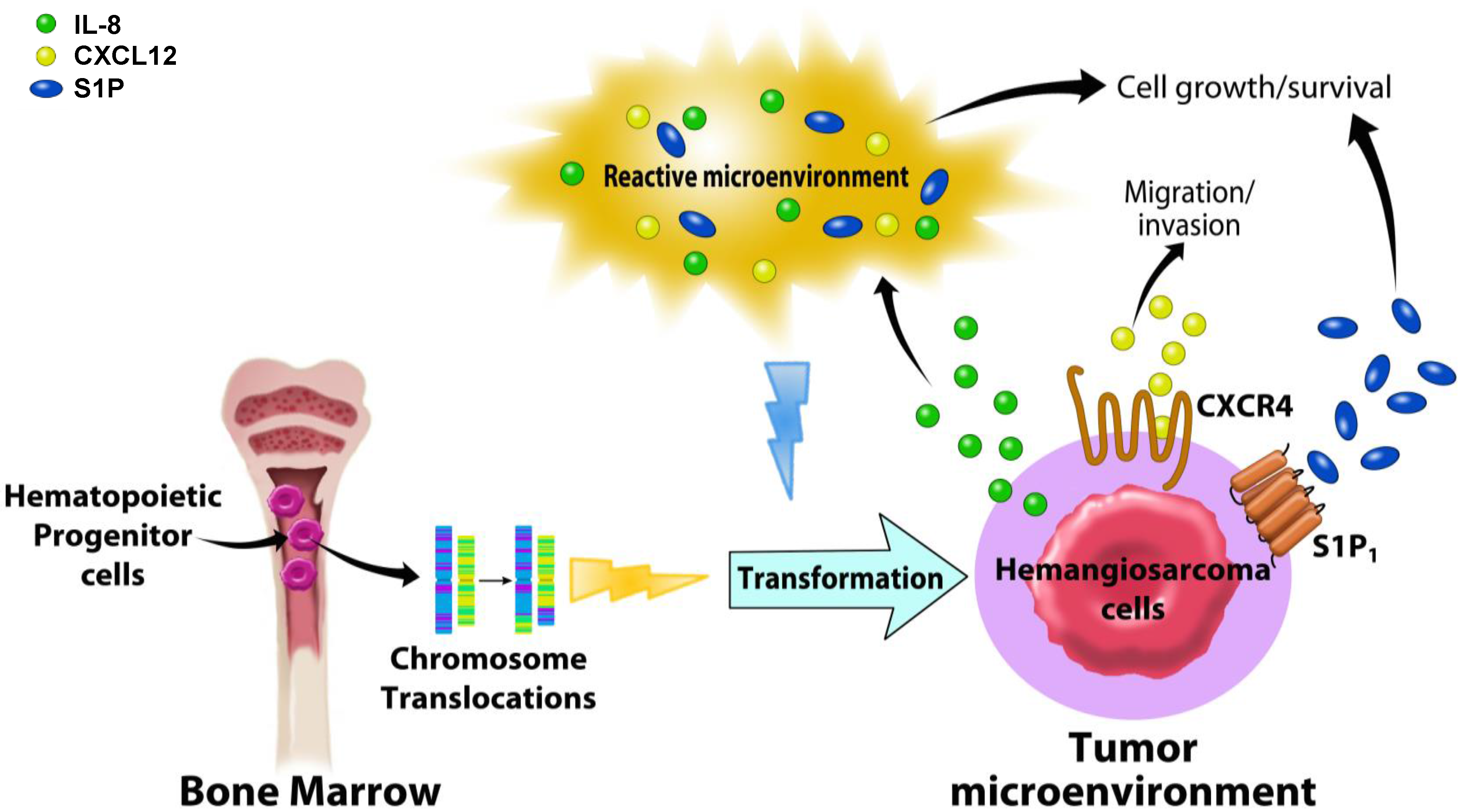
5. HSA and the Tumor Microenvironment
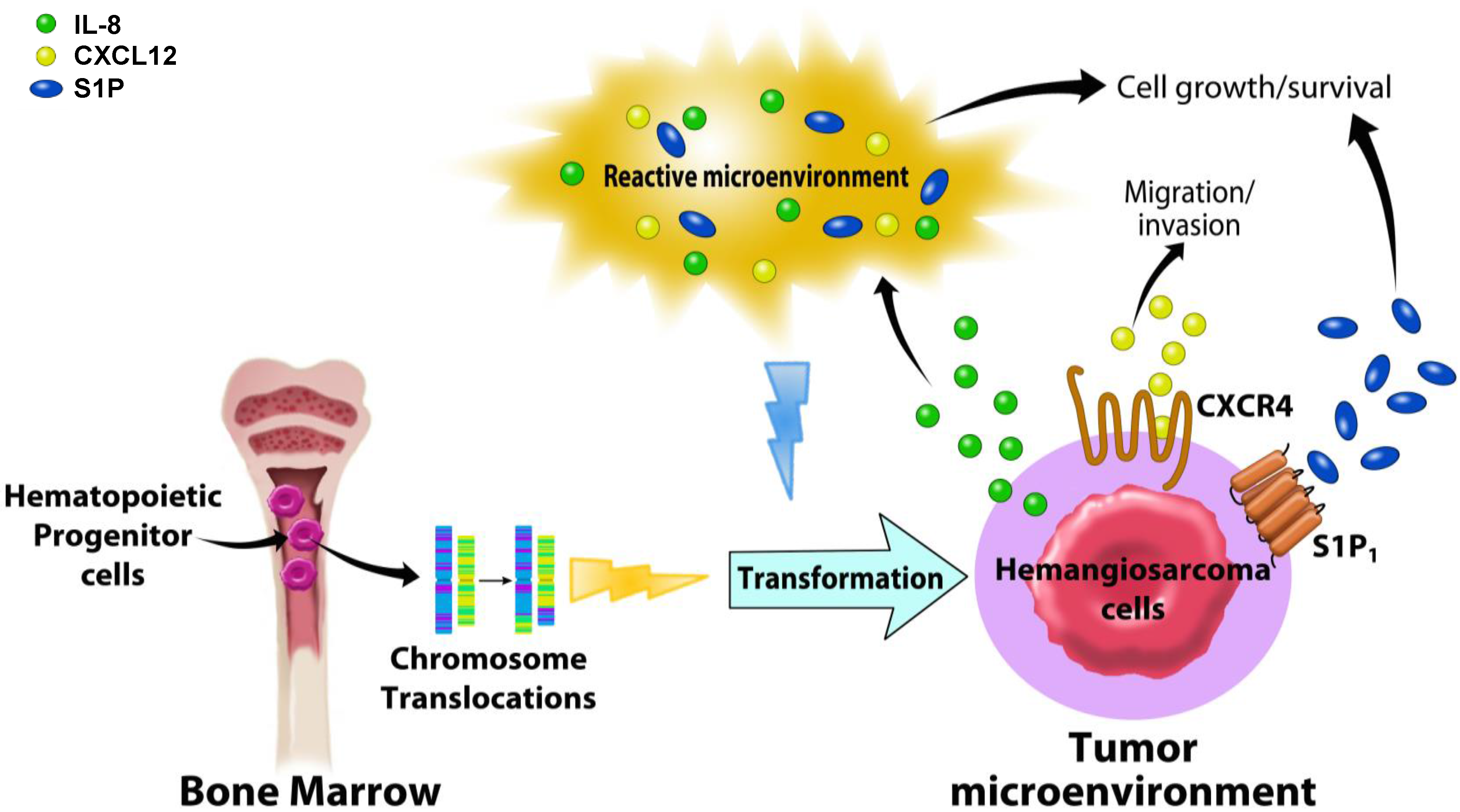
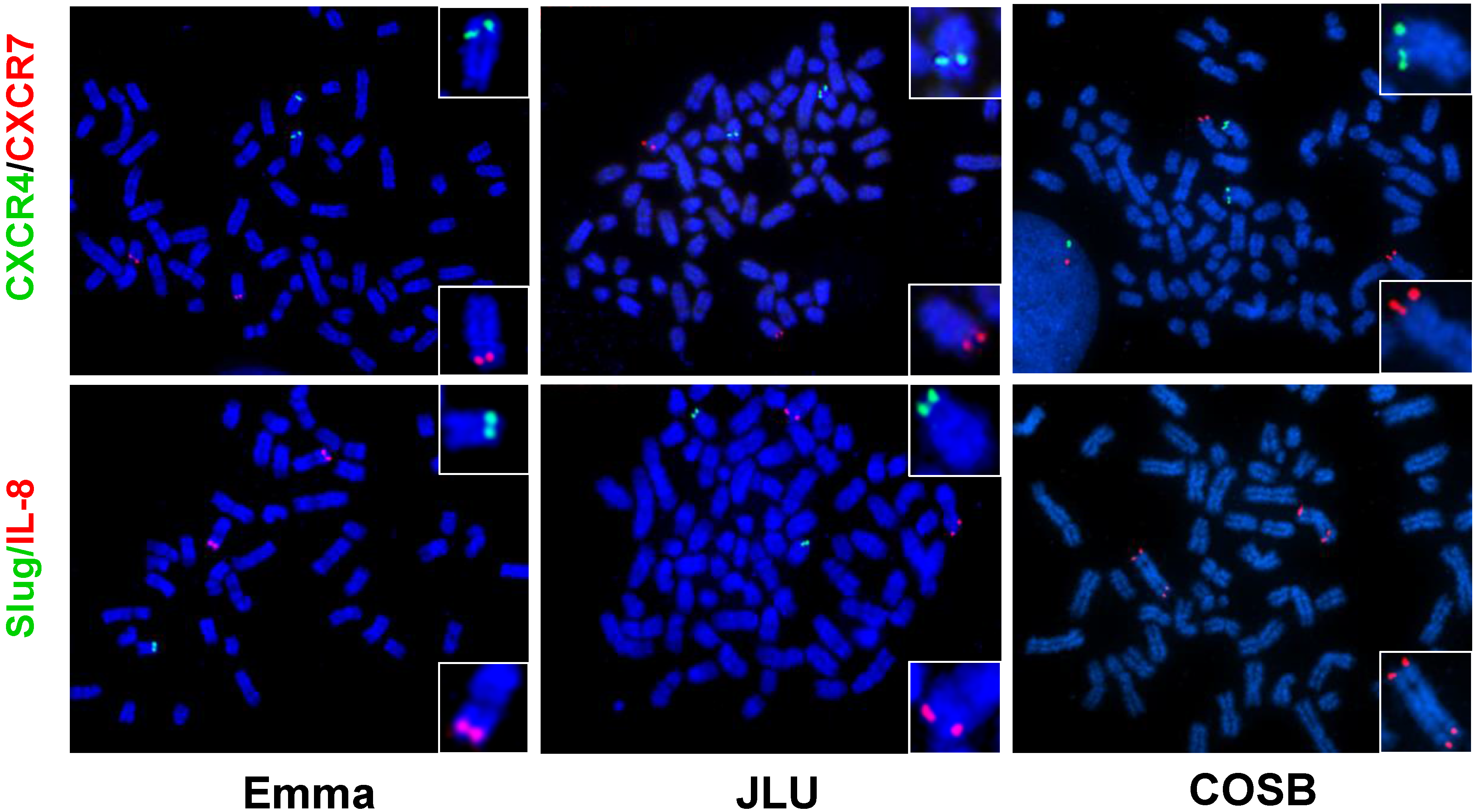
6. Future Directions
6.1. Establishment of Xenograft Models
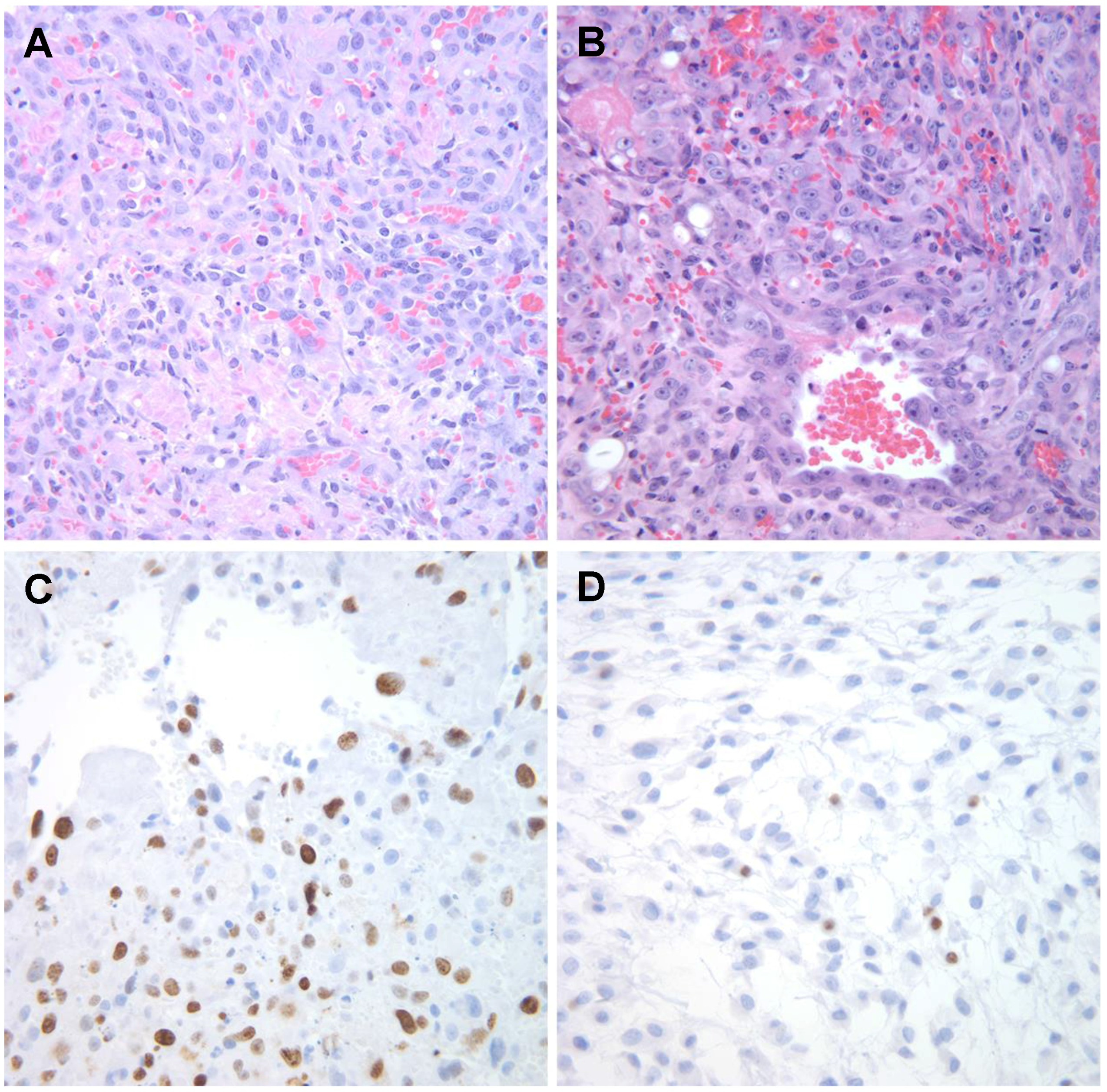
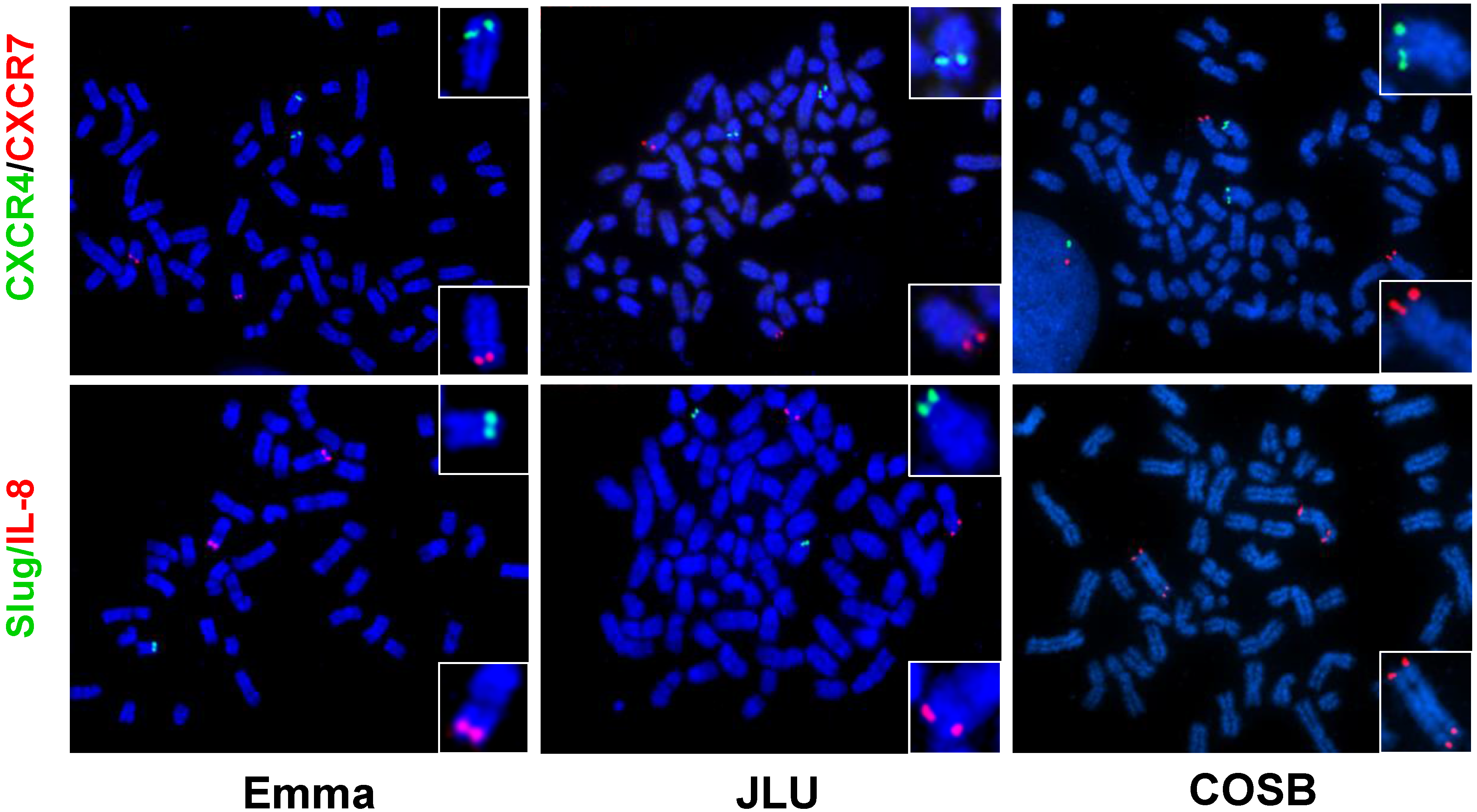
6.2. Chromosomal Translocations
7. Conclusions
Acknowledgments
Conflicts of Interest
References
- Bergman, P.J. Anticancer vaccines. Vet. Clin. North Am. Small Anim. Pract. 2007, 37, 1111–1119. [Google Scholar] [PubMed]
- London, C.A.; Malpas, P.B.; Wood-Follis, S.L.; Boucher, J.F.; Rusk, A.W.; Rosenberg, M.P.; Henry, C.J.; Mitchener, K.L.; Klein, M.K.; Hintermeister, J.G.; et al. Multi-center, placebo-controlled, double-blind, randomized study of oral toceranib phosphate (Su11654), a receptor tyrosine kinase inhibitor, for the treatment of dogs with recurrent (either local or distant) mast cell tumor following surgical excision. Clin. Cancer Res. 2009, 15, 3856–3865. [Google Scholar] [CrossRef] [PubMed]
- Spirtas, R.; Beebe, G.; Baxter, P.; Dacey, E.; Faber, M.; Falk, H.; van Kaick, G.; Stafford, J. Angiosarcoma as a model for comparative carcinogenesis (Letter to the Editor). Lancet 1983. [Google Scholar] [CrossRef]
- Falk, H.; Herbert, J.; Crowley, S.; Ishak, K.G.; Thomas, L.B.; Popper, H.; Caldwell, G.G. Epidemiology of hepatic angiosarcoma in the United States: 1964–1974. Environ. Health Perspect. 1981, 41, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, L.L. Malignant hemangioendothelioma of the canine heart. J. Am. Vet. Med. Assoc. 1955, 126, 296. [Google Scholar] [PubMed]
- Quigley, P.J.; De Saram, W.; Dawson, I.M.; Pryse-Davies, J. Two cases of haemangiosarcoma of the radius in the dog. Vet. Rec. 1965, 77, 1207–1209. [Google Scholar] [CrossRef] [PubMed]
- Geib, L.W. Primary angiomatous tumors of the heart and great vessels. A report of two cases in the dog. Cornell Vet. 1967, 57, 292–296. [Google Scholar] [PubMed]
- Slaughter, L.J.; Herman, L. Hemangiosarcoma in a dog. J. Am. Vet. Med. Assoc. 1969, 154, 17–21. [Google Scholar] [PubMed]
- Appleby, E.C.; Hayward, A.H.; Douce, G. German shepherds and splenic tumors. Vet. Rec. 1978. [Google Scholar] [CrossRef]
- Brown, N.O.; Patnaik, A.K.; MacEwen, E.G. Canine hemangiosarcoma: Retrospective analysis of 104 cases. J. Am. Vet. Med. Assoc. 1985, 186, 56–58. [Google Scholar] [PubMed]
- Prymak, C.; McKee, L.J.; Goldschmidt, M.H.; Glickman, L.T. Epidemiologic, clinical, pathologic, and prognostic characteristics of splenic hemangiosarcoma and splenic hematoma in dogs: 217 cases (1985). J. Am. Vet. Med. Assoc. 1988, 193, 706–712. [Google Scholar] [PubMed]
- Spangler, W.L.; Culbertson, M.R. Prevalence, type, and importance of splenic diseases in dogs: 1480 cases (1985–1989). J. Am. Vet. Med. Assoc. 1992, 200, 829–834. [Google Scholar] [PubMed]
- Priester, W.A.; McKay, F.W. The occurrence of tumors in domestic animals. Natl. Cancer Inst. Monogr. 1980, 54, 1–210. [Google Scholar] [PubMed]
- Ritt, M.G. Epidemiology of hematopoietic neoplasia. In Schalm’s Veterinary Hematology, 6th ed.; Weiss, D.J., Wardrop, K.J., Eds.; Blackwell Publishing: Ames, IA, USA, 2009. [Google Scholar]
- Parker, H.G.; Shearin, A.L.; Ostrander, E.A. Man’s best friend becomes biology’s best in show: Genome analyses in the domestic dog. Annu. Rev. Genet. 2010, 44, 309–336. [Google Scholar] [CrossRef] [PubMed]
- Koch, M.; Nielsen, G.P.; Yoon, S.S. Malignant tumors of blood vessels: Angiosarcomas, hemangioendotheliomas, and hemangioperictyomas. J. Surg. Oncol. 2008, 97, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.M.; Storer, R.D.; Criswell, K.A.; Doerrer, N.G.; Dellarco, V.L.; Pegg, D.G.; Wojcinski, Z.W.; Malarkey, D.E.; Jacobs, A.C.; Klaunig, J.E.; et al. Hemangiosarcoma in rodents: Mode-of-action evaluation and human relevance. Toxicol. Sci. 2009, 111, 4–18. [Google Scholar] [CrossRef] [PubMed]
- Fosmire, S.P.; Dickerson, E.B.; Scott, A.M.; Bianco, S.R.; Pettengill, M.J.; Meylemans, H.; Padilla, M.; Frazer-Abel, A.A.; Akhtar, N.; Getzy, D.M.; et al. Canine malignant hemangiosarcoma as a model of primitive angiogenic endothelium. Lab. Invest. 2004, 84, 562–572. [Google Scholar] [CrossRef] [PubMed]
- Schultheiss, P.C. A retrospective study of visceral and nonvisceral hemangiosarcoma and hemangiomas in domestic animals. J. Vet. Diagn. Invest. 2004, 16, 522–526. [Google Scholar] [CrossRef] [PubMed]
- Gorden, B.H.; Kim, J.H.; Sarver, A.L.; Frantz, A.M.; Breen, M.; Lindblad-Toh, K.; O’Brien, T.D.; Sharkey, L.C.; Modiano, J.F.; Dickerson, E.B. Identification of three molecular and functional subtypes in canine hemangiosarcoma through gene expression profiling and progenitor cell characterization. Am. J. Pathol. 2014, 184, 985–995. [Google Scholar] [CrossRef] [PubMed]
- Andersen, N.J.; Nickoloff, B.J.; Dykema, K.J.; Boguslawski, E.A.; Krivochenitser, R.I.; Froman, R.E.; Dawes, M.J.; Baker, L.H.; Thomas, D.G.; Kamstock, D.A.; et al. Pharmacologic inhibition of mek signaling prevents growth of canine hemangiosarcoma. Mol. Cancer Ther. 2013, 12, 1701–1714. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Frantz, A.M.; Anderson, K.L.; Graef, A.J.; Scott, M.C.; Robinson, S.; Sharkey, L.C.; O’Brien, T.D.; Dickerson, E.B.; Modiano, J.F. Interleukin-8 promotes canine hemangiosarcoma growth by regulating the tumor microenvironment. Exp. Cell Res. 2014, 323, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Ruoslahti, E. Specialization of tumour vasculature. Nat. Rev. Cancer 2002, 2, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Italiano, A.; Thomas, R.; Breen, M.; Zhang, L.; Crago, A.M.; Singer, S.; Khanin, R.; Maki, R.G.; Mihailovic, A.; Hafner, M.; et al. The miR-17–92 cluster and its target THBS1 are differentially expressed in angiosarcomas dependent on MYC amplification. Genes Chromosom. Cancer 2012, 51, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Antonescu, C. Malignant vascular tumors—An update. Mod. Pathol. 2014, 27, 30–38. [Google Scholar]
- Dobson, J.M. Breed-predispositions to cancer in pedigree dogs. ISRN Vet. Sci. 2013. [Google Scholar] [CrossRef] [PubMed]
- Tonomura, N.; Elvers, I.; Thomas, R.; Megquier, K.; Turner-Maier, J.; Howald, C.; Sarver, A.L.; Swofford, R.; Frantz, A.M.; Ito, D.; et al. Genome-wide association study identifies shared risk loci common to two malignancies in golden retrievers. PLoS Genet. 2015. [Google Scholar] [CrossRef] [PubMed]
- Tomasetti, C.; Vogelstein, B. Cancer etiology. Variation in cancer risk among tissues can be explained by the number of stem cell divisions. Science 2015, 347, 78–81. [Google Scholar] [CrossRef] [PubMed]
- Modiano, J.F. The genetic basis of cancer. In Withrow and MacEwen’s Small Animal Clinical Oncology, 5th ed.; Withrow, S.J., MacEwen, V.D., Page, R.L., Eds.; Elsevier/Saunders: St. Louis, MO, USA, 2013; pp. 1–15. [Google Scholar]
- Benjamin, S.A.; Hahn, F.F.; Chieffelle, T.L.; Boecker, B.B.; Hobbs, C.H. Occurrence of hemangiosarcomas in beagles with internally deposited radionuclides. Cancer Res. 1975, 35, 1745–1755. [Google Scholar] [PubMed]
- Benjamin, S.A.; Saunders, W.J.; Angleton, G.M.; Lee, A.C. Radiation carcinogenesis in dogs irradiated during prenatal and postnatal development. J. Radiat. Res. 1991, 32, 86–103. [Google Scholar] [CrossRef] [PubMed]
- Hahn, F.F.; Benjamin, S.A.; Boecker, B.B.; Chiffelle, T.L.; Hobbs, C.H.; Jones, R.K.; McClellan, R.O.; Pickrell, J.A.; Redman, H.C. Primary pulmonary neoplasms in beagle dogs exposed to aerosols of 144 CE in fused-clay particles. J. Natl. Cancer Inst. 1973, 50, 625–698. [Google Scholar] [PubMed]
- Hahn, F.F.; Benjamin, S.A.; Boecker, B.B.; Hobbs, C.H.; Jones, R.K.; McClellan, R.O.; Snipes, M.B. Comparative pulmonary carcinogenicity of inhaled beta-emitting radionuclides in beagle dogs. Inhaled Part 1975, 4 Pt 2, 625–636. [Google Scholar] [PubMed]
- Margarito, J.M.; Ginel, P.J.; Molleda, J.M.; Moreno, P.; Novales, M.; Lopez, R. Haemangiosarcoma associated with leishmaniasis in three dogs. Vet. Rec. 1994, 134, 66–67. [Google Scholar] [CrossRef] [PubMed]
- Varanat, M.; Maggi, R.G.; Linder, K.E.; Breitschwerdt, E.B. Molecular prevalence of Bartonella, Babesia, and hemotropic Mycoplasma sp. In dogs with splenic disease. J. Vet. Intern. Med. 2011, 25, 1284–1291. [Google Scholar] [CrossRef] [PubMed]
- Tamburini, B.A.; Trapp, S.; Phang, T.L.; Schappa, J.T.; Hunter, L.E.; Modiano, J.F. Gene expression profiles of sporadic canine hemangiosarcoma are uniquely associated with breed. PLoS ONE 2009. [Google Scholar] [CrossRef] [PubMed]
- Khammanivong, A.; Gorden, B.H.; Frantz, A.M.; Graef, A.J.; Dickerson, E.B. Identification of drug-resistant subpopulations in canine hemangiosarcoma. Vet. Comp. Oncol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Halsey, C.H.; Worley, D.R.; Curran, K.; Charles, J.B.; Ehrhart, E.J. The use of novel lymphatic endothelial cell-specific immunohistochemical markers to differentiate cutaneous angiosarcomas in dogs. Vet. Comp. Oncol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Lamerato-Kozicki, A.R.; Helm, K.M.; Jubala, C.M.; Cutter, G.R.; Modiano, J.F. Canine hemangiosarcoma originates from hematopoietic precursors with potential for endothelial differentiation. Exp. Hematol. 2006, 34, 870–878. [Google Scholar] [CrossRef] [PubMed]
- Tamburini, B.A.; Phang, T.L.; Fosmire, S.P.; Scott, M.C.; Trapp, S.C.; Duckett, M.M.; Robinson, S.R.; Slansky, J.E.; Sharkey, L.C.; Cutter, G.R.; et al. Gene expression profiling identifies inflammation and angiogenesis as distinguishing features of canine hemangiosarcoma. BMC Cancer 2010. [Google Scholar] [CrossRef] [PubMed]
- Dickerson, E.B.; Thomas, R.; Fosmire, S.P.; Lamerato-Kozicki, A.R.; Bianco, S.R.; Wojcieszyn, J.W.; Breen, M.; Helfand, S.C.; Modiano, J.F. Mutations of phosphatase and tensin homolog deleted from chromosome 10 in canine hemangiosarcoma. Vet. Pathol. 2005, 42, 618–632. [Google Scholar] [CrossRef] [PubMed]
- Laifenfeld, D.; Gilchrist, A.; Drubin, D.; Jorge, M.; Eddy, S.F.; Frushour, B.P.; Ladd, B.; Obert, L.A.; Gosink, M.M.; Cook, J.C.; et al. The role of hypoxia in 2-butoxyethanol-induced hemangiosarcoma. Toxicol. Sci. 2010, 113, 254–266. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Kakiuchi-Kiyota, S.; Arnold, L.L.; Johansson, S.L.; Wert, D.; Cohen, S.M. Pathogenesis of human hemangiosarcomas and hemangiomas. Hum. Pathol. 2013, 44, 2302–2311. [Google Scholar] [CrossRef] [PubMed]
- Gorden, B.H.; Saha, J.; Khammanivong, A.; Schwartz, G.K.; Dickerson, E.B. Lysosomal drug sequestration as a mechanism of drug resistance in vascular sarcoma cells marked by high CSF-1R expression. Vasc. Cell 2014. [Google Scholar] [CrossRef] [PubMed]
- Antonescu, C.R.; Yoshida, A.; Guo, T.; Chang, N.E.; Zhang, L.; Agaram, N.P.; Qin, L.X.; Brennan, M.F.; Singer, S.; Maki, R.G. Kdr activating mutations in human angiosarcomas are sensitive to specific kinase inhibitors. Cancer Res. 2009, 69, 7175–7179. [Google Scholar] [CrossRef] [PubMed]
- Italiano, A.; Chen, C.L.; Thomas, R.; Breen, M.; Bonnet, F.; Sevenet, N.; Longy, M.; Maki, R.G.; Coindre, J.M.; Antonescu, C.R. Alterations of the p53 and PIK3CA/AKT/mTOR pathways in angiosarcomas: A pattern distinct from other sarcomas with complex genomics. Cancer 2012, 118, 5878–5887. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Kantrow, S.; Sai, J.; Hawkins, O.E.; Boothby, M.; Ayers, G.D.; Young, E.D.; Demicco, E.G.; Lazar, A.J.; Lev, D.; et al. INK4A/ARF [corrected] inactivation with activation of the NF-κB/IL-6 pathway is sufficient to drive the development and growth of angiosarcoma. Cancer Res. 2012, 72, 4682–4695. [Google Scholar] [CrossRef] [PubMed]
- Weihrauch, M.; Bader, M.; Lehnert, G.; Koch, B.; Wittekind, C.; Wrbitzky, R.; Tannapfel, A. Mutation analysis of k-Ras-2 in liver angiosarcoma and adjacent nonneoplastic liver tissue from patients occupationally exposed to vinyl chloride. Environ. Mol. Mutagen. 2002, 40, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Thamm, D.H.; Dickerson, E.B.; Akhtar, N.; Lewis, R.; Auerbach, R.; Helfand, S.C.; MacEwen, E.G. Biological and molecular characterization of a canine hemangiosarcoma-derived cell line. Res. Vet. Sci. 2006, 81, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Lyles, S.E.; Milner, R.J.; Kow, K.; Salute, M.E. In vitro effects of the tyrosine kinase inhibitor, masitinib mesylate, on canine hemangiosarcoma cell lines. Vet. Comp. Oncol. 2012, 10, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Dickerson, E.B.; Marley, K.; Edris, W.; Tyner, J.W.; Schalk, V.; Macdonald, V.; Loriaux, M.; Druker, B.J.; Helfand, S.C. Imatinib and dasatinib inhibit hemangiosarcoma and implicate PDGFR-beta and SRC in tumor growth. Transl. Oncol. 2013, 6, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Marley, K.; Maier, C.S.; Helfand, S.C. Phosphotyrosine enrichment identifies focal adhesion kinase and other tyrosine kinases for targeting in canine hemangiosarcoma. Vet. Comp. Oncol. 2012, 10, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Murai, A.; Abou Asa, S.; Kodama, A.; Sakai, H.; Hirata, A.; Yanai, T. Immunohistochemical analysis of the AKT/mTOR/4e-BP1 signalling pathway in canine haemangiomas and haemangiosarcomas. J. Comp. Pathol. 2012, 147, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Murai, A.; Asa, S.A.; Kodama, A.; Hirata, A.; Yanai, T.; Sakai, H. Constitutive phosphorylation of the mTORc2/AKT/4e-BP1 pathway in newly derived canine hemangiosarcoma cell lines. BMC Vet. Res. 2012, 8, 128. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.; Borst, L.; Rotroff, D.; Motsinger-Reif, A.; Lindblad-Toh, K.; Modiano, J.F.; Breen, M. Genomic profiling reveals extensive heterogeneity in somatic DNA copy number aberrations of canine hemangiosarcoma. Chromosome Res. 2014, 22, 305–319. [Google Scholar] [CrossRef] [PubMed]
- Verbeke, S.L.; Bertoni, F.; Bacchini, P.; Oosting, J.; Sciot, R.; Krenacs, T.; Bovee, J.V. Active tgf-beta signaling and decreased expression of pten separates angiosarcoma of bone from its soft tissue counterpart. Mod. Pathol. 2013, 26, 1211–1221. [Google Scholar] [CrossRef] [PubMed]
- Tate, G.; Suzuki, T.; Mitsuya, T. Mutation of the pten gene in a human hepatic angiosarcoma. Cancer Genet. Cytogenet. 2007, 178, 160–162. [Google Scholar] [CrossRef] [PubMed]
- Choorapoikayil, S.; Kuiper, R.V.; de Bruin, A.; den Hertog, J. Haploinsufficiency of the genes encoding the tumor suppressor pten predisposes zebrafish to hemangiosarcoma. Dis. Model. Mech. 2012, 5, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Frantz, A.M. Comparative and Molecular Approaches to Improve Identification, Classification, and Therapeutic Approaches to Cancer; University of Minnesota: Minneapolis, MN, USA, 2012. [Google Scholar]
- Schappa, J.T.; Frantz, A.M.; Gorden, B.H.; Dickerson, E.B.; Vallera, D.A.; Modiano, J.F. Hemangiosarcoma and its cancer stem cell subpopulation are effectively killed by a toxin targeted through epidermal growth factor and urokinase receptors. Int. J. Cancer 2013, 133, 1936–1944. [Google Scholar] [CrossRef] [PubMed]
- Im, K.S.; Graef, A.J.; Breen, M.; Lindblad-Toh, K.; Modiano, J.F.; Kim, J.H. Interactions between CXCR4 and CXCL12 promote cell migration and invasion of canine hemangiosarcoma. Vet. Comp. Oncol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.M.; Graef, A.J.; LeVine, D.N.; Cohen, I.R.; Modiano, J.F.; Kim, J.H. Association of sphingosine-1-phosphate (S1P)/S1P receptor-1 pathway with cell proliferation and survival in canine hemangiosarcoma. J. Vet. Intern. Med. 2015, 29, 1088–1097. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-related inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Waugh, D.J.; Wilson, C. The interleukin-8 pathway in cancer. Clin. Cancer Res. 2008, 14, 6735–6741. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Aric, M.; Frantz, A.M.; Graef, A.J.; Hwang, T.; Scott, M.C.; Sarver, A.L.; Sharkey, L.C.; O’Brien, T.D.; Dickerson, E.B.; et al. Relationship between il-8 and Slug in Proliferation and Survival of Canine Hemangiosarcoma Cells; Abstracts Presented at the Second World Veterinary Cancer Congress, Paris, France, 1–3 March 2012.
- Askari, A.T.; Unzek, S.; Popovic, Z.B.; Goldman, C.K.; Forudi, F.; Kiedrowski, M.; Rovner, A.; Ellis, S.G.; Thomas, J.D.; DiCorleto, P.E.; et al. Effect of stromal-cell-derived factor 1 on stem-cell homing and tissue regeneration in ischaemic cardiomyopathy. Lancet 2003, 362, 697–703. [Google Scholar] [CrossRef]
- Muller, A.; Homey, B.; Soto, H.; Ge, N.; Catron, D.; Buchanan, M.E.; McClanahan, T.; Murphy, E.; Yuan, W.; Wagner, S.N.; et al. Involvement of chemokine receptors in breast cancer metastasis. Nature 2001, 410, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Balkwill, F. The significance of cancer cell expression of the chemokine receptor CXCR4. Semin. Cancer Biol. 2004, 14, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Balkwill, F. Cancer and the chemokine network. Nat. Rev. Cancer 2004, 4, 540–550. [Google Scholar] [CrossRef] [PubMed]
- Pyne, S.; Pyne, N.J. Sphingosine 1-phosphate signalling in mammalian cells. Biochem. J. 2000, 349, 385–402. [Google Scholar] [CrossRef] [PubMed]
- Spiegel, S.; Milstien, S. Sphingosine-1-phosphate: An enigmatic signalling lipid. Nat. Rev. Mol. Cell Biol. 2003, 4, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Kunkel, G.T.; Maceyka, M.; Milstien, S.; Spiegel, S. Targeting the sphingosine-1-phosphate axis in cancer, inflammation and beyond. Nat. Rev. Drug Discov. 2013, 12, 688–702. [Google Scholar] [CrossRef] [PubMed]
- Pyne, N.J.; Pyne, S. Sphingosine 1-phosphate and cancer. Nat. Rev. Cancer 2010, 10, 489–503. [Google Scholar] [CrossRef] [PubMed]
- Morton, C.L.; Houghton, P.J. Establishment of human tumor xenografts in immunodeficient mice. Nat. Protoc. 2007, 2, 247–250. [Google Scholar] [CrossRef] [PubMed]
- Camacho, L.; Pena, L.; Gonzalez Gil, A.; Caceres, S.; Diez, L.; Illera, J.C. Establishment and characterization of a canine xenograft model of inflammatory mammary carcinoma. Res. Vet. Sci. 2013, 95, 1068–1075. [Google Scholar] [CrossRef] [PubMed]
- Sonzogni-Desautels, K.; Knapp, D.W.; Sartin, E.; Dore, M. Effect of cyclooxygenase inhibitors in a xenograft model of canine mammary tumours. Vet. Comp. Oncol. 2011, 9, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Ito, D.; Endicott, M.M.; Jubala, C.M.; Helm, K.M.; Burnett, R.C.; Husbands, B.D.; Borgatti, A.; Henson, M.S.; Burgess, K.E.; Bell, J.S.; et al. A tumor-related lymphoid progenitor population supports hierarchical tumor organization in canine B-cell lymphoma. J. Vet. Intern. Med. 2011, 25, 890–896. [Google Scholar] [CrossRef] [PubMed]
- Laborda, E.; Puig-Saus, C.; Rodriguez-Garcia, A.; Moreno, R.; Cascallo, M.; Pastor, J.; Alemany, R. A PRB-responsive, rgd-modified, and hyaluronidase-armed canine oncolytic adenovirus for application in veterinary oncology. Mol. Ther. 2014, 22, 986–998. [Google Scholar] [CrossRef] [PubMed]
- Scharf, V.F.; Farese, J.P.; Siemann, D.W.; Abbott, J.R.; Kiupel, M.; Salute, M.E.; Milner, R.J. Effects of aurothiomalate treatment on canine osteosarcoma in a murine xenograft model. Anticancer Drugs 2014, 25, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, N.; Padilla, M.L.; Dickerson, E.B.; Steinberg, H.; Breen, M.; Auerbach, R.; Helfand, S.C. Interleukin-12 inhibits tumor growth in a novel angiogenesis canine hemangiosarcoma xenograft model. Neoplasia 2004, 6, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Kodama, A.; Sakai, H.; Matsuura, S.; Murakami, M.; Murai, A.; Mori, T.; Maruo, K.; Kimura, T.; Masegi, T.; Yanai, T. Establishment of canine hemangiosarcoma xenograft models expressing endothelial growth factors, their receptors, and angiogenesis-associated homeobox genes. BMC Cancer 2009. [Google Scholar] [CrossRef] [PubMed]
- Taylor, B.S.; Barretina, J.; Maki, R.G.; Antonescu, C.R.; Singer, S.; Ladanyi, M. Advances in sarcoma genomics and new therapeutic targets. Nat. Rev. Cancer 2011, 11, 541–557. [Google Scholar] [CrossRef] [PubMed]
- Neuville, A.; Ranchere-Vince, D.; Dei Tos, A.P.; Montesco, M.C.; Hostein, I.; Toffolatti, L.; Chibon, F.; Pissaloux, D.; Alberti, L.; Decouvelaere, A.V.; et al. Impact of molecular analysis on the final sarcoma diagnosis: A study on 763 cases collected during a european epidemiological study. Am. J. Surg. Pathol. 2013, 37, 1259–1268. [Google Scholar] [CrossRef] [PubMed]
- Helman, L.J.; Meltzer, P. Mechanisms of sarcoma development. Nat. Rev. Cancer 2003, 3, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Mertens, F.; Antonescu, C.R.; Hohenberger, P.; Ladanyi, M.; Modena, P.; D’Incalci, M.; Casali, P.G.; Aglietta, M.; Alvegard, T. Translocation-related sarcomas. Semin. Oncol. 2009, 36, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Maher, C.A.; Kumar-Sinha, C.; Cao, X.; Kalyana-Sundaram, S.; Han, B.; Jing, X.; Sam, L.; Barrette, T.; Palanisamy, N.; Chinnaiyan, A.M. Transcriptome sequencing to detect gene fusions in cancer. Nature 2009, 458, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Maher, C.A.; Palanisamy, N.; Brenner, J.C.; Cao, X.; Kalyana-Sundaram, S.; Luo, S.; Khrebtukova, I.; Barrette, T.R.; Grasso, C.; Yu, J.; et al. Chimeric transcript discovery by paired-end transcriptome sequencing. Proc. Natl. Acad. Sci. USA 2009, 106, 12353–12358. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.-H.; Graef, A.J.; Dickerson, E.B.; Modiano, J.F. Pathobiology of Hemangiosarcoma in Dogs: Research Advances and Future Perspectives. Vet. Sci. 2015, 2, 388-405. https://doi.org/10.3390/vetsci2040388
Kim J-H, Graef AJ, Dickerson EB, Modiano JF. Pathobiology of Hemangiosarcoma in Dogs: Research Advances and Future Perspectives. Veterinary Sciences. 2015; 2(4):388-405. https://doi.org/10.3390/vetsci2040388
Chicago/Turabian StyleKim, Jong-Hyuk, Ashley J. Graef, Erin B. Dickerson, and Jaime F. Modiano. 2015. "Pathobiology of Hemangiosarcoma in Dogs: Research Advances and Future Perspectives" Veterinary Sciences 2, no. 4: 388-405. https://doi.org/10.3390/vetsci2040388
APA StyleKim, J.-H., Graef, A. J., Dickerson, E. B., & Modiano, J. F. (2015). Pathobiology of Hemangiosarcoma in Dogs: Research Advances and Future Perspectives. Veterinary Sciences, 2(4), 388-405. https://doi.org/10.3390/vetsci2040388