

Advancing Zebrafish (Danio rerio) Welfare Using Immersion Analgesics





Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Database Search
2.2. Eligibility Criteria
2.3. Data Extraction and Organization
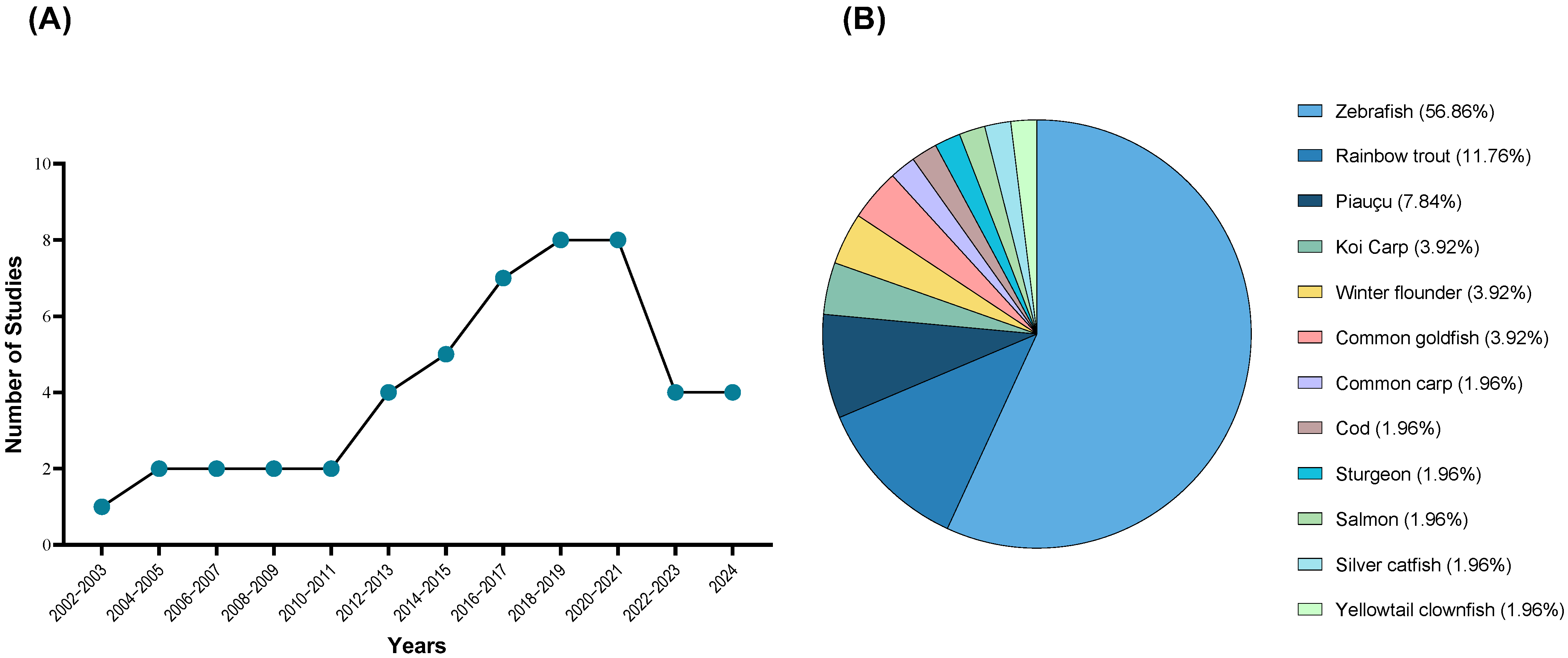
3. Results
3.1. Study Selection
3.2. Study Characteristics
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| TRP | Transient Receptor Potential |
| TRPV1 | Transient Receptor Potential Vanilloid Type-1 |
| TRPA1 | Transient Receptor Potential Ankyrin1 |
| NSAIDs | Nonsteroidal anti-inflammatory drugs |
| AITC | Allyl-isothiocyanate |
| dpf | Days Post Fertilization |
| Na+ | Sodium |
| K+ | Potassium |
| Ca2+ | Calcium |
| CO2 | Carbon dioxide |
| COX | Cyclooxygenase |
References
- Martins, T.; Valentim, A.; Pereira, N.; Antunes, L.M. Anaesthetics and analgesics used in adult fish for research: A review. Lab. Anim. 2019, 53, 325–341. [Google Scholar] [CrossRef] [PubMed]
- Sneddon, L.U. Evolution of nociception and pain: Evidence from fish models. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20190290. [Google Scholar] [CrossRef] [PubMed]
- Sneddon, L.U. The evidence for pain in fish: The use of morphine as an analgesic. Appl. Anim. Behav. Sci. 2003, 83, 153–162. [Google Scholar] [CrossRef]
- Sneddon, L.U. Trigeminal somatosensory innervation of the head of a teleost fish with particular reference to nociception. Brain Res. 2003, 972, 44–52. [Google Scholar] [CrossRef]
- Ohnesorge, N.; Heinl, C.; Lewejohann, L. Current Methods to Investigate Nociception and Pain in Zebrafish. Front. Neurosci. 2021, 15, 632634. [Google Scholar] [CrossRef]
- Sneddon, L.U. Anatomical and electrophysiological analysis of the trigeminal nerve in a teleost fish, Oncorhynchus mykiss. Neurosci. Lett. 2002, 319, 167–171. [Google Scholar] [CrossRef]
- Newby, N.C.; Gamperl, A.K.; Stevens, E.D. Cardiorespiratory effects and efficacy of morphine sulfate in winter flounder (Pseudopleuronectes americanus). Am. J. Vet. Res. 2007, 68, 592–597. [Google Scholar] [CrossRef]
- Newby, N.C.; Wilkie, M.P.; Stevens, E.D. Morphine uptake, disposition, and analgesic efficacy in the common goldfish (Carassius auratus). Can J. Zool. 2009, 87, 388–399. [Google Scholar] [CrossRef]
- Nordgreen, J.; Garner, J.P.; Janczak, A.M.; Ranheim, B.; Muir, W.M.; Horsberg, T.E. Thermonociception in fish: Effects of two different doses of morphine on thermal threshold and post-test behaviour in goldfish (Carassius auratus). Appl. Anim. Behav. Sci. 2009, 119, 101–107. [Google Scholar] [CrossRef]
- Correia, A.D.; Cunha, S.R.; Scholze, M.; Stevens, E.D. A Novel Behavioral Fish Model of Nociception for Testing Analgesics. Pharmaceuticals 2011, 4, 665–680. [Google Scholar] [CrossRef]
- Steenbergen, P.J.; Bardine, N. Antinociceptive effects of buprenorphine in zebrafish larvae: An alternative for rodent models to study pain and nociception? Appl. Anim. Behav. Sci. 2014, 152, 92–99. [Google Scholar] [CrossRef]
- Nordgreen, J.; Bjørge, M.H.; Janczak, A.M.; Poppe, T.; Koppang, E.O.; Ranheim, B.; Horsberg, T.E. The effect of morphine on changes in behaviour and physiology in intraperitoneally vaccinated Atlantic salmon (Salmo salar). Appl. Anim. Behav. Sci. 2013, 145, 129–137. [Google Scholar] [CrossRef]
- Rodrigues, P.; Barbosa, L.B.; Bianchini, A.E.; Ferrari, F.T.; Baldisserotto, B.; Heinzmann, B.M. Nociceptive-like behavior and analgesia in silver catfish (Rhamdia quelen). Physiol. Behav. 2019, 210, 112648. [Google Scholar] [CrossRef] [PubMed]
- Raja, S.N.; Carr, D.B.; Cohen, M.; Finnerup, N.B.; Flor, H.; Gibson, S.; Keefe, F.J.; Mogil, J.S.; Ringkamp, M.; Sluka, K.A.; et al. The revised International Association for the Study of Pain definition of pain: Concepts, challenges, and compromises. Pain 2020, 161, 1976–1982. [Google Scholar] [CrossRef]
- Sneddon, L.U. Ethics and welfare: Pain perception in fish. Bull. Eur. Ass. Fish Pathol. 2006, 26, 6–10. [Google Scholar]
- Dunlop, R.; Laming, P. Mechanoreceptive and nociceptive responses in the central nervous system of goldfish (Carassius auratus) and trout (Oncorhynchus mykiss). J. Pain 2005, 6, 561–568. [Google Scholar] [CrossRef]
- Sneddon, L.U.; Schroeder, P.; Roque, A.; Finger-Baier, K.; Fleming, A.; Tinman, S.; Collet, B. Pain management in zebrafish: Report from a FELASA Working Group. Lab. Anim. 2024, 58, 261–276. [Google Scholar] [CrossRef]
- Sneddon, L.U.; Braithwaite, V.A.; Gentle, M.J. Novel object test: Examining nociception and fear in the rainbow trout. J. Pain 2003, 4, 431–440. [Google Scholar] [CrossRef]
- Briggs, P.J. The zebrafish: A new model organism for integrative physiology. Am. J. Physiol. 2002, 282, R3–R9. [Google Scholar] [CrossRef]
- Meyers, J.R. Zebrafish: Development of a Vertebrate Model Organism. Curr. Protoc. Essent. Lab. Tech. 2018, 16, e19. [Google Scholar] [CrossRef]
- Weber, E.S. Fish analgesia: Pain, stress, fear aversion, or nociception? Vet. Clin. N. Am. Exot. Anim. Pract. 2011, 14, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Chatigny, F.; Creighton, C.M.; Stevens, D. Updated Review of Fish Analgesia. J. Am. Assoc. Lab. Anim. 2018, 57, 5–12. [Google Scholar]
- Harms, C. Surgery in Fish Research: Common Procedures and Postoperative Care. Lab. Anim. 2005, 34, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Palazzo, E.; Rossi, F.; Maione, S. Role of TRPV1 receptors in descending modulation of pain. Mol. Cell. Endocrinol. 2008, 286, S79–S83. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Chang, R.B.; Allgood, S.D.; Silver, W.L.; Liman, E.R. A TRPA1-dependent mechanism for the pungent sensation of weak acids. J. Gen. Physiol. 2011, 137, 493–505. [Google Scholar] [CrossRef]
- Curtright, A.; Rosser, M.; Goh, S.; Keown, B.; Wagner, E.; Sharifi, J.; Raible, D.W.; Dhaka, A. Modeling nociception in zebrafish: A way forward for unbiased analgesic discovery. PLoS ONE 2015, 10, e0116766. [Google Scholar] [CrossRef]
- Prober, D.A.; Zimmerman, S.; Myers, B.R.; McDermott, B.M., Jr.; Kim, S.H.; Caron, S.; Rihel, J.; Solnica-Krezel, L.; Julius, D.; Hudspeth, A.J.; et al. Zebrafish TRPA1 channels are required for chemosensation but not for thermosensation or mechanosensory hair cell function. J. Neurosci. 2008, 28, 10102–10110. [Google Scholar] [CrossRef]
- Deakin, A.G.; Buckley, J.; AlZu’bi, H.S.; Cossins, A.R.; Spencer, J.W.; Al’Nuaimy, W.; Young, I.S.; Thomson, J.S.; Sneddon, L.U. Automated monitoring of behaviour in zebrafish after invasive procedures. Sci. Rep. 2019, 9, 9042. [Google Scholar] [CrossRef]
- Lopez-Luna, J.; Al-Jubouri, Q.; Al-Nuaimy, W.; Sneddon, L.U. Impact of analgesic drugs on the behavioural responses of larval zebrafish to potentially noxious temperatures. Appl. Anim. Behav. Sci. 2017, 188, 97–105. [Google Scholar] [CrossRef]
- Steenbergen, P.J. Response of zebrafish larvae to mild electrical stimuli: A 96-well setup for behavioural screening. J. Neurosci. Methods 2018, 301, 52–61. [Google Scholar] [CrossRef]
- Reilly, S.C.; Quinn, J.P.; Cossins, A.R.; Sneddon, L.U. Behavioural analysis of a nociceptive event in fish: Comparisons between three species demonstrate specific responses. Appl. Anim. Behav. Sci. 2008, 114, 248–259. [Google Scholar] [CrossRef]
- Kalueff, A.V.; Gebhardt, M.; Stewart, A.M.; Cachat, J.M.; Brimmer, M.; Chawla, J.S.; Craddock, C.; Kyzar, E.J.; Roth, A.; Landsman, S.; et al. Towards a Comprehensive Catalog of Zebrafish Behavior 1.0 and Beyond. Zebrafish 2013, 10, 70–86. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, F.E.A.; de Sousa, C.; Santos, S.; Menezes, R.B.; Batista, F.L.A.; Abreu, A.O.; de Oliveira, M.V.; Moura, L.; Raposo, R.D.S.; Campos, A.R. Adult Zebrafish (Danio rerio): An Alternative Behavioral Model of Formalin-Induced Nociception. Zebrafish 2017, 14, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Baker, T.R.; Baker, B.B.; Johnson, S.M.; Sladky, K.K. Comparative analgesic efficacy of morphine sulfate and butorphanol tartrate in koi (Cyprinus carpio) undergoing unilateral gonadectomy. J. Am. Vet. Med. Assoc. 2013, 243, 882–890. [Google Scholar] [CrossRef]
- Mettam, J.J.; Oulton, L.J.; McCrohan, C.R.; Sneddon, L.U. The efficacy of three types of analgesic drugs in reducing pain in the rainbow trout, Oncorhynchus mykiss. Appl. Anim. Behav. Sci. 2011, 133, 265–274. [Google Scholar] [CrossRef]
- Lopez-Luna, J.; Canty, M.N.; Al-Jubouri, Q.; Al-Nuaimy, W.; Sneddon, L.U. Behavioural responses of fish larvae modulated by analgesic drugs after a stress exposure. Appl. Anim. Behav. Sci. 2017, 195, 115–120. [Google Scholar] [CrossRef]
- Correia, A.M.; Pedrazzani, A.S.; Mendonca, R.C.; Massucatto, A.; Ozorio, R.A.; Tsuzuki, M.Y. Basil, tea tree and clove essential oils as analgesics and anaesthetics in Amphiprion clarkii (Bennett, 1830). Braz. J. Biol. 2018, 78, 436–442. [Google Scholar] [CrossRef]
- Soares, I.C.R.; Santos, S.A.A.R.; Coelho, R.F.; Alves, Y.A.; Vieira-Neto, A.E.; Tavares, K.C.S.; Magalhaes, F.E.A.; Campos, A.R. Oleanolic acid promotes orofacial antinociception in adult zebrafish (Danio rerio) through TRPV1 receptors. Chem. Biol. Interact. 2019, 299, 37–43. [Google Scholar] [CrossRef]
- Pereira, W.F.; Everson da Silva, L.; do Amaral, W.; Andrade Rebelo, R.; Quefi, B.; Wlisses da Silva, A.; Silva Marinho, E.; Borges Leal, A.L.A.; Mesquita Cajazeiras, F.F.; Amancio Ferreira, M.K.; et al. Essential Oils from the Genus Piper Promote Antinociception by Modulating TRP Channels and Anti-Inflammatory Effects in Adult Zebrafish. Chem. Biodivers. 2024, 21, e202301807. [Google Scholar] [CrossRef]
- Rocha, C.A.; Felix, L.M.; Monteiro, S.M.; Venancio, C. Antinociceptive Analysis of Natural Monoterpenes Eugenol, Menthol, Carvacrol and Thymol in a Zebrafish Larval Model. Pharmaceuticals 2024, 17, 457. [Google Scholar] [CrossRef]
- Schroeder, P.G.; Sneddon, L.U. Exploring the efficacy of immersion analgesics in zebrafish using an integrative approach. Appl. Anim. Behav. Sci. 2017, 187, 93–102. [Google Scholar] [CrossRef]
- Zahl, I.H.; Samuelsen, O.; Kiessling, A. Anaesthesia of farmed fish: Implications for welfare. Fish Physiol. Biochem. 2012, 38, 201–218. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. J. Clin. Epidemiol. 2021, 134, 178–189. [Google Scholar] [CrossRef]
- Neiffer, D.L. Anesthesia and Analgesia. In Clinical Guide to Fish Medicine, 1st ed.; Hadfield, C.A., Clayton, L.A., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2021; pp. 198–212. [Google Scholar] [CrossRef]
- Esmail, M.Y.; Astrofsky, K.M.; Lawrence, C.; Serluca, F.C. The Biology and Management of the Zebrafish. In Laboratory Animal Medicine, 3rd ed.; Fox, J.G., Anderson, L.C., Otto, G.M., Pritchett-Corning, K.R., Whary, M.T., Eds.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 1015–1062. [Google Scholar] [CrossRef]
- Sneddon, L.U. Clinical Anesthesia and Analgesia in Fish. J. Exot. Pet Med. 2012, 21, 32–43. [Google Scholar] [CrossRef]
- Bramer, W.M.; Giustini, D.; de Jonge, G.B.; Holland, L.; Bekhuis, T. De-duplication of database search results for systematic reviews in EndNote. J. Med. Libr. Assoc. 2016, 104, 240–243. [Google Scholar] [CrossRef]
- McGrath, S.; Zhao, X.; Steele, R.; Thombs, B.D.; Benedetti, A. Estimating the sample mean and standard deviation from commonly reported quantiles in meta-analysis. Stat. Methods Med. Res. 2020, 29, 2520–2537. [Google Scholar] [CrossRef]
- Alestrom, P.; D’Angelo, L.; Midtlyng, P.J.; Schorderet, D.F.; Schulte-Merker, S.; Sohm, F.; Warner, S. Zebrafish: Housing and husbandry recommendations. Lab. Anim. 2020, 54, 213–224. [Google Scholar] [CrossRef]
- Zaig, S.; da Silveira Scarpellini, C.; Montandon, G. Respiratory depression and analgesia by opioid drugs in freely behaving larval zebrafish. eLife 2021, 10, e63407. [Google Scholar] [CrossRef]
- Lopez-Luna, J.; Al-Jubouri, Q.; Al-Nuaimy, W.; Sneddon, L.U. Impact of stress, fear and anxiety on the nociceptive responses of larval zebrafish. PLoS ONE 2017, 12, e0181010. [Google Scholar] [CrossRef]
- Lopez-Luna, J.; Al-Jubouri, Q.; Al-Nuaimy, W.; Sneddon, L.U. Reduction in activity by noxious chemical stimulation is ameliorated by immersion in analgesic drugs in zebrafish. J. Exp. Biol. 2017, 220 Pt 8, 1451–1458. [Google Scholar] [CrossRef]
- Sneddon, L.U. Pain Perception in Fish: Indicators and Endpoints. ILAR J. 2009, 50, 338–342. [Google Scholar] [CrossRef]
- Ko, M.J.; Ganzen, L.C.; Coskun, E.; Mukadam, A.A.; Leung, Y.F.; van Rijn, R.M. A critical evaluation of TRPA1-mediated locomotor behavior in zebrafish as a screening tool for novel anti-nociceptive drug discovery. Sci. Rep. 2019, 9, 2430. [Google Scholar] [CrossRef]
- Segner, H. Fish. Nociception and Pain. A Biological Perspective; Federal Office for Buildings and Logistics (FOBL): Bern, Switzerland, 2012; Volume 9. [Google Scholar]
- Guimaraes, A.G.; Quintans, J.S.; Quintans, L.J., Jr. Monoterpenes with analgesic activity—A systematic review. Phytother. Res. 2013, 27, 1–15. [Google Scholar] [CrossRef]
- Yang, X.; Wei, X.; Mu, Y.; Li, Q.; Liu, J. A review of the mechanism of the central analgesic effect of lidocaine. Medicine 2020, 99, e19898. [Google Scholar] [CrossRef]
- Stanley, T.H. The fentanyl story. J. Pain 2014, 15, 1215–1226. [Google Scholar] [CrossRef]
- Rosland, J.H.; Tjølsen, A.; Bjørn, M.; Hole, K. The formalin test in mice: Effect of formalin concentration. Pain 1990, 42, 235–242. [Google Scholar] [CrossRef]
- Sawynok, J.; Liu, X.J. The Formalin Test: Characteristics and Usefulness of the Model. Rev. Analg. 2003, 7, 145–163. [Google Scholar] [CrossRef]
- Hunskaar, S.; Hole, K. The formalin test in mice: Dissociation between inflammatory and non-inflammatory pain. Pain 1987, 30, 103–114. [Google Scholar] [CrossRef]
- Oguri, G.; Nakajima, T.; Kikuchi, H.; Obi, S.; Nakamura, F.; Komuro, I. Allyl isothiocyanate (AITC) activates nonselective cation currents in human cardiac fibroblasts: Possible involvement of TRPA1. Heliyon 2021, 7, e05816. [Google Scholar] [CrossRef]
- Roberts, A.C.; Alzagatiti, J.B.; Ly, D.T.; Chornak, J.M.; Ma, Y.; Razee, A.; Zavradyan, G.; Khan, U.; Lewis, J.; Natarajan, A.; et al. Induction of Short-Term Sensitization by an Aversive Chemical Stimulus in Zebrafish Larvae. eNeuro 2020, 7, ENEURO.0336-19.2020. [Google Scholar] [CrossRef]
- Bao, Y.; Gao, Y.; Yang, L.; Kong, X.; Yu, J.; Hou, W.; Hua, B. The mechanism of mu-opioid receptor (MOR)-TRPV1 crosstalk in TRPV1 activation involves morphine anti-nociception, tolerance and dependence. Channels 2015, 9, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Stein, C. New concepts in opioid analgesia. Expert Opin. Investig. Drugs 2018, 27, 765–775. [Google Scholar] [CrossRef] [PubMed]
- Webster, L.R.; Grabois, M. Current Regulations Related to Opioid Prescribing. PM&R 2015, 7, S236–S247. [Google Scholar] [CrossRef]
- Cashman, J.N. The Mechanisms of Action of NSAIDs in Analgesia. Drugs 1996, 52, 3–23. [Google Scholar] [CrossRef]
- Munir, M.A.; Enany, N.; Zhang, J.M. Nonopioid analgesics. Anesthesiol. Clin. 2007, 25, 761–774. [Google Scholar] [CrossRef]
- Armstrong, S.A.; Herr, M.J. Physiology, Nociception. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2024. Available online: https://www.ncbi.nlm.nih.gov/pubmed/31855389 (accessed on 10 November 2024).
- Flecknell, P.A. Analgesia in small mammals. Semin. Avian Exot. Pet Med. 1998, 7, 41–47. [Google Scholar] [CrossRef]
- Hammer, H.S. Water Quality for Zebrafish Culture. In The Zebrafish in Biomedical Research, 1st ed.; Cartner, S.C., Eisen, J.S., Farmer, S.C., Guillemin, K.J., Kent, M.L., Sanders, G.E., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 321–335. [Google Scholar] [CrossRef]
- Lee, C.J.; Paull, G.C.; Tyler, C.R. Improving zebrafish laboratory welfare and scientific research through understanding their natural history. Biol. Rev. Camb. Philos. Soc. 2022, 97, 1038–1056. [Google Scholar] [CrossRef]
- Lawrence, C. The husbandry of zebrafish (Danio rerio): A review. Aquaculture 2007, 269, 1–20. [Google Scholar] [CrossRef]
- Leal, J.F.; Neves, M.G.P.M.S.; Santos, E.B.H.; Esteves, V.I. Use of formalin in intensive aquaculture: Properties, application and effects on fish and water quality. Rev. Aquac. 2016, 10, 281–295. [Google Scholar] [CrossRef]
- Neiffer, D.L.; Stamper, M.A. Fish Sedation, Anesthesia, Analgesia, and Euthanasia: Considerations, Methods, and Types of Drugs. ILAR J. 2009, 50, 343–360. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Noxious Stimulus | Zebrafish Stage | Compound | Concentration (mg/L) | Normalized Swimming Velocity | Reference | |
|---|---|---|---|---|---|---|
| Chemical | Acetic acid (0.05%) | 5 dpf | Eugenol | 10 | 0.76 | [40] |
| Acetic acid (0.05%) | 5 dpf | Menthol | 2 | 1.11 | [40] | |
| Acetic acid (0.05%) | 5 dpf | Thymol | 2 | 1.04 | [40] | |
| Acetic acid (0.05%) | 5 dpf | Carvacrol | 2 | 1.14 | [40] | |
| Acetic acid (0.1%) | 5 dpf | Lidocaine | 5 | 0.91 | [51] | |
| Formalin (0.05%) | 12–14 dpf | Fentanyl | 1 | 2.31 | [50] | |
| Allyl isothiocyanate (AITC) (100 uM) | 12–14 dpf | Fentanyl | 2 | 1.28 | [50] | |
| Thermal | Hot water (40 °C) | 5 dpf | Morphine | 48 | 1.01 | [29] |
| Hot water (40 °C) | 5 dpf | Lidocaine | 5 | 0.97 | [29] | |
| Noxious Stimulus | Zebrafish Stage | Compound | Concentration (mg/L) | Normalized % Active Time | Reference | |
|---|---|---|---|---|---|---|
| Chemical | Acetic acid (0.1%) | 5 dpf | Lidocaine | 5 | 0.86 | [51] |
| Acetic acid (0.1%) | 5 dpf | Lidocaine | 5 | 1.02 | [52] | |
| Acetic acid (0.1%) | 5 dpf | Morphine | 48 | 0.86 | [52] | |
| Acetic acid (0.1%) | 5 dpf | Aspirin | 2.5 | 0.88 | [52] | |
| Soda water (10%) | 5 dpf | Lidocaine | 5 | 0.96 | [36] | |
| Soda water (10%) | 5 dpf | Morphine | 48 | 1.00 | [36] | |
| Thermal | Hot water (40 °C) | 5 dpf | Flunixin | 20 | 1.15 | [29] |
| Hot water (40 °C) | 5 dpf | Lidocaine | 5 | 1.22 | [29] | |
| Hot water (40 °C) | 5 dpf | Morphine | 48 | 1.10 | [29] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rocha, C.A.; Félix, L.M.; Monteiro, S.M.; Venâncio, C. Advancing Zebrafish (Danio rerio) Welfare Using Immersion Analgesics. Vet. Sci. 2025, 12, 571. https://doi.org/10.3390/vetsci12060571
Rocha CA, Félix LM, Monteiro SM, Venâncio C. Advancing Zebrafish (Danio rerio) Welfare Using Immersion Analgesics. Veterinary Sciences. 2025; 12(6):571. https://doi.org/10.3390/vetsci12060571
Chicago/Turabian StyleRocha, Cláudia A., Luís M. Félix, Sandra M. Monteiro, and Carlos Venâncio. 2025. "Advancing Zebrafish (Danio rerio) Welfare Using Immersion Analgesics" Veterinary Sciences 12, no. 6: 571. https://doi.org/10.3390/vetsci12060571
APA StyleRocha, C. A., Félix, L. M., Monteiro, S. M., & Venâncio, C. (2025). Advancing Zebrafish (Danio rerio) Welfare Using Immersion Analgesics. Veterinary Sciences, 12(6), 571. https://doi.org/10.3390/vetsci12060571