
Foot-and-Mouth Disease Virus-like Particles Produced in E. coli as Potential Antigens for a Novel Vaccine

 ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
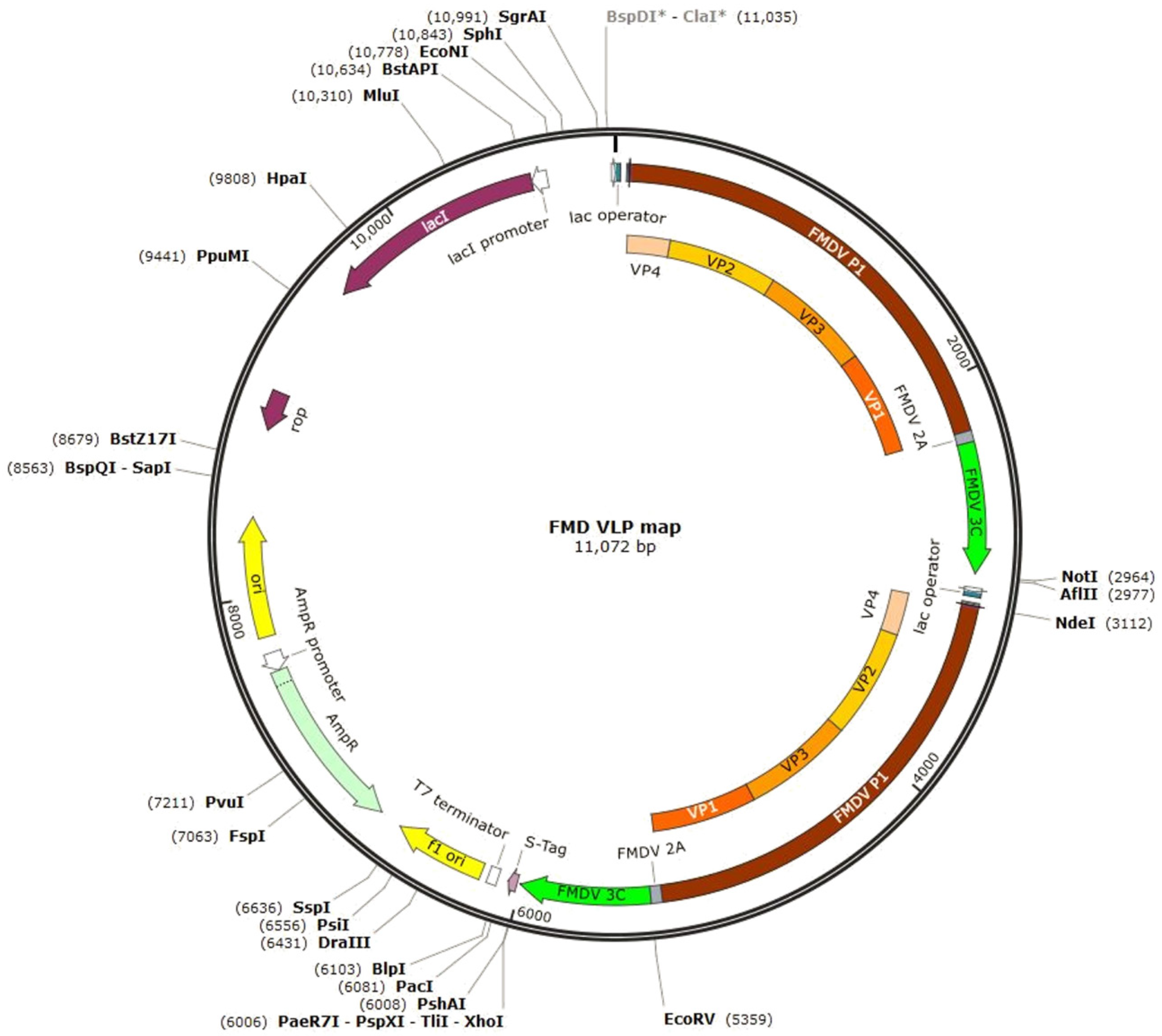
2.1. Plasmid Construction
2.2. Preparation of FMDV VLP Antigen
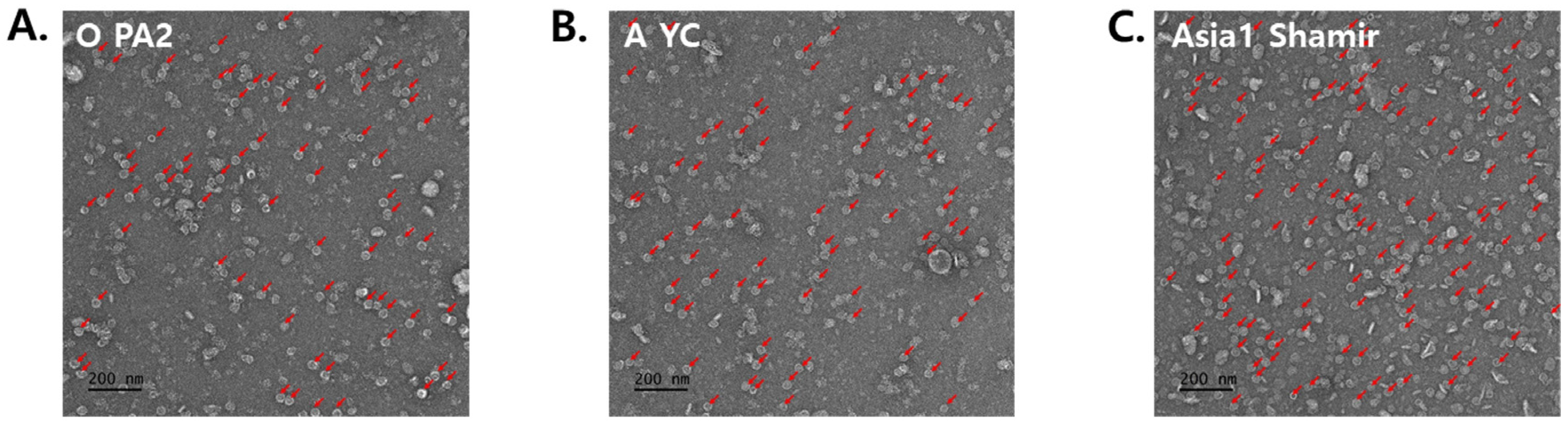
2.3. Characterization and Measurement of FMDV VLPs
2.4. Purification of VLPs Using Sucrose Gradient Centrifugation
2.5. Vaccine Preparation and Mouse Immunization
2.6. Detection of FMDV-VLP Specific Antibodies in Mouse Sera
2.7. Neutralizing Antibody Titer Assay in Mouse Sera
2.8. Challenge Study in Immunized Mice
2.9. Challenge Study in Immunized Swine
2.10. Control Groups
2.11. Statistical Analysis
3. Results
3.1. Expression and Purification of FMD VLPs
3.2. Structural Characteristics of FMDV VLPs
3.3. Evaluation of the Efficacy of FMD VLPs in Mice After Vaccination
3.4. Virus Challenge of Mice After Vaccination
3.5. Virus Challenge of Swine After Vaccination
4. Discussion
4.1. Immunogenicity and Protective Efficacy of FMDV VLPs
4.2. Broader Implications, Limitations, and Future Perspectives
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| FMDV | Foot-and-mouth disease virus |
| VLP | Virus-like particle |
| VNT | Virus neutralization test |
| TEM | Transmission electron microscopy |
| dpv | Days post-vaccination |
| dpc | Days post-challenge |
| PI | Protection index |
| PD50 | Protective efficacy |
References
- Brown, E.; Nelson, N.; Gubbins, S.; Colenutt, C. Airborne Transmission of Foot-and-Mouth Disease Virus: A Review of Past and Present Perspectives. Viruses 2022, 14, 1009. [Google Scholar] [CrossRef] [PubMed]
- Knight-Jones, T.J.; Rushton, J. The economic impacts of foot and mouth disease—What are they, how big are they and where do they occur? Prev. Vet. Med. 2013, 112, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Yu, S.; Wang, W.; Chen, W.; Wang, X.; Wu, K.; Li, X.; Fan, S.; Ding, H.; Yi, L.; et al. Development of Foot-and-Mouth Disease Vaccines in Recent Years. Vaccines 2022, 10, 1817. [Google Scholar] [CrossRef] [PubMed]
- Kenubih, A. Foot and Mouth Disease Vaccine Development and Challenges in Inducing Long-Lasting Immunity: Trends and Current Perspectives. Vet. Med. 2021, 12, 205–215. [Google Scholar] [CrossRef]
- Grubman, M.J.; Baxt, B. Foot-and-mouth disease. Clin. Microbiol. Rev. 2004, 17, 465–493. [Google Scholar] [CrossRef]
- Diaz-San Segundo, F.; Medina, G.N.; Stenfeldt, C.; Arzt, J.; de Los Santos, T. Foot-and-mouth disease vaccines. Vet. Microbiol. 2017, 206, 102–112. [Google Scholar] [CrossRef]
- Dill, V.; Eschbaumer, M. Cell culture propagation of foot-and-mouth disease virus: Adaptive amino acid substitutions in structural proteins and their functional implications. Virus Genes 2020, 56, 1–15. [Google Scholar] [CrossRef]
- Jamal, S.M.; Belsham, G.J. Foot-and-mouth disease: Past, present and future. Vet. Res. 2013, 44, 116. [Google Scholar] [CrossRef]
- Puckette, M.; Primavera, V.; Martel, E.; Barrera, J.; Hurtle, W.; Clark, B.; Kamicker, B.; Zurita, M.; Brake, D.; Neilan, J. Transiently Transfected Mammalian Cell Cultures: An Adaptable and Effective Platform for Virus-like Particle-Based Vaccines against Foot-and-Mouth Disease Virus. Viruses 2022, 14, 989. [Google Scholar] [CrossRef]
- Puckette, M.; Clark, B.A.; Smith, J.D.; Turecek, T.; Martel, E.; Gabbert, L.; Pisano, M.; Hurtle, W.; Pacheco, J.M.; Barrera, J.; et al. Foot-and-Mouth Disease (FMD) Virus 3C Protease Mutant L127P: Implications for FMD Vaccine Development. J. Virol. 2017, 91, 10–1128. [Google Scholar] [CrossRef]
- Guo, H.C.; Sun, S.Q.; Jin, Y.; Yang, S.L.; Wei, Y.Q.; Sun, D.H.; Yin, S.H.; Ma, J.W.; Liu, Z.X.; Guo, J.H.; et al. Foot-and-mouth disease virus-like particles produced by a SUMO fusion protein system in Escherichia coli induce potent protective immune responses in guinea pigs, swine and cattle. Vet. Res. 2013, 44, 48. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Zhang, S.; Yan, H.; Geng, X.; Wang, Y.; Xu, X.; Wang, M.; Zhang, H.; Huang, B.; Pang, W.; et al. The High Immunity Induced by the Virus-Like Particles of Foot-and-Mouth Disease Virus Serotype O. Front. Vet. Sci. 2021, 8, 633706. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Lu, Y.; Zhang, Y.; Mu, S.; Wang, N.; Du, P.; Zhi, X.; Wen, X.; Wang, X.; Sun, S.; et al. A Heat-Induced Mutation on VP1 of Foot-and-Mouth Disease Virus Serotype O Enhanced Capsid Stability and Immunogenicity. J. Virol. 2021, 95, e0017721. [Google Scholar] [CrossRef] [PubMed]
- Tahara, N.; Tachibana, I.; Takeo, K.; Yamashita, S.; Shimada, A.; Hashimoto, M.; Ohno, S.; Yokogawa, T.; Nakagawa, T.; Suzuki, F.; et al. Boosting Auto-Induction of Recombinant Proteins in Escherichia coli with Glucose and Lactose Additives. Protein Pept. Lett. 2021, 28, 1180–1190. [Google Scholar] [CrossRef]
- Kim, A.Y.; Park, S.Y.; Park, S.H.; Kim, J.Y.; Jin, J.S.; Kim, E.S.; Park, J.H.; Ko, Y.J. Comparison of High-Performance Liquid Chromatography with Sucrose Density Gradient Ultracentrifugation for the Quantification of Foot-and-Mouth Disease Vaccine Antigens. Vaccines 2022, 10, 667. [Google Scholar] [CrossRef]
- Yang, Y.; Li, H.; Li, Z.; Zhang, Y.; Zhang, S.; Chen, Y.; Yu, M.; Ma, G.; Su, Z. Size-exclusion HPLC provides a simple, rapid, and versatile alternative method for quality control of vaccines by characterizing the assembly of antigens. Vaccine 2015, 33, 1143–1150. [Google Scholar] [CrossRef]
- Fukai, K.; Morioka, K.; Yamada, M.; Nishi, T.; Yoshida, K.; Kitano, R.; Yamazoe, R.; Kanno, T. Comparative performance of fetal goat tongue cell line ZZ-R 127 and fetal porcine kidney cell line LFBK-alphavbeta6 for Foot-and-mouth disease virus isolation. J. Vet. Diagn. Investig. 2015, 27, 516–521. [Google Scholar] [CrossRef]
- Xiao, Y.; Chen, H.Y.; Wang, Y.; Yin, B.; Lv, C.; Mo, X.; Yan, H.; Xuan, Y.; Huang, Y.; Pang, W.; et al. Large-scale production of foot-and-mouth disease virus (serotype Asia1) VLP vaccine in Escherichia coli and protection potency evaluation in cattle. BMC Biotechnol. 2016, 16, 56. [Google Scholar] [CrossRef]
- Le, N.M.T.; So, K.K.; Chun, J.; Kim, D.H. Expression of virus-like particles (VLPs) of foot-and-mouth disease virus (FMDV) using Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2024, 108, 81. [Google Scholar] [CrossRef]
- Kheirvari, M.; Liu, H.; Tumban, E. Virus-like Particle Vaccines and Platforms for Vaccine Development. Viruses 2023, 15, 1109. [Google Scholar] [CrossRef]
- Zhou, Y.; Yin, W.; Teng, Z.; Zhao, Y.; Lu, Y.; Qian, Y.; Deng, B. Enhanced Immunogenicity of Foot-and-Mouth Disease Virus-like Particles Using a Water-in-Oil-in-Water Adjuvant. Vaccines 2024, 13, 24. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, R.; Borisova, G.P.; Gren, E.; Berzin, I.; Pumpen, P.; Eckert, R.; Ose, V.; Siakkou, H.; Gren, E.J.; von Baehr, R.; et al. Immunogenicity of recombinant core particles of hepatitis B virus containing epitopes of human immunodeficiency virus 1 core antigen. Arch. Virol. 1992, 126, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Lua, L.H.; Connors, N.K.; Sainsbury, F.; Chuan, Y.P.; Wibowo, N.; Middelberg, A.P. Bioengineering virus-like particles as vaccines. Biotechnol. Bioeng. 2014, 111, 425–440. [Google Scholar] [CrossRef] [PubMed]
- Arzt, J.; Belsham, G.J.; Lohse, L.; Botner, A.; Stenfeldt, C. Transmission of Foot-and-Mouth Disease from Persistently Infected Carrier Cattle to Naive Cattle via Transfer of Oropharyngeal Fluid. mSphere 2018, 3, 10–1128. [Google Scholar] [CrossRef]
- Alexandersen, S.; Zhang, Z.; Donaldson, A.I.; Garland, A.J. The pathogenesis and diagnosis of foot-and-mouth disease. J. Comp. Pathol. 2003, 129, 1–36. [Google Scholar] [CrossRef]
- Reeve, R.; Borley, D.W.; Maree, F.F.; Upadhyaya, S.; Lukhwareni, A.; Esterhuysen, J.J.; Harvey, W.T.; Blignaut, B.; Fry, E.E.; Parida, S.; et al. Tracking the Antigenic Evolution of Foot-and-Mouth Disease Virus. PLoS ONE 2016, 11, e0159360. [Google Scholar] [CrossRef]
- Alexandersen, S.; Zhang, Z.; Donaldson, A.I. Aspects of the persistence of foot-and-mouth disease virus in animals--the carrier problem. Microbes Infect. 2002, 4, 1099–1110. [Google Scholar] [CrossRef]
- Stenfeldt, C.; Arzt, J. The Carrier Conundrum; A Review of Recent Advances and Persistent Gaps Regarding the Carrier State of Foot-and-Mouth Disease Virus. Pathogens 2020, 9, 167. [Google Scholar] [CrossRef]
- Rweyemamu, M.; Roeder, P.; MacKay, D.; Sumption, K.; Brownlie, J.; Leforban, Y. Planning for the progressive control of foot-and-mouth disease worldwide. Transbound. Emerg. Dis. 2008, 55, 73–87. [Google Scholar] [CrossRef]
- Li, H.; Liu, P.; Dong, H.; Dekker, A.; Harmsen, M.M.; Guo, H.; Wang, X.; Sun, S. Foot-and-mouth disease virus antigenic landscape and reduced immunogenicity elucidated in atomic detail. Nat. Commun. 2024, 15, 8774. [Google Scholar] [CrossRef]
- Wang, J.W.; Roden, R.B. Virus-like particles for the prevention of human papillomavirus-associated malignancies. Expert Rev. Vaccines 2013, 12, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Moradi Vahdat, M.; Hemmati, F.; Ghorbani, A.; Rutkowska, D.; Afsharifar, A.; Eskandari, M.H.; Rezaei, N.; Niazi, A. Hepatitis B core-based virus-like particles: A platform for vaccine development in plants. Biotechnol. Rep. 2021, 29, e00605. [Google Scholar] [CrossRef] [PubMed]
- Dhawan, M.; Saied, A.A.; Sharma, M. Virus-like particles (VLPs)-based vaccines against COVID-19: Where do we stand amid the ongoing evolution of SARS-CoV-2? Health Sci. Rev. 2023, 9, 100127. [Google Scholar] [CrossRef]
- Mignaqui, A.C.; Ferella, A.; Cass, B.; Mukankurayija, L.; L’Abbe, D.; Bisson, L.; Sanchez, C.; Scian, R.; Cardillo, S.B.; Durocher, Y.; et al. Foot-and-Mouth Disease: Optimization, Reproducibility, and Scalability of High-Yield Production of Virus-Like Particles for a Next-Generation Vaccine. Front. Vet. Sci. 2020, 7, 601. [Google Scholar] [CrossRef]
- Lomonossoff, G.P.; D’Aoust, M.A. Plant-produced biopharmaceuticals: A case of technical developments driving clinical deployment. Science 2016, 353, 1237–1240. [Google Scholar] [CrossRef]
- Doel, T.R. FMD vaccines. Virus Res. 2003, 91, 81–99. [Google Scholar] [CrossRef]
- Mahapatra, M.; Parida, S. Foot and mouth disease vaccine strain selection: Current approaches and future perspectives. Expert Rev. Vaccines 2018, 17, 577–591. [Google Scholar] [CrossRef]
- Liu, X.; Fang, Y.; Zhou, P.; Lu, Y.; Zhang, Q.; Xiao, S.; Dong, Z.; Pan, L.; Lv, J.; Zhang, Z.; et al. Chimeric virus-like particles elicit protective immunity against serotype O foot-and-mouth disease virus in guinea pigs. Appl. Microbiol. Biotechnol. 2017, 101, 4905–4914. [Google Scholar] [CrossRef]
- Gupta, R.; Arora, K.; Roy, S.S.; Joseph, A.; Rastogi, R.; Arora, N.M.; Kundu, P.K. Platforms, advances, and technical challenges in virus-like particles-based vaccines. Front. Immunol. 2023, 14, 1123805. [Google Scholar] [CrossRef]
- Mohana Subramanian, B.; Madhanmohan, M.; Sriraman, R.; Chandrasekhar Reddy, R.V.; Yuvaraj, S.; Manikumar, K.; Rajalakshmi, S.; Nagendrakumar, S.B.; Rana, S.K.; Srinivasan, V.A. Development of foot-and-mouth disease virus (FMDV) serotype O virus-like-particles (VLPs) vaccine and evaluation of its potency. Antivir. Res. 2012, 96, 288–295. [Google Scholar] [CrossRef]
- Mason, P.W.; Piccone, M.E.; McKenna, T.S.; Chinsangaram, J.; Grubman, M.J. Evaluation of a live-attenuated foot-and-mouth disease virus as a vaccine candidate. Virology 1997, 227, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Belsham, G.J. Towards improvements in foot-and-mouth disease vaccine performance. Acta Vet. Scand. 2020, 62, 20. [Google Scholar] [CrossRef] [PubMed]
- Rathogwa, N.M.; Scott, K.A.; Opperman, P.; Theron, J.; Maree, F.F. Efficacy of SAT2 Foot-and-Mouth Disease Vaccines Formulated with Montanide ISA 206B and Quil-A Saponin Adjuvants. Vaccines 2021, 9, 996. [Google Scholar] [CrossRef] [PubMed]
- Porta, C.; Xu, X.; Loureiro, S.; Paramasivam, S.; Ren, J.; Al-Khalil, T.; Burman, A.; Jackson, T.; Belsham, G.J.; Curry, S.; et al. Efficient production of foot-and-mouth disease virus empty capsids in insect cells following down regulation of 3C protease activity. J. Virol. Methods 2013, 187, 406–412. [Google Scholar] [CrossRef]
- Zhang, C.; Ku, Z.; Liu, Q.; Wang, X.; Chen, T.; Ye, X.; Li, D.; Jin, X.; Huang, Z. High-yield production of recombinant virus-like particles of enterovirus 71 in Pichia pastoris and their protective efficacy against oral viral challenge in mice. Vaccine 2015, 33, 2335–2341. [Google Scholar] [CrossRef]
- Chung, Y.C.; Ho, M.S.; Wu, J.C.; Chen, W.J.; Huang, J.H.; Chou, S.T.; Hu, Y.C. Immunization with virus-like particles of enterovirus 71 elicits potent immune responses and protects mice against lethal challenge. Vaccine 2008, 26, 1855–1862. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serotype | SP-ELISA PI (%) | VNT (1:X) | ||
|---|---|---|---|---|
| 21 dpv | 42 dpv | 21 dpv | 42 dpv | |
| A-type | <50% | >50% | 112 | 676 |
| O-type | >50% | >50% | 143 | 767 |
| Asia1-type | >50% | >50% | 47 | 295 |
| Serotype | PD50 | |
|---|---|---|
| Mice | Swine | |
| A-type | 73.5 | – |
| O-type | 32.0 | 10.6 |
| Asia1-type | 55.7 | 22.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, S.-C.; Lee, I.-K.; Kong, H.-S.; Shin, S.-H.; Hwang, S.-Y.; Ahn, Y.-J.; Park, J.-H.; Kim, B.-Y.; Song, Y.-C. Foot-and-Mouth Disease Virus-like Particles Produced in E. coli as Potential Antigens for a Novel Vaccine. Vet. Sci. 2025, 12, 539. https://doi.org/10.3390/vetsci12060539
Yu S-C, Lee I-K, Kong H-S, Shin S-H, Hwang S-Y, Ahn Y-J, Park J-H, Kim B-Y, Song Y-C. Foot-and-Mouth Disease Virus-like Particles Produced in E. coli as Potential Antigens for a Novel Vaccine. Veterinary Sciences. 2025; 12(6):539. https://doi.org/10.3390/vetsci12060539
Chicago/Turabian StyleYu, Sang-Cheol, In-Kyu Lee, Hyun-Seok Kong, Sung-Ho Shin, Sung-Yoon Hwang, Yu-Jin Ahn, Jong-Hyeon Park, Bong-Yoon Kim, and Young-Cheon Song. 2025. "Foot-and-Mouth Disease Virus-like Particles Produced in E. coli as Potential Antigens for a Novel Vaccine" Veterinary Sciences 12, no. 6: 539. https://doi.org/10.3390/vetsci12060539
APA StyleYu, S.-C., Lee, I.-K., Kong, H.-S., Shin, S.-H., Hwang, S.-Y., Ahn, Y.-J., Park, J.-H., Kim, B.-Y., & Song, Y.-C. (2025). Foot-and-Mouth Disease Virus-like Particles Produced in E. coli as Potential Antigens for a Novel Vaccine. Veterinary Sciences, 12(6), 539. https://doi.org/10.3390/vetsci12060539