The Pivotal Interaction Between Serotonin and Calcium Shifts in Lactating Pregnant Spanish Purebred Mares: The Aging Effect

 , , , , ,
, , , , ,  and
and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mares
2.2. Blood Samples and Analyses
2.3. Analytical Procedures
2.4. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Portbury, A.L.; Chandra, R.; Groelle, M.; McMillian, M.K.; Elias, A.; Herlong, J.R.; Chikaraishi, D.M. Catecholamines act via a beta-adrenergic receptor to maintain fetal heart rate and survival. Am. J. Physiol. Heart Circ. Physiol. 2003, 284, 2069–2077. [Google Scholar] [CrossRef] [PubMed]
- Torfs, S.C.; Maes, A.A.; Delesalle, C.J.; Deprez, P.; Croubels, S.M. Comparative analysis of serotonin in equine plasma with liquid chromatography-tandem mass spectrometry and enzyme-linked immunosorbent assay. J. Vet. Diagn. Investig. 2012, 24, 1035–1042. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Toyofuku, Y.; Lynn, F.C.; Chak, E.; Uchida, T.; Mizukami, H.; Fujitani, Y.; Kawamori, R.; Miyatsuka, T.; Kosaka, Y.; et al. Serotonin regulates pancreatic beta cell mass during pregnancy. Nat. Med. 2010, 16, 804–808. [Google Scholar] [CrossRef] [PubMed]
- Marcilla, M.; Muñoz, A.; Satué, K. Longitudinal changes in serum catecholamines, dopamine, serotonin, ACTH and cortisol in pregnant Spanish mares. Res. Vet. Sci. 2017, 115, 29–33. [Google Scholar] [CrossRef]
- Wysolmerski, J.J. Parathyroid hormone-related protein: An update. J. Clin. Endocrinol. Metab. 2012, 97, 2947–2956. [Google Scholar] [CrossRef]
- Horseman, N.D.; Collier, R.J. Serotonin: A local regulator in the mammary gland epithelium. Annu. Rev. Anim. Biosci. 2014, 2, 353–374. [Google Scholar] [CrossRef]
- Weaver, S.R.; Laporta, J.; Moore, S.A.E.L.L.; Hernandez, L.L. Serotonin and calcium homeostasis during the transition period. Dom. Anim. Endocrinol. 2016, 56, S147–S154. [Google Scholar] [CrossRef]
- Connelly, M.K.; Cheng, A.A.; Hernandez, L.L. Graduate Student Literature Review: Serotonin and calcium metabolism: A story unfolding. J. Dairy Sci. 2021, 104, 13008–13019. [Google Scholar] [CrossRef]
- Laporta, J.; Moore, S.A.E.; Weaver, S.R.; Cronick, C.M.; Olsen, M.; Prichard, A.P.; Schnell, B.P.; Crenshaw, T.D.; Penagaricano, F.; Bruckmaier, R.M.; et al. Increasing serotonin concentrations alter calcium and energy metabolism in dairy cows. J. Endocrinol. 2015, 226, 43–55. [Google Scholar] [CrossRef]
- Harvey, J.W.; Pate, M.G.; Kivipelto, J.; Asquith, R.L. Clinical biochemistry of pregnant and nursing mares. Vet. Clin. Pathol. 2005, 34, 248–254. [Google Scholar] [CrossRef]
- Holley, D.C.; Evans, J.W. Determination of total and ultrafiltrabile calcium and magnesium in normal equine serum. Am. J. Vet. Res. 1977, 38, 259–262. [Google Scholar] [CrossRef] [PubMed]
- Martin, K.L.; Hoffman, R.M.; Kronfeld, D.S.; Ley, W.B.; Warnick, L.D. Calcium decreases and parathyroid hormone increases in serum of periparturient mares. J. Anim. Sci. 1996, 74, 834–839. [Google Scholar] [CrossRef] [PubMed]
- Stojević, F.N.; Prvanović, Z.; Zvonimir, T.N. The influence of late pregnancy and lactation on bone metabolism in mares. Res. Vet. Sci. 2010, 88, 405–410. [Google Scholar]
- De Palo, P.; Auclair-Ronzaud, J.; Maggiolino, A. Mammary gland physiology and farm management of dairy mares and jennies. JDS Commun. 2022, 3, 234–237. [Google Scholar] [CrossRef]
- Guo, J.; Huang, X.; Dou, L.; Yan, M.; Shen, T.; Tang, W.; Li, J. Aging and aging-related diseases: From molecular mechanisms to interventions and treatments. Signal Transduct. Target. Ther. 2022, 7, 391. [Google Scholar] [CrossRef] [PubMed]
- Satué, K.; Fazio, E.; Damiá, E.; Barbiera, G.; Medica, P.; Cravana, C. Effect of age on androgen pattern in cyclic mares. Res. Vet. Sci. 2024, 173, 103276. [Google Scholar] [CrossRef]
- Satué, K.; Fazio, E.; Velasco-Martínez, M.G.; La Fauci, D.; Barbiera, G.; Medica, P.; Cravana, C. Can the reduced GH, IGH-1, and ovarian steroids concentrations be considered as suspected biomarkers of age-associated functional deficit in mares? Theriogenology 2024, 223, 75–80. [Google Scholar] [CrossRef]
- Berlin, D.; Aroch, I. Concentrations of ionized and total magnesium and calcium in healthy horses: Effects of age, pregnancy, lactation, pH and sample type. Vet. J. 2009, 181, 305–311. [Google Scholar] [CrossRef]
- Holcombe, S.J.; Embertson, R.M.; Kurtz, K.A.; Roessner, H.A.; Wismer, S.E.; Geor, R.J.; Kaneene, J.B. Increased serum nonesterified fatty acid and low ionised calcium concentrations are associated with post partum colic in mares. Equine Vet. J. 2016, 48, 39–44. [Google Scholar] [CrossRef]
- Satué, K.; Fazio, E.; Velasco-Martínez, M.G.; La Fauci, D.; Cravana, C.; Medica, P. Effect of age on amplitude of circulating catecholamine’s change of healthy cyclic mares. Vet. Res. Commun. 2024, 48, 2863–2868. [Google Scholar] [CrossRef]
- Andersen, J.T.; Andersen, N.L.; Horwitz, H.; Poulsen, H.E.; Jimenez-Solem, E. Exposure to serotonin reuptake inhibitors in early pregnancy and the risk of miscarriage. Obstet. Gynecol. 2014, 124, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Richardson, J.D.; Harrison, L.J.; Edwards, G.B. Two horses with hypocalcaemia. Vet. Rec. 1991, 129, 98. [Google Scholar] [CrossRef] [PubMed]
- McArt, J.A.A.; Oetzel, G.R. Considerations in the Diagnosis and Treatment of Early Lactation Calcium Disturbances. Vet. Clin. North Am. Food Anim. Pract. 2023, 39, 241–259. [Google Scholar] [CrossRef]
- Muñoz, A.; Riber, C.; Trigo, P.; Castejón, F. Age- and gender-related variations in hematology, clinical biochemistry, and hormones in Spanish fillies and colts. Res. Vet. Sci. 2012, 93, 943–949. [Google Scholar] [CrossRef]
- van der Kolk, J.H.; Nachreiner, R.F.; Refsal, K.R.; Brouillet, D.; Wensing, T. Heparinised blood ionised calcium concentrations in horses with colic or diarrhoea compared to nor-mal subjects. Equine Vet. J. 2002, 34, 528–531. [Google Scholar] [CrossRef] [PubMed]
- Lopez, I.; Estepa, J.C.; Mendoza, F.J.; Mayer-Valor, R.; Aguilera-Tejero, E. Fractionation of calcium and magnesium in equine serum. Am. J. Vet. Res. 2006, 67, 463–466. [Google Scholar] [CrossRef]
- Schumacher, S.A.; Yardley, J.; Bertone, A.L. Ionized magnesium and calcium concentration and their ratio in equine plasma samples as determined by a regulatory laboratory compared to a clinical reference laboratory. Drug Test. Anal. 2019, 11, 455–460. [Google Scholar] [CrossRef]
- Aguilera-Tejero, E.; Estepa, J.C.; López, I.; Bas, S.; Garfia, B.; Rodríguez, M. Plasma ionized calcium and parathyroid hormone concentrations in horses after endurance rides. J. Am. Vet. Med. Assoc. 2001, 219, 488–490. [Google Scholar] [CrossRef]
- Sanmartí, J.; Robles-Guirado, J.A.; Jose-Cunilleras, E.; Bassols, A. Sample stability and heparin interference in ionized calcium and ionized magnesium measurements in horses using the Stat Profile Prime Plus co-oximetry electrolyte analyzer. Vet. Clin. Pathol. 2023, 52, 252–260. [Google Scholar] [CrossRef]
- Kovacs, C.S. Maternal mineral and bone metabolism during pregnancy, lactation, and post-weaning recovery. Physiol. Rev. 2016, 96, 449–547. [Google Scholar] [CrossRef]
- Almaghamsi, A.; Almalki, M.H.; Buhary, B.M. Hypocalcemia in Pregnancy: A Clinical Review Update. Oman Med. J. 2018, 33, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Ajong, A.B.; Kenfack, B.; Ali, I.M.; Yakum, M.N.; Onydinma, U.P.; Mangala, F.N.; Aljerf, L.; Telefo, P.B. Ionised and total hypocalcaemia in pregnancy: An analysis of prevalence and risk factors in a resource-limited setting, Cameroon. PLoS ONE 2022, 17, e0268643. [Google Scholar] [CrossRef] [PubMed]
- Goltzman, D. Physiology of parathyroid hormone. Endocrinol. Metab. Clin. North Am. 2018, 47, 743–758. [Google Scholar] [CrossRef]
- Wooding, F.B.P.; Morgan, G.; Fowden, A.L.; Allen, W.R. Separate sites and mechanisms of placental transport of calcium, iron and glucose in the equine placenta. Placenta 2000, 21, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Melendez, P.; Lopez, F.; Lama, J.; Leon, B.; Pinedo, P. Plasma ionized calcium and magnesium concentrations and prevalence of subclinical hypocalcemia and hypomagnesemia in postpartum grazing Holstein cows from southern Chile. Vet. Anim. Sci. 2023, 19, 100277. [Google Scholar] [CrossRef]
- Acevedo-Rodriguez, E.M.; Aris, A.; Bach, A. Associations between subclinical hypocalcemia and postparturient diseases in dairy cows. J. Dairy Sci. 2017, 100, 7427–7434. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulou, A.; Bountouvi, E.; Karachaliou, F.E. The molecular basis of calcium and phosphorus inherited metabolic disorders. Genes 2021, 12, 734. [Google Scholar] [CrossRef]
- Zepperitz, H.; Gürtler, H. Ionisiertes Calcium und Gesamtcalcium im Blut von Rindern, Schafen, Schweinen und Pferden verschiedener Alters- und Reproduktionsstadien und Nutzungsrichtungen [Ionized calcium and total calcium in the blood of cattle, sheep, swine and horses of different ages, reproductive stages and uses]. Berl. Munch Tierarztl. Wochensch. 1992, 105, 328–332. [Google Scholar]
- Terrell, K.; Suyun Choi, S.; Choi, S. Calcium’s Role and Signaling in Aging Muscle, Cellular Senescence, and Mineral Interactions. Int. J. Mol. Sci. 2023, 24, 17034. [Google Scholar] [CrossRef]
- Bauman, D.E.; Currie, W.B. Partitioning of nutrients during pregnancy and lactation: A review of mechanisms involving homeostasis and homeorhesis. J. Dairy Sci. 1980, 63, 1514–1529. [Google Scholar] [CrossRef]
- VanHouten, J.N.; Dann, P.; Stewart, A.F.; Watson, C.J.; Pollak, M.; Karaplis, A.C.; Wysolmerski, J.J. Mammary-specific deletion of parathyroid hormone-related protein preserves bone mass during lactation. J. Clin. Investig. 2023, 112, 1429–1436. [Google Scholar] [CrossRef]
- Laporta, J.; Keil, K.P.; Vezina, C.M.; Hernandez, L.L. Peripheral serotonin regulates maternal calcium trafficking in mammary epithelial cells during lactation in mice. PLoS ONE 2014, 9, e110190. [Google Scholar] [CrossRef]
- Bornstein, S.; Brown, S.A.; Le, P.T.; Wang, X.; DeMambro, V.; Horowitz, M.C.; MacDougald, O.; Baron, R.; Lotinun, S.; Karsenty, G.; et al. FGF-21 and skeletal remodeling during and after lactation in C57BL/6JMice. Endocrinology 2014, 155, 3516–3526. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.R.; Jury, N.J.; Gregerson, K.A.; Horseman, N.D.; Hernandez, L.L. Characterization of mammary-specific disruptions for Tph1 and Lrp5 during murine lactation. Sci. Rep. 2017, 7, 15155. [Google Scholar] [CrossRef]
- Gumusoglu, S.; Scroggins, S.; Vignato, J.; Santillan, D.; Santillan, M. The serotonin-immune axis in preeclampsia. Curr. Hypertens. Rep. 2021, 23, 37. [Google Scholar] [CrossRef] [PubMed]
- Bonnin, A.; Levitt, P. Fetal, maternal, and placental sources of serotonin and new implications for developmental programming of the brain. Neuroscience 2011, 197, 1–7. [Google Scholar] [CrossRef]
- Bonnin, A.; Goeden, N.; Wilson, M.L.; King, J.; Shih, J.C.; Blakely, R.D.; Deneris, E.S.; Levitt, P. A transient placental source of serotonin for the fetal forebrain. Nature 2011, 472, 347–350. [Google Scholar] [CrossRef]
- Hadden, C.; Fahmi, T.; Cooper, A.; Savenka, A.V.; Lupashin, V.V.; Roberts, D.J.; Maroteaux, L.; Hauguel-de Mouzon, S.; Kilic, F. Serotonin transporter protects the placental cells against apoptosis in caspase 3-independent pathway. J. Cell. Physiol. 2017, 232, 3520–3529. [Google Scholar] [CrossRef]
- Rosenfeld, C.S. Placental serotonin signaling, pregnancy outcomes, and regulation of fetal brain development. Biol. Reprod. 2020, 13, 532–538. [Google Scholar] [CrossRef]
- Ginther, O.J.; Gastal, E.L.; Gastal, M.O.; Bergfelt, D.R.; Baerwald, A.R.; Pierson, R.A. Comparative study of the dynamics of follicular waves in mares and women. Biol. Reprod. 2004, 71, 1195–1201. [Google Scholar] [CrossRef]
- Carnevale, E.M. The mare model for follicular maturation and reproductive aging in the woman. Theriogenology 2008, 69, 23–30. [Google Scholar] [CrossRef]
- Hernandez-Castellano, L.E.; Hernandez, L.L.; Weaver, S.; Bruckmaier, R.M. Increased se-rum serotonin improves parturient calcium homeostasis in dairy cows. J. Dairy Sci. 2017, 100, 1580–1587. [Google Scholar] [CrossRef] [PubMed]
- Rodney, R.M.; Martinez, N.; Block, E.; Hernandez, L.L.; Celi, P.; Nelson, C.D.; Santos, J.; Lean, I.J. Effects of prepartum dietary cation-anion difference and source of vitamin D in dairy cows: Vitamin D, mineral, and bone metabolism. J. Dairy Sci. 2018, 101, 2519–2543. [Google Scholar] [CrossRef]
- Slater, C.J.; Endres, E.L.; Weaver, S.R.; Cheng, A.A.; Lauber, M.R.; Endres, S.F.; Olstad, E.; DeBruin, A.; Crump, P.M.; Block, E.; et al. Interaction of 5-hydroxy-l-tryptophan and negative dietary cation-anion difference on calcium homeostasis in multiparous peri-partum dairy cows. J. Dairy Sci. 2018, 101, 5486–5501. [Google Scholar] [CrossRef]
- Musumeci, G.; Loreto, C.; Trovato, F.M.; Giunta, S.; Imbesi, R.; Castrogiovanni, P. Serotonin (5HT) expression in rat pups treated with high-tryptophan diet during fetal and early post-natal development. Acta Histochem. 2014, 116, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Gall, V.; Kosec, V.; Field, T.; Diego, M.; Hernandez-Reif, M.; Figueiredo, B.; Ascencio, A.; Schanberg, S.; Kuhn, C. Prenatal serotonin and neonatal outcome: Brief report. Infant Behav. Dev. 2008, 3, 316–320. [Google Scholar]
- Janssen, J.A.M.J.L. New Insights into the Role of Insulin and Hypothalamic-Pituitary-Adrenal (HPA) Axis in the Metabolic Syndrome. Int. J. Mol. Sci. 2022, 23, 8178. [Google Scholar] [CrossRef]
- Watanabe, H.; Saito, R.; Nakano, T.; Takahashi, H.; Takahashi, Y.; Sumiyoshi, K.; Sato, K.; Ghen, Y.; Okada, N.; Iwasaki, S.; et al. Effect of peripheral 5-HT on glucose and lipid metabolism in wether sheep. PLoS ONE 2014, 9, 88058. [Google Scholar] [CrossRef]
- Lebin, L.G.; Novick, A.M. Selective Serotonin Reuptake Inhibitors (SSRIs) in Pregnancy: An Updated Review on Risks to Mother, Fetus, and Child. Curr. Psychiatry Rep. 2022, 24, 687–695. [Google Scholar] [CrossRef]
- Laporta, J.; Keil, K.P.; Weaver, S.R.; Cronick, C.M.; Prichard, A.P.; Crenshaw, T.D.; Heyne, G.W.; Vezina, C.M.; Lipinski, R.J.; Hernandez, L.L. Serotonin regulates calcium homeostasis in lactation by epigenetic activation of hedgehog signaling. Mol. Endocrinol. 2014, 28, 1866–1874. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
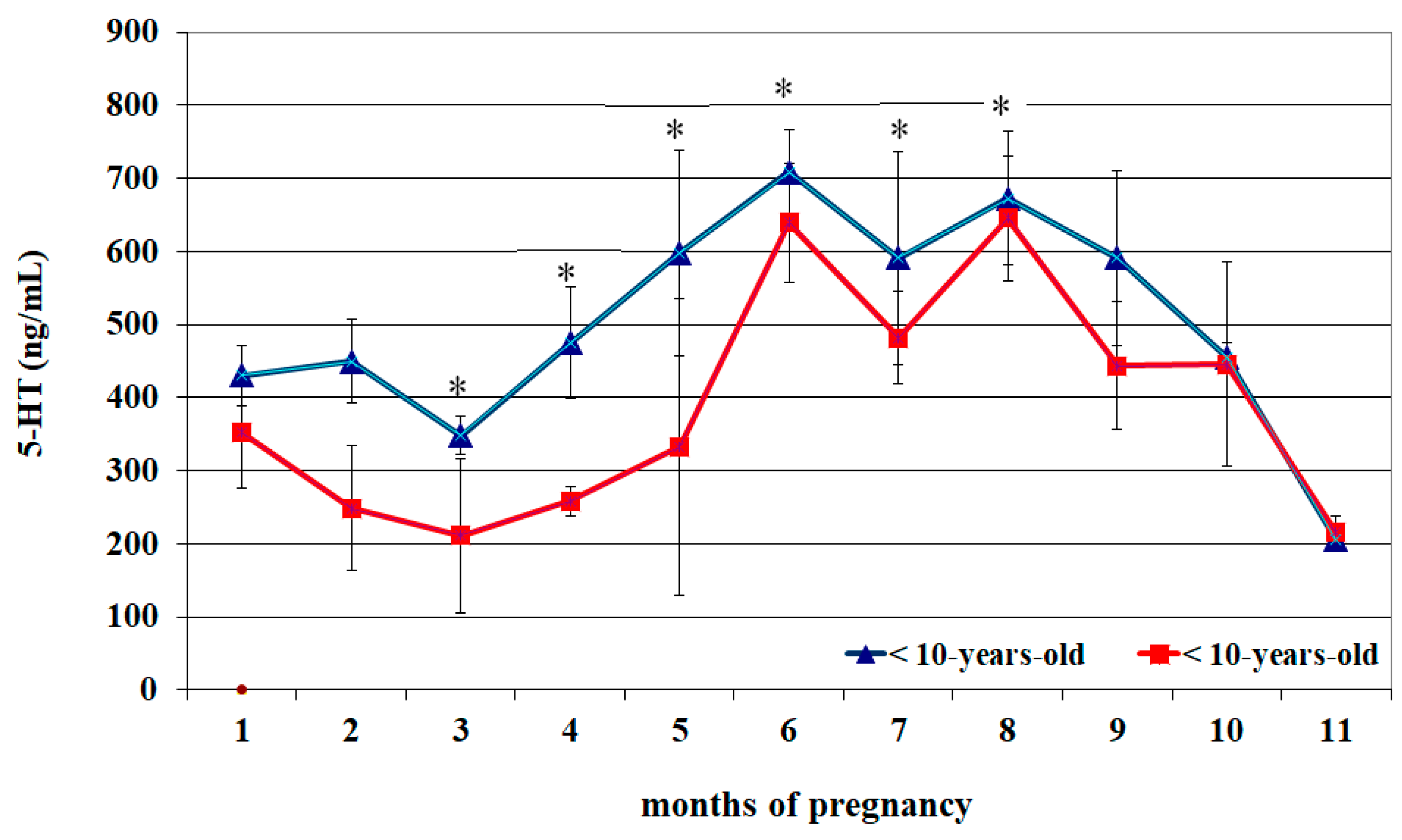
| Month of Pregnancy | 5-HT (ng/mL) (<10 Years Old) | 5-HT (ng/mL) (>10 Years Old) |
|---|---|---|
| 1 | 430.4 ± 41.4 a | 352.9 ± 76.4 A |
| 2 | 450.0 ± 58.8 a | 249.3 ± 85.3 A |
| 3 | 358.6 ± 26.4 a | 211.1 ± 106.9 A |
| 4 | 475.5 ± 76.9 | 259.9 ± 20.9 |
| 5 | 598.1 ± 141.1 | 333.6 ± 203.8 |
| 6 | 709.7 ± 57.1 b | 639.7 ± 81.5 B |
| 7 | 591.9 ± 145.9 | 482.7 ± 63.2 |
| 8 | 673.2 ± 92.8 | 645.9 ± 85.2 |
| 9 | 591.8 ± 119.3 | 444.6 ± 87.7 |
| 10 | 456.6 ± 207.1 | 446.1 ± 139.6 |
| 11 | 205.4 ± 11.1 c | 215.0 ± 23.5 C |
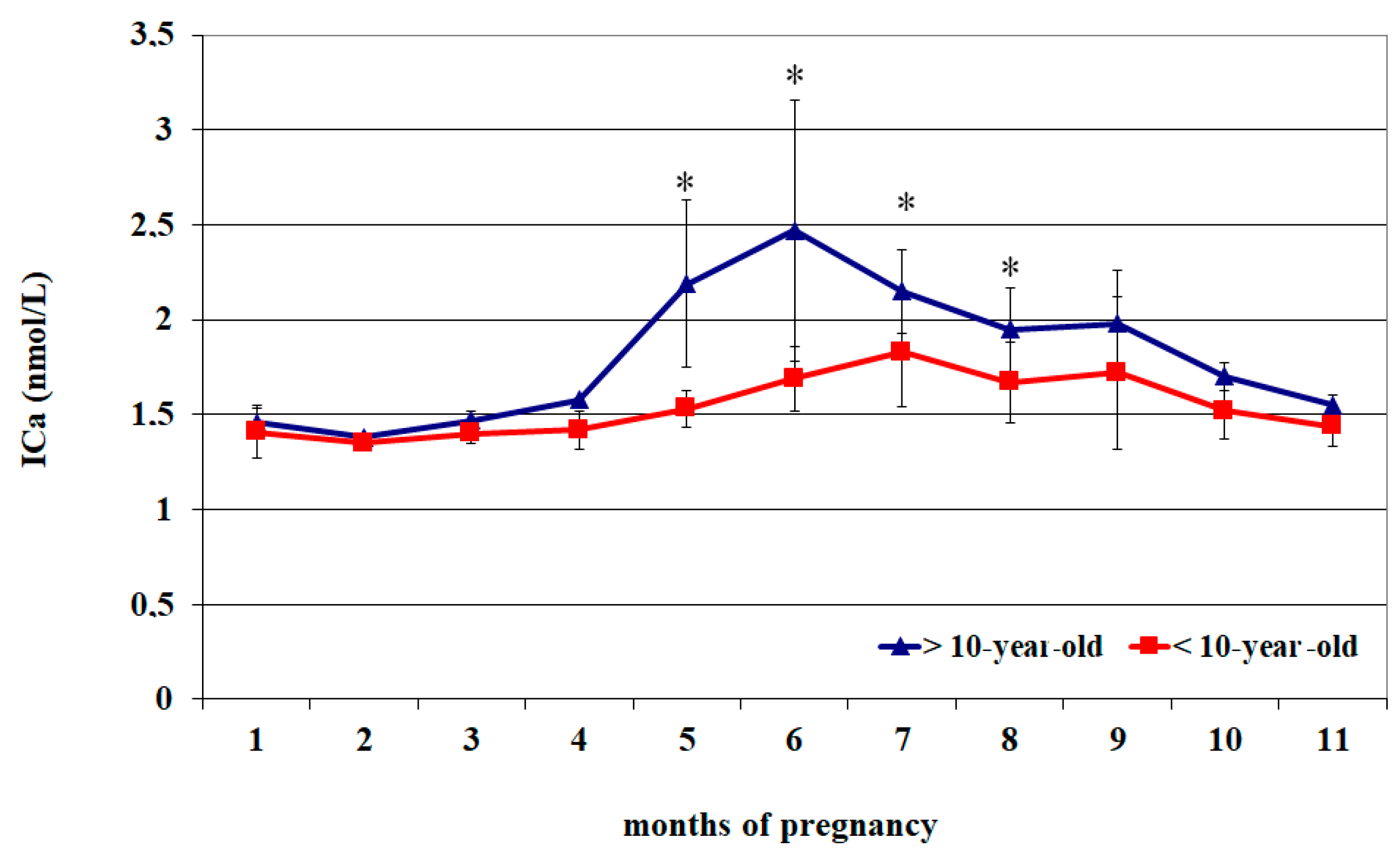
| Month of Pregnancy | ICa2+ (mmol/L) (<10 Years Old) | ICa2+ (mmol/L) (>10 Years Old) |
|---|---|---|
| 1 | 1.46 ± 0.07 a | 1.41 ± 0.14 A |
| 2 | 1.38 ± 0.01 a | 1.35 ± 0.02 A |
| 3 | 1.47 ± 0.05 a | 1.40 ± 0.05 A |
| 4 | 1.58 ± 0.01 a | 1.42 ± 0.10 A |
| 5 | 2.19 ± 0.44 b | 1.53 ± 0.10 |
| 6 | 2.47 ± 0.69 b | 1.69 ± 0.17 |
| 7 | 2.15 ± 0.22 | 1.83 ± 0.29 |
| 8 | 1.95 ± 0.22 | 1.67 ± 0.21 |
| 9 | 1.98 ± 0.28 | 1.72 ± 0.40 |
| 10 | 1.70 ± 0.07 | 1.52 ± 0.15 |
| 11 | 1.55 ± 0.05 a | 1.44 ± 0.11 A |
| Month of Pregnancy | TCa2+ (mmol/L) (<10 Years Old) | TCa2+ (mmol/L) (>10 Years Old) |
|---|---|---|
| 1 | 2.42 ± 0.06 a | 2.58 ± 0.03 A |
| 2 | 2.53 ± 0.09 a | 2.62 ± 0.03 A |
| 3 | 2.62 ± 0.11 a | 2.76 ± 0.07 A |
| 4 | 2.81 ± 0.24 b | 2.83 ± 0.08 B |
| 5 | 2.88 ± 0.13 b | 2.86 ± 0.09 B |
| 6 | 2.55 ± 0.32 | 2.58 ± 0.15 |
| 7 | 2.62 ± 0.23 | 2.31 ± 0.14 |
| 8 | 2.53 ± 0.17 | 2.23 ± 0.17 |
| 9 | 2.74 ± 0.15 c | 2.65 ± 0.1 C |
| 10 | 2.38 ± 0.13 | 2.16 ± 0.13 |
| 11 | 2.20 ± 0.15 | 2.05 ± 0.09 |
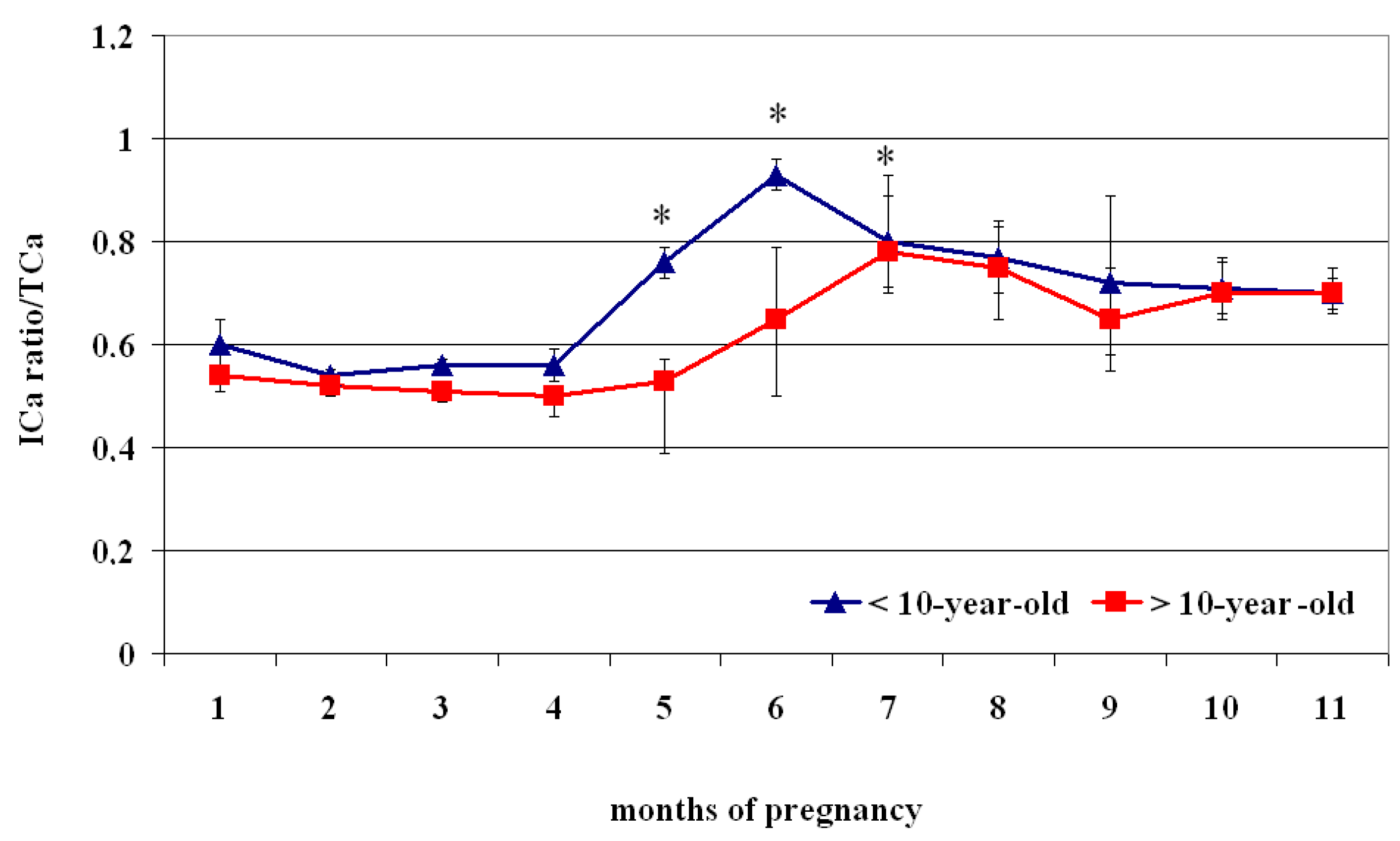
| Month of Pregnancy | ICa2+/TCa2+ Ratio (<10 Years Old) | ICa2+/TCa2+ Ratio (>10 Years Old) |
|---|---|---|
| 1 | 060 ± 0.03 a | 0.54 ± 0.05 A |
| 2 | 0.54 ± 0.02 b | 0.52 ± 0.01 A |
| 3 | 0.56 ± 0.02 b | 0.51 ± 0.01 A |
| 4 | 0.56 ± 0.04 b | 0.50 ± 0.03 A |
| 5 | 0.76 ± 0.14 c | 0.53 ± 0.03 A |
| 6 | 0.93 ± 0.15 d | 0.65 ± 0.03 |
| 7 | 0.80 ± 0.08 | 0.78 ± 0.09 |
| 8 | 0.77 ± 0.10 | 0.75 ± 0.07 |
| 9 | 0.72 ± 0.07 | 0.65 ± 0.17 |
| 10 | 0.71 ± 0.05 | 0.70 ± 0.05 |
| 11 | 0.70 ± 0.04 | 0.70 ± 0.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Satué, K.; Fazio, E.; Medica, P.; Velasco-Martinez, M.G.; Cravana, C.; Bruschetta, G.; La Fauci, D. The Pivotal Interaction Between Serotonin and Calcium Shifts in Lactating Pregnant Spanish Purebred Mares: The Aging Effect. Vet. Sci. 2025, 12, 398. https://doi.org/10.3390/vetsci12050398
Satué K, Fazio E, Medica P, Velasco-Martinez MG, Cravana C, Bruschetta G, La Fauci D. The Pivotal Interaction Between Serotonin and Calcium Shifts in Lactating Pregnant Spanish Purebred Mares: The Aging Effect. Veterinary Sciences. 2025; 12(5):398. https://doi.org/10.3390/vetsci12050398
Chicago/Turabian StyleSatué, Katiuska, Esterina Fazio, Pietro Medica, Maria Gemma Velasco-Martinez, Cristina Cravana, Giuseppe Bruschetta, and Deborah La Fauci. 2025. "The Pivotal Interaction Between Serotonin and Calcium Shifts in Lactating Pregnant Spanish Purebred Mares: The Aging Effect" Veterinary Sciences 12, no. 5: 398. https://doi.org/10.3390/vetsci12050398
APA StyleSatué, K., Fazio, E., Medica, P., Velasco-Martinez, M. G., Cravana, C., Bruschetta, G., & La Fauci, D. (2025). The Pivotal Interaction Between Serotonin and Calcium Shifts in Lactating Pregnant Spanish Purebred Mares: The Aging Effect. Veterinary Sciences, 12(5), 398. https://doi.org/10.3390/vetsci12050398