

Distribution and Prevalence of Ticks and Tick-Borne Pathogens at the Wildlife-Livestock Interface in Africa: A Systematic Review


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
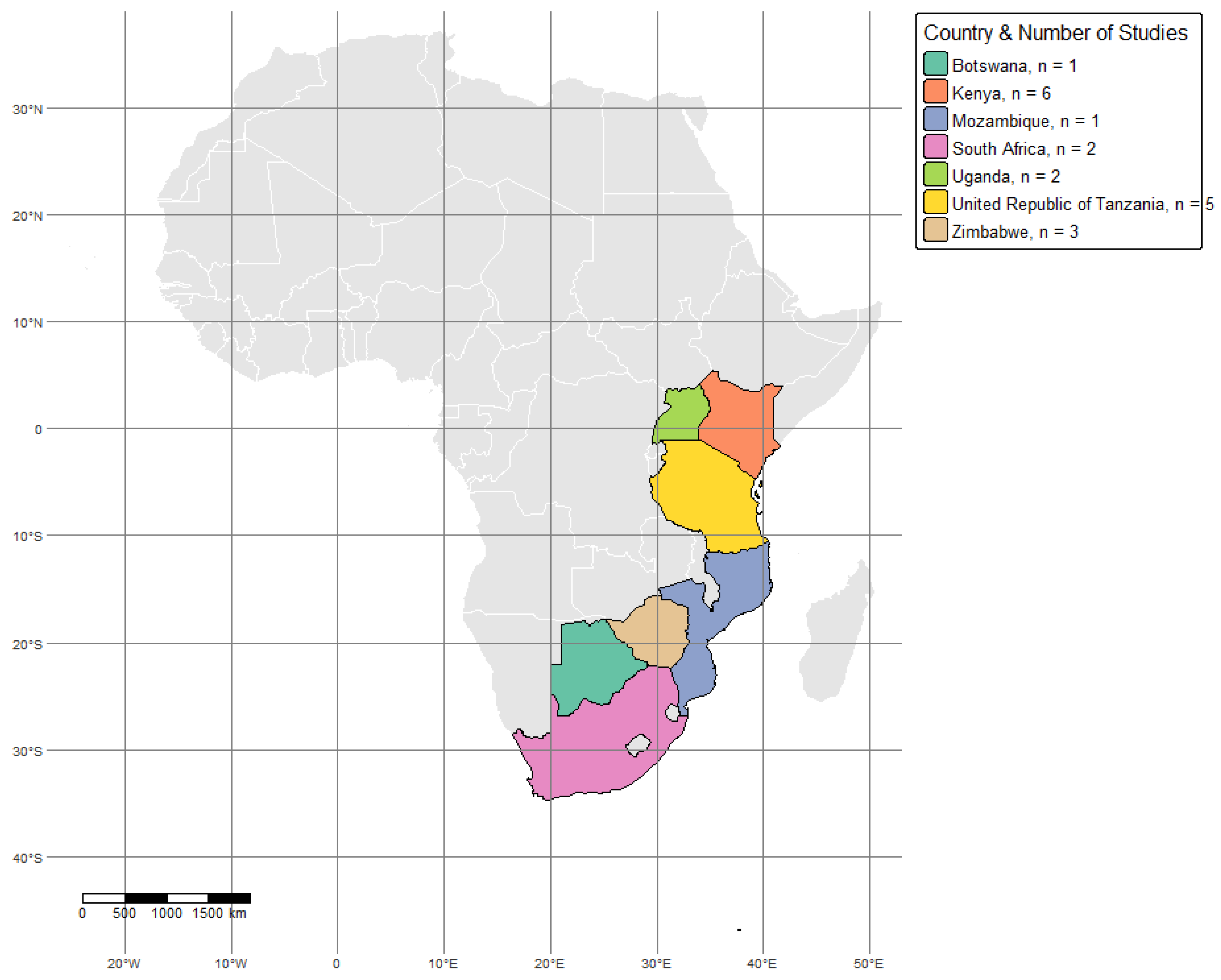
2.1. Study Area
2.2. Search Strategy
2.3. Screening Process
2.4. Data Extraction and Quality Assessment
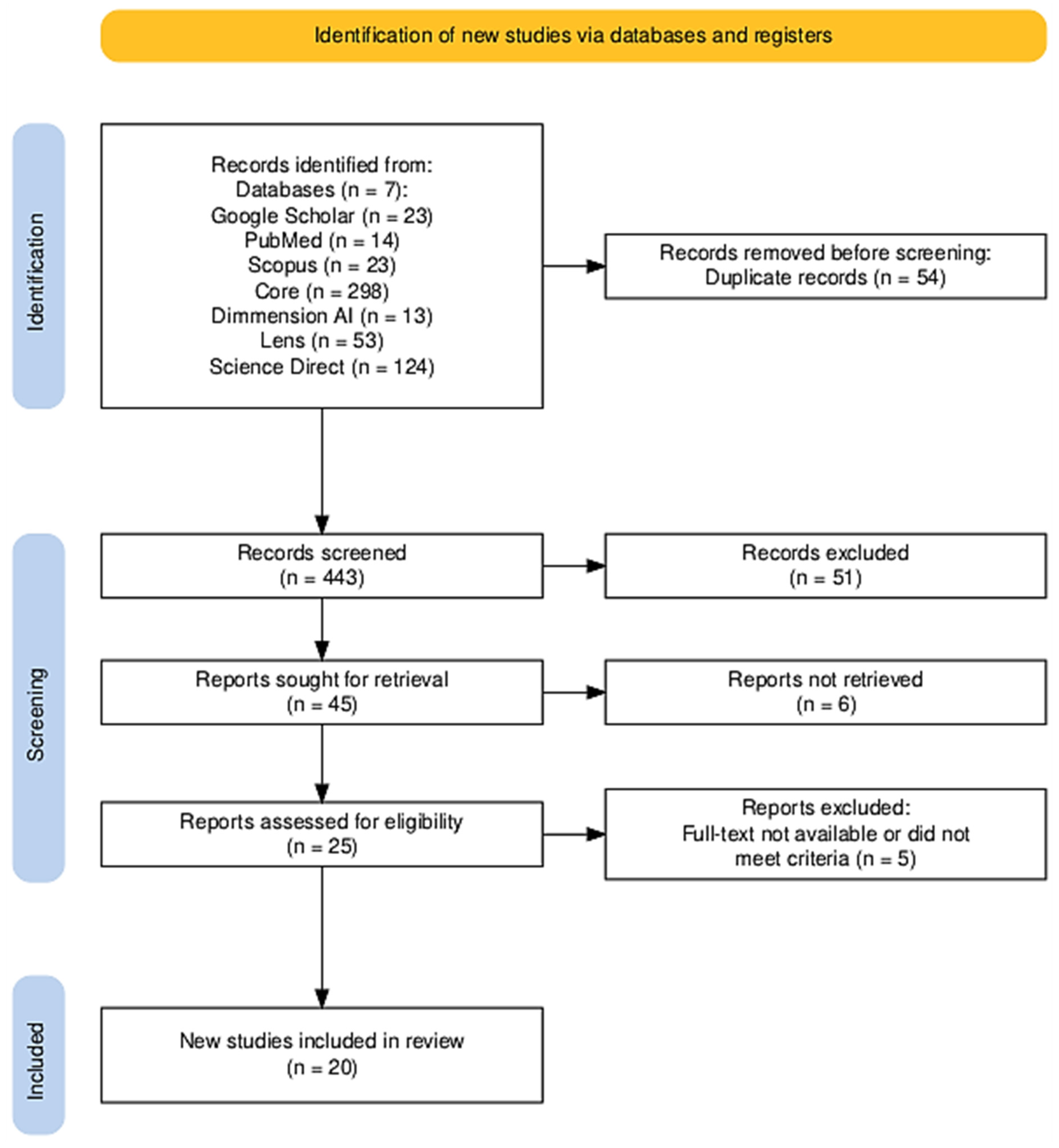
2.5. Study Inclusion and PRISMA Flow Diagram
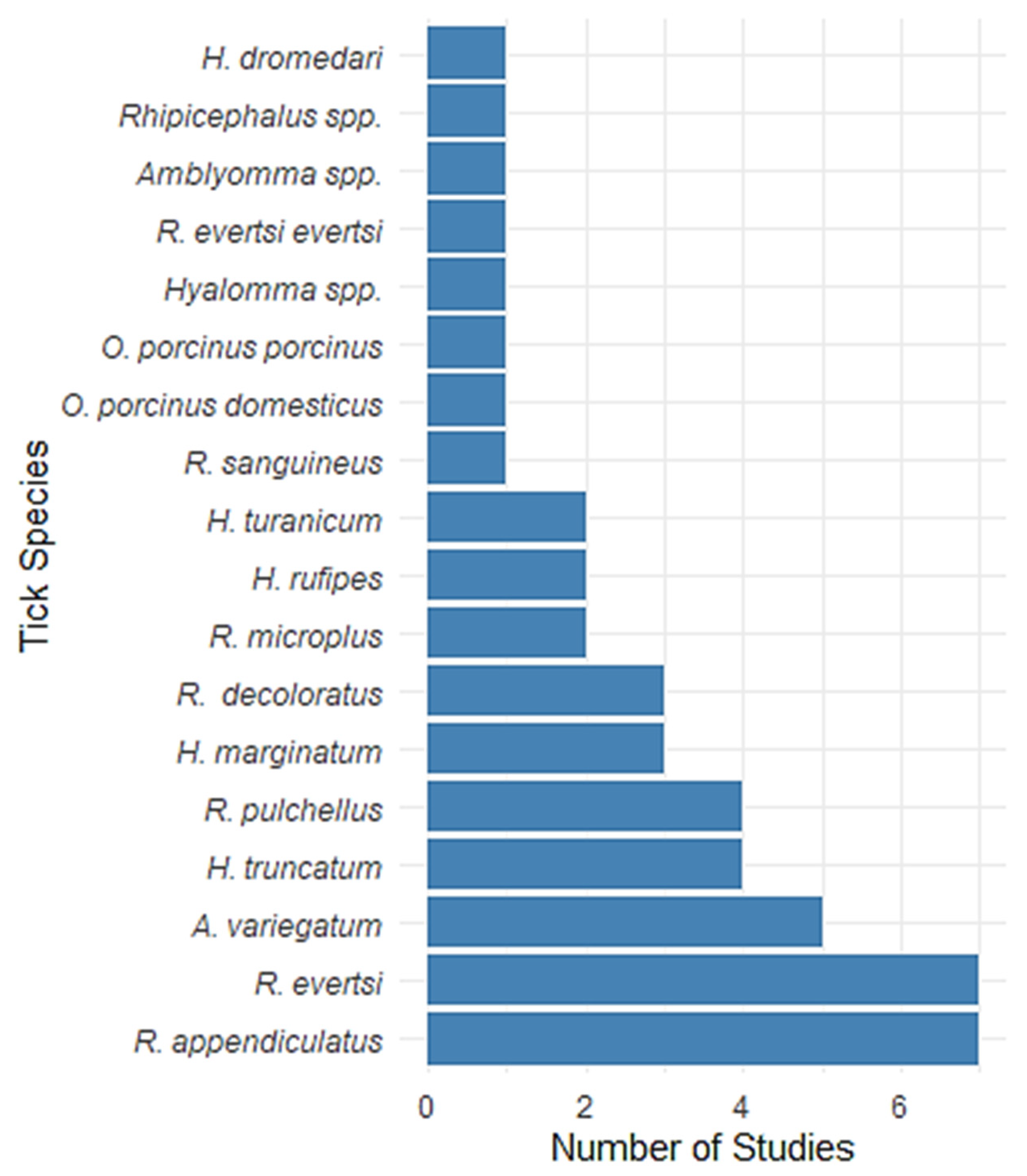
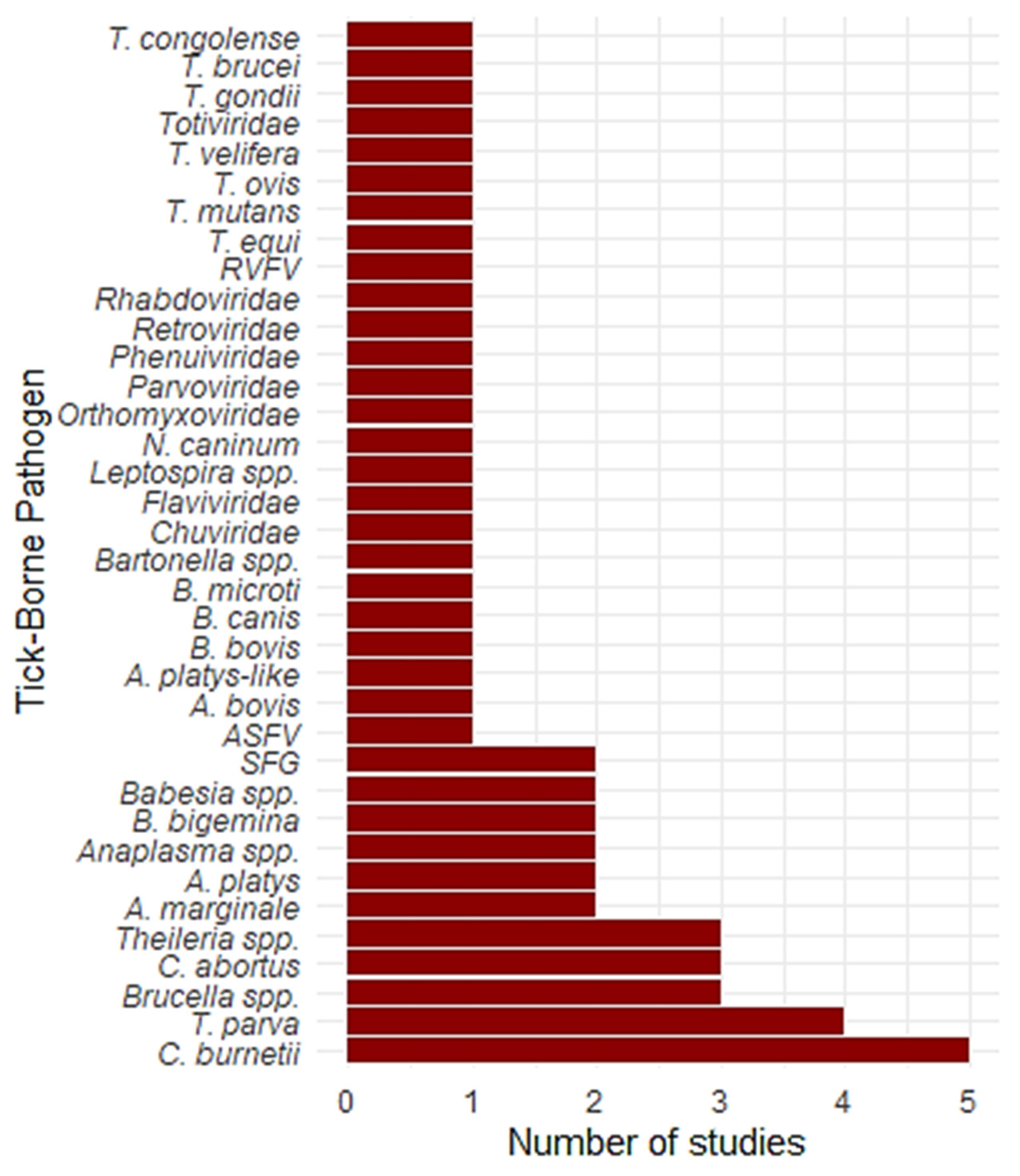
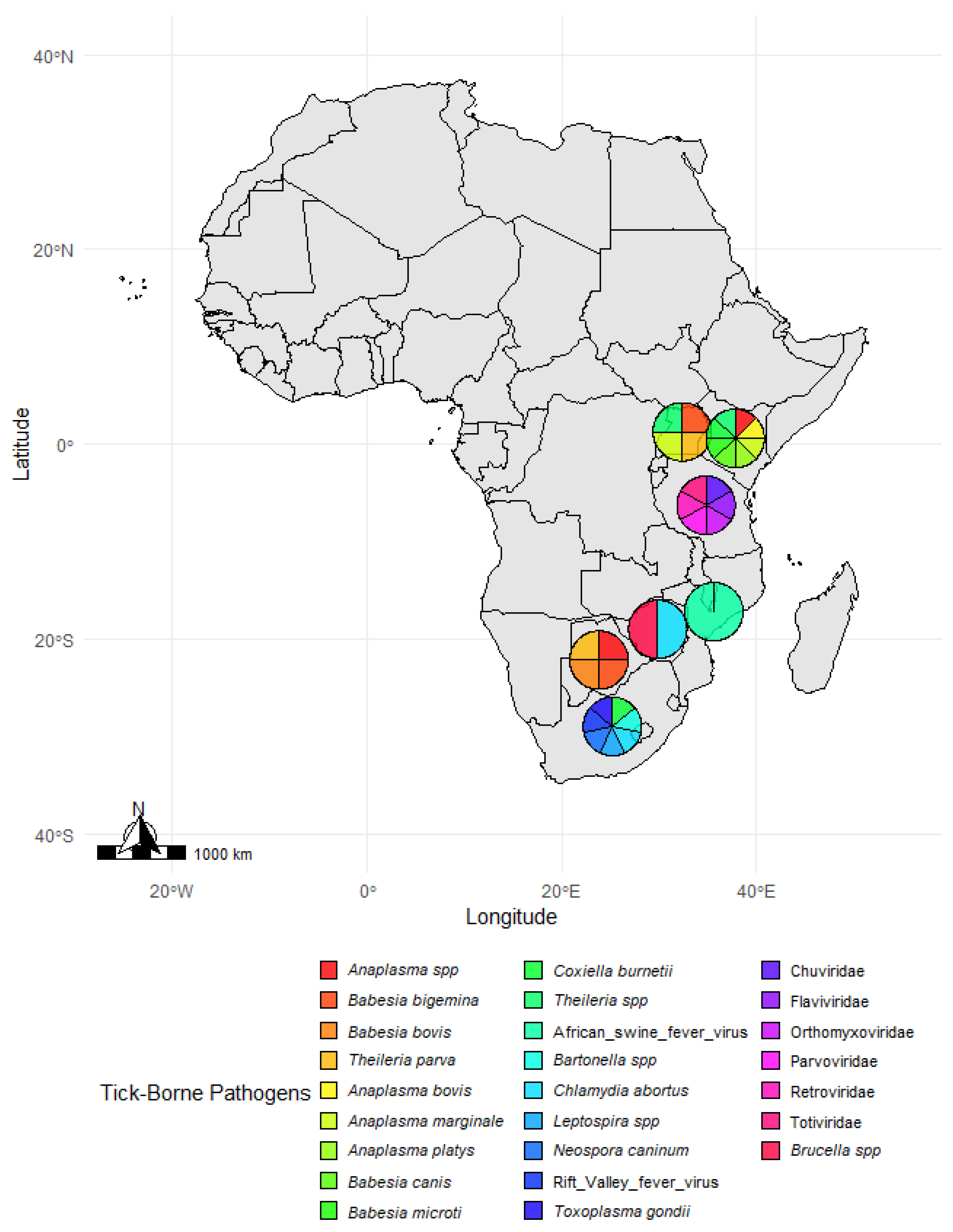
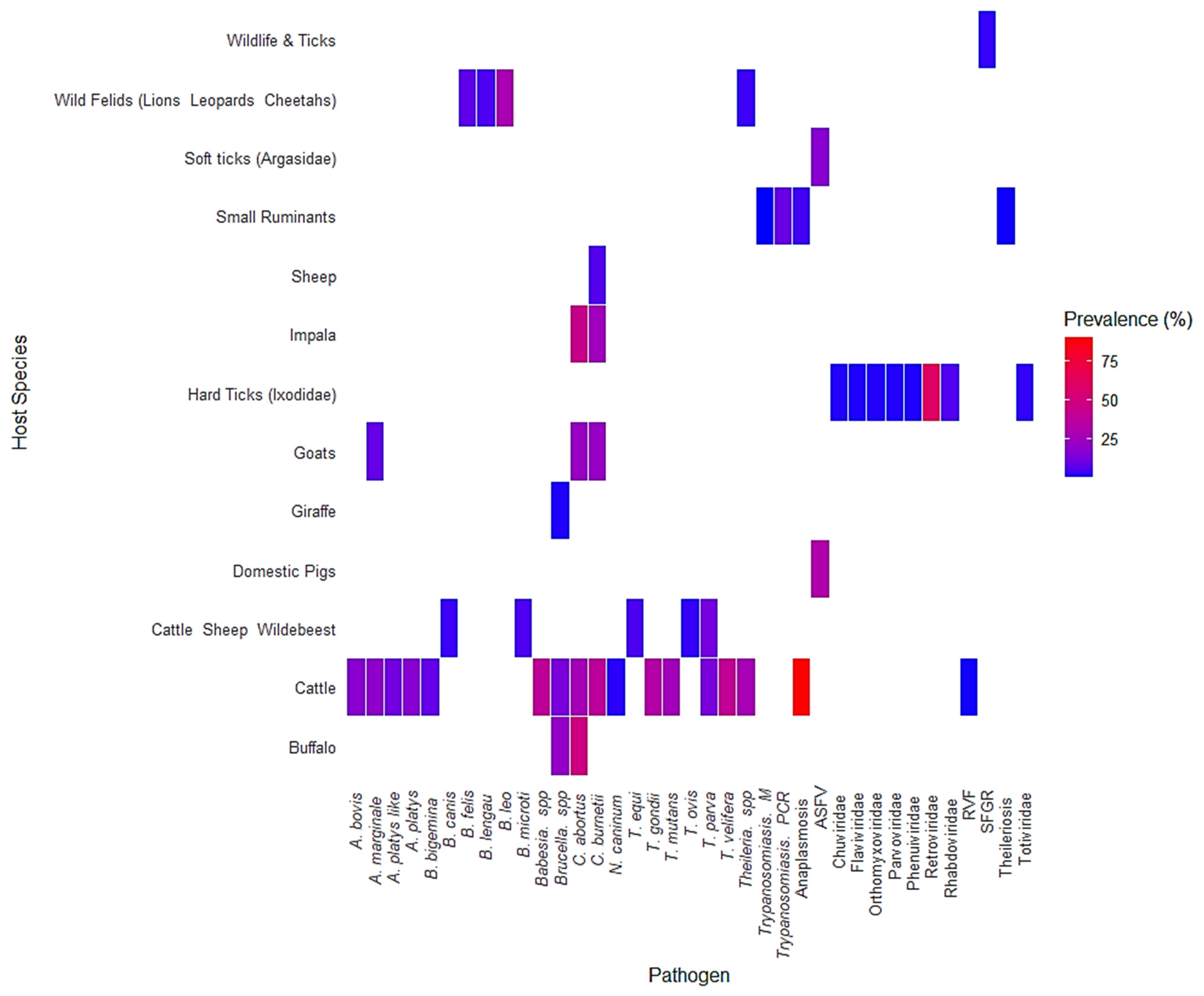
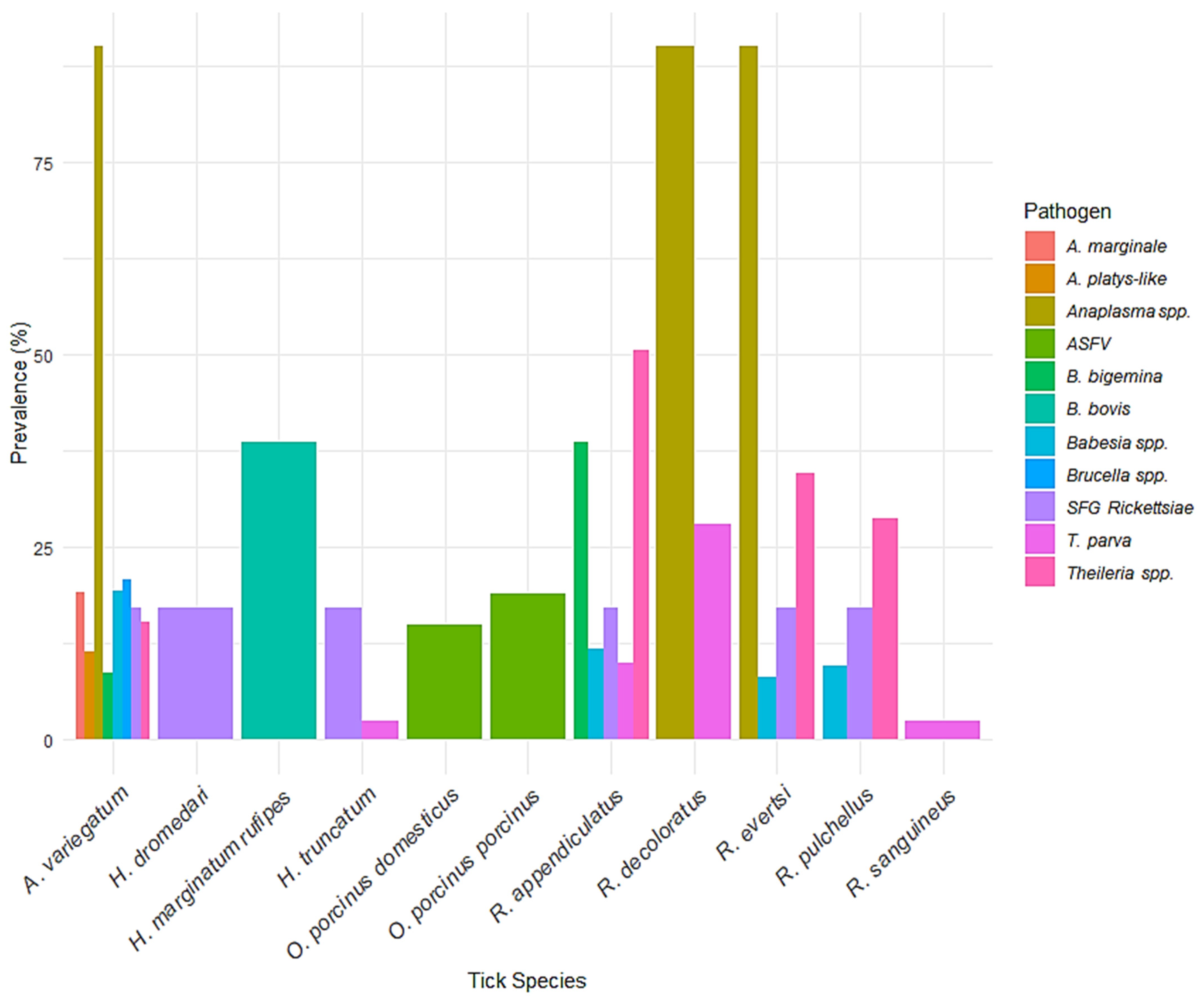
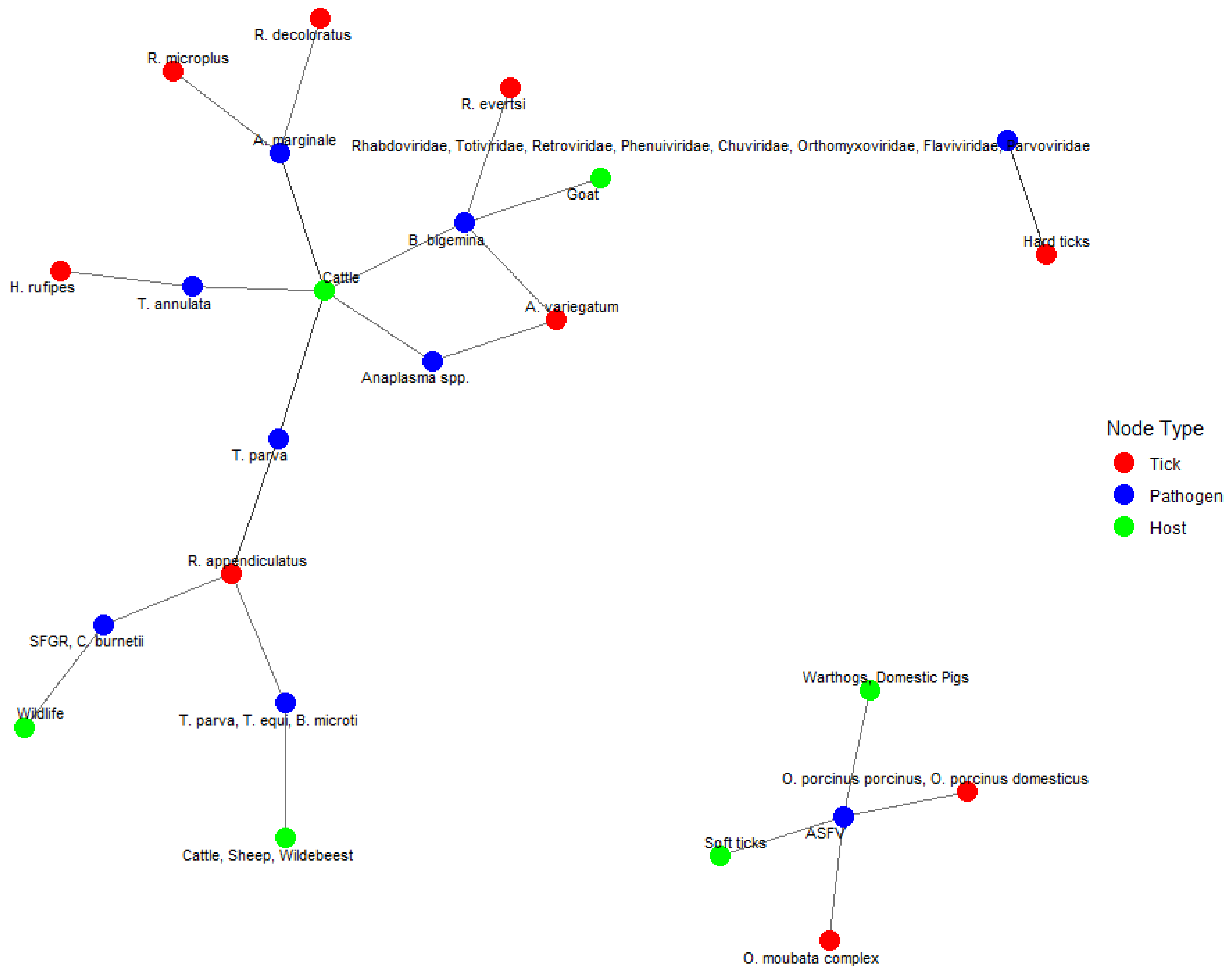
3. Results
4. Discussion
4.1. Prevalence and Distribution of Tick Species
4.2. Pathogen Prevalence and Host Interactions
4.3. Implications for Disease Control
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Topic | No. | Item | Location Where Item Is Reported |
|---|---|---|---|
| TITLE | |||
| Title | 1 | Identify the report as a systematic review. | Title: “A Systematic Review” is included |
| ABSTRACT | |||
| Abstract | 2 | See the PRISMA 2020 for Abstracts checklist | |
| INTRODUCTION | |||
| Rationale | 3 | Describe the rationale for the review in the context of existing knowledge. | Introduction: Provides background and justification |
| Objectives | 4 | Provide an explicit statement of the objective(s) or question(s) the review addresses. | Introduction: Clearly states the research question |
| METHODS | |||
| Eligibility criteria | 5 | Specify the inclusion and exclusion criteria for the review and how studies were grouped for the syntheses. | Section 2 |
| Information sources | 6 | Specify all databases, registers, websites, organisations, reference lists and other sources searched or consulted to identify studies. Specify the date when each source was last searched or consulted. | Section 2.3 |
| Search strategy | 7 | Present the full search strategies for all databases, registers, and websites, including any filters and limits used. | Section 2.2 |
| Selection process | 8 | Specify the methods used to decide whether a study met the inclusion criteria of the review, including how many reviewers screened each record and each report retrieved, whether they worked independently, and if applicable, details of automation tools used in the process. | Table 1—Summary of database search strategies |
| Data collection process | 9 | Specify the methods used to collect data from reports, including how many reviewers collected data from each report, whether they worked independently, any processes for obtaining or confirming data from study investigators, and if applicable, details of automation tools used in the process. | Section 2.4 |
| Data items | 10a | List and define all outcomes for which data were sought. Specify whether all results that were compatible with each outcome domain in each study were sought (e.g., for all measures, time points, analyses), and if not, the methods used to decide which results to collect. | Section 2.4 |
| 10b | List and define all other variables for which data were sought (e.g., participant and intervention characteristics, funding sources). Describe any assumptions made about any missing or unclear information. | Section 2.4 | |
| Study risk of bias assessment | 11 | Specify the methods used to assess risk of bias in the included studies, including details of the tool(s) used, how many reviewers assessed each study and whether they worked independently, and if applicable, details of automation tools used in the process. | Risk of Bias Assessment section |
| Effect measures | 12 | Specify for each outcome the effect measure(s) (e.g., risk ratio, mean difference) used in the synthesis or presentation of results. | Section 3 |
| Synthesis methods | 13a | Describe the processes used to decide which studies were eligible for each synthesis (e.g., tabulating the study intervention characteristics and comparing against the planned groups for each synthesis (item 5)). | Synthesis Methods section, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8, Figure 9 and Figure 10 |
| 13b | Describe any methods required to prepare the data for presentation or synthesis, such as handling of missing summary statistics, or data conversions. | Synthesis Methods section | |
| 13c | Describe any methods used to tabulate or visually display results of individual studies and syntheses. | Tables and Figures | |
| 13d | Describe any methods used to synthesize results and provide a rationale for the choice(s). If meta-analysis was performed, describe the model(s), method(s) to identify the presence and extent of statistical heterogeneity, and software package(s) used. | Synthesis Methods section, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8, Figure 9 and Figure 10 | |
| 13e | Describe any methods used to explore possible causes of heterogeneity among study results (e.g., subgroup analysis, meta-regression). | Section 3 | |
| 13f | Describe any sensitivity analyses conducted to assess robustness of the synthesized results. | Section 3 | |
| Reporting bias assessment | 14 | Describe any methods used to assess risk of bias due to missing results in a synthesis (arising from reporting biases). | Risk of Bias Assessment section |
| Certainty assessment | 15 | Describe any methods used to assess certainty (or confidence) in the body of evidence for an outcome. | Risk of Bias Assessment section |
| RESULTS | |||
| Study selection | 16a | Describe the results of the search and selection process, from the number of records identified in the search to the number of studies included in the review, ideally using a flow diagram. | PRISMA Flow Diagram (Figure 2) |
| 16b | Cite studies that might appear to meet the inclusion criteria, but which were excluded, and explain why they were excluded. | Section 2.5 | |
| Study characteristics | 17 | Cite each included study and present its characteristics. | Table 2—Characteristics of included studies |
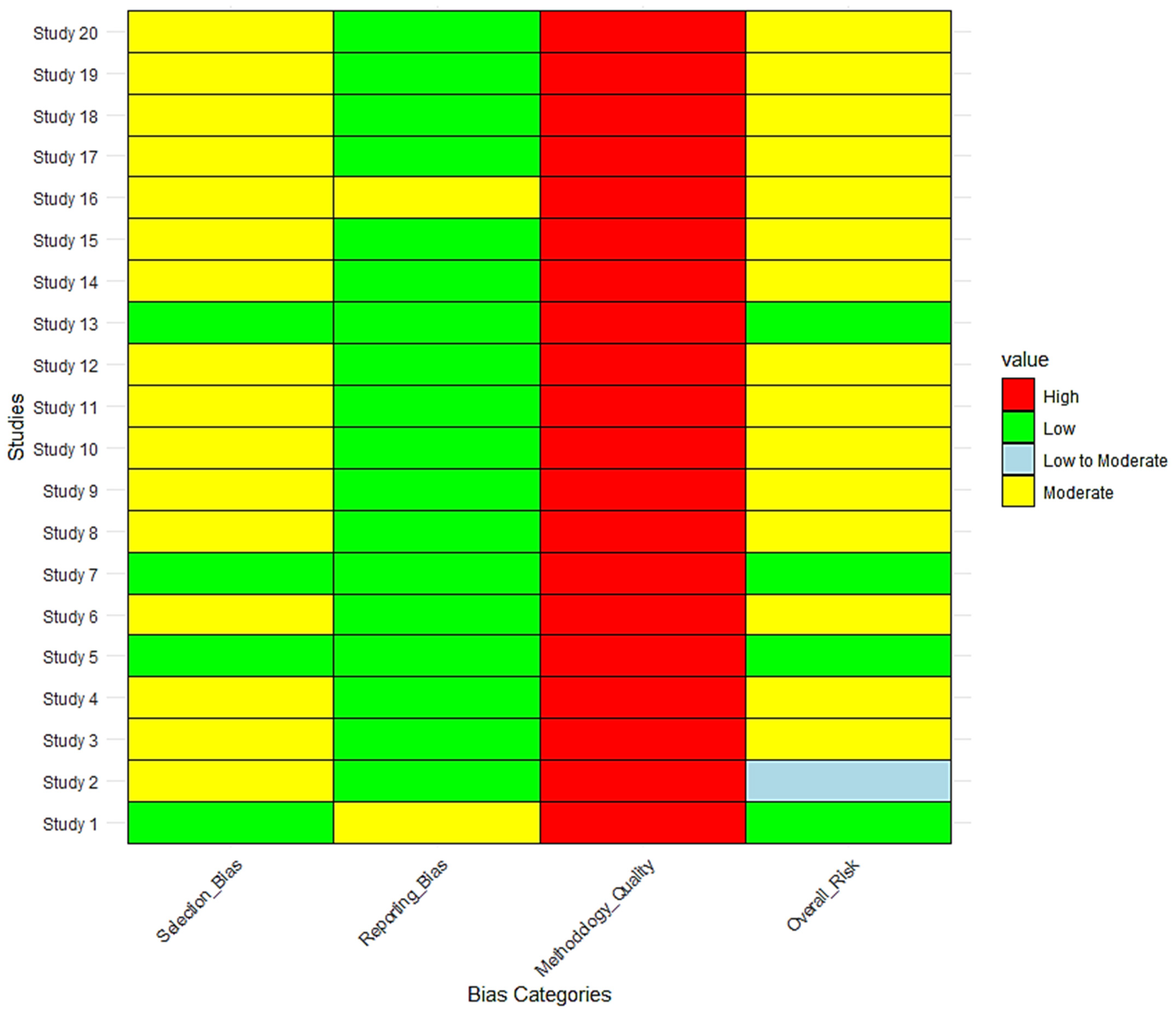
| Risk of bias in studies | 18 | Present assessments of risk of bias for each included study. | Risk of Bias Assessment section (Figure 11) |
| Results of individual studies | 19 | For all outcomes, present, for each study: (a) summary statistics for each group (where appropriate) and (b) an effect estimate and its precision (e.g., confidence/credible interval), ideally using structured tables or plots. | Tables and Figures (e.g., prevalence per tick species) |
| Results of syntheses | 20a | For each synthesis, briefly summarise the characteristics and risk of bias among contributing studies. | Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8, Figure 9 and Figure 10 |
| 20b | Present results of all statistical syntheses conducted. If meta-analysis was conducted, present for each the summary estimate and its precision (e.g., confidence/credible interval) and measures of statistical heterogeneity. If comparing groups, describe the direction of the effect. | Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8, Figure 9 and Figure 10 | |
| 20c | Present results of all investigations of possible causes of heterogeneity among study results. | Section 3 | |
| 20d | Present results of all sensitivity analyses conducted to assess the robustness of the synthesized results. | Section 3 | |
| Reporting biases | 21 | Present assessments of risk of bias due to missing results (arising from reporting biases) for each synthesis assessed. | Risk of Bias section |
| Certainty of evidence | 22 | Present assessments of certainty (or confidence) in the body of evidence for each outcome assessed. | Risk of Bias section |
| DISCUSSION | |||
| Discussion | 23a | Provide a general interpretation of the results in the context of other evidence. | Section 4 |
| 23b | Discuss any limitations of the evidence included in the review. | Discussion—Regional and Surveillance Gaps | |
| 23c | Discuss any limitations of the review processes used. | Discussion—Screening and Selection Process Limitations | |
| 23d | Discuss implications of the results for practice, policy, and future research. | Discussion—Future Directions and One Health Approach | |
| OTHER INFORMATION | |||
| Registration and protocol | 24a | Provide registration information for the review, including register name and registration number, or state that the review was not registered. | The review was not registered. |
| 24b | Indicate where the review protocol can be accessed, or state that a protocol was not prepared. | A protocol was not prepared. | |
| 24c | Describe and explain any amendments to information provided at registration or in the protocol. | Not applicable | |
| Support | 25 | Describe sources of financial or non-financial support for the review, and the role of the funders or sponsors in the review. | Not applicable |
| Competing interests | 26 | Declare any competing interests of review authors. | Conflicts of Interest section |
| Availability of data, code and other materials | 27 | Report which of the following are publicly available and where they can be found: template data collection forms; data extracted from included studies; data used for all analyses; analytic code; any other materials used in the review. | Data Availability Statement |
Appendix B
| Topic | No. | Item | Reported? |
|---|---|---|---|
| TITLE | |||
| Title | 1 | Identify the report as a systematic review. | Yes |
| BACKGROUND | |||
| Objectives | 2 | Provide an explicit statement of the main objective(s) or question(s) the review addresses. | Yes |
| METHODS | |||
| Eligibility criteria | 3 | Specify the inclusion and exclusion criteria for the review. | Yes |
| Information sources | 4 | Specify the information sources (e.g., databases, registers) used to identify studies and the date when each was last searched. | Yes |
| Risk of bias | 5 | Specify the methods used to assess risk of bias in the included studies. | Yes |
| Synthesis of results | 6 | Specify the methods used to present and synthesize results. | Yes |
| RESULTS | |||
| Included studies | 7 | Give the total number of included studies and participants and summarise relevant characteristics of studies. | Yes |
| Synthesis of results | 8 | Present results for main outcomes, preferably indicating the number of included studies and participants for each. If meta-analysis was conducted, report the summary estimate and confidence/credible interval. If comparing groups, indicate the direction of the effect (i.e., which group is favoured). | Yes |
| DISCUSSION | |||
| Limitations of evidence | 9 | Provide a brief summary of the limitations of the evidence included in the review (e.g., study risk of bias, inconsistency and imprecision). | Yes |
| Interpretation | 10 | Provide a general interpretation of the results and important implications. | Yes |
| OTHER | |||
| Funding | 11 | Specify the primary source of funding for the review. | Yes |
| Registration | 12 | Provide the register name and registration number. | No |
References
- Brites-Neto, J.; Duarte, K.M.; Martins, T.F. Tick-borne infections in human and animal population worldwide. Vet. World 2015, 8, 301–315. [Google Scholar] [CrossRef] [PubMed]
- Ledwaba, M.B.; Nozipho, K.; Tembe, D.; Onyiche, T.E.; Chaisi, M.E. Distribution and prevalence of ticks and tick-borne pathogens of wild animals in South Africa: A systematic review. Curr. Res. Parasitol. Vector-Borne Dis. 2022, 2, 100088. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Wang, H.; Wang, T.; Sun, W.; Yang, X.; Liu, J. Tick-borne pathogens and the vector potential of ticks in China. Parasites Vectors 2015, 8, 24. [Google Scholar] [CrossRef]
- Makwarela, T.G.; Seoraj-Pillai, N.; Nangammbi, T.C. Tick Control Strategies: Critical Insights into Chemical, Biological, Physical, and Integrated Approaches for Effective Hard Tick Management. Vet. Sci. 2025, 12, 114. [Google Scholar] [CrossRef] [PubMed]
- Rochlin, I.; Toledo, A. Emerging tick-borne pathogens of public health importance: A mini-review. J. Med. Microbiol. 2020, 69, 781–791. [Google Scholar] [CrossRef]
- Godde, C.M.; Mason-D’Croz, D.; Mayberry, D.E.; Thornton, P.K.; Herrero, M. Impacts of climate change on the livestock food supply chain; a review of the evidence. Glob. Food Sec. 2021, 28, 100488. [Google Scholar] [CrossRef]
- Ryan, J. Important diseases in sheep in South Africa. Farmer’s Wkly. 2022, 2022, 40–43. [Google Scholar]
- Van Helden, L.; Van Helden, P.D.; Meiring, C. Pathogens of Vertebrate Animals as Invasive Species: Insights from South Africa. In Biological Invasions in South Africa; Van Wilgen, B.W., Measey, J., Richardson, D.M., Wilson, J.R., Zengeya, T.A., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 249–274. [Google Scholar]
- Hassell, J.M.; Begon, M.; Ward, M.J.; Fèvre, E.M. Urbanization and Disease Emergence: Dynamics at the Wildlife–Livestock–Human Interface. Trends Ecol. Evol. 2017, 32, 55–67. [Google Scholar] [CrossRef]
- Katswara, T.; Mukaratirwa, S. Knowledge, Attitudes and Practices on African Tick Bite Fever of Rural Livestock Communities Living in a Livestock-Wildlife Interface Area in the Eastern Cape Province of South Africa. BMC Infect. Dis. 2021, 21, 497. [Google Scholar] [CrossRef]
- Liyanaarachchi, D.R.; Rajakaruna, R.S.; Dikkumbura, A.W.; Rajapakse, R.P. Ticks infesting wild and domestic animals and humans of Sri Lanka with new host records. Acta Trop. 2015, 142, 64–70. [Google Scholar] [CrossRef]
- Oundo, J.W.; Villinger, J.; Jeneby, M.; Ong’amo, G.; Otiende, M.; Makhulu, E.E.; Musa, A.A.; Ouso, D.O.; Wambua, L. Pathogens, Endosymbionts, and Blood-Meal Sources of Host-Seeking Ticks in the Fast-Changing Maasai Mara Wildlife Ecosystem. PLoS ONE 2020, 15, e0228366. [Google Scholar] [CrossRef] [PubMed]
- Raboloko, O.O.; Ramabu, S.S.; Guerrini, L.; Jori, F. Seroprevalence of Selected Tick Borne Pathogens and Diversity and Abundance of Ixodid Ticks (Acari: Ixodidae) at the Wildlife-Livestock Interface in Northern Botswana. Front. Vet. Sci. 2020, 7, 187. [Google Scholar] [CrossRef] [PubMed]
- Alale, T.Y.; Sormunen, J.J.; Nzeh, J.; Agjei, R.O.; Vesterinen, E.J.; Klemola, T. Public knowledge and awareness of tick-borne pathogens and diseases: A cross-sectional study in Ghana. Curr. Res. Parasitol. Vector-Borne Dis. 2024, 6, 100228. [Google Scholar] [CrossRef] [PubMed]
- Kimaro, E. Cattle Vector-Borne Disease Occurrence and Management and Climate Change Experiences in Pastoral Communities in Northern Tanzania. Ph.D. Thesis, The University of Sydney, Sydney, Australia, 2018. [Google Scholar]
- Byamukama, B.; Vudriko, P.; Tumwebaze, M.A.; Tayebwa, D.S.; Byaruhanga, J.; Angwe, M.K.; Li, J.; Galon, E.M.; Ringo, A.; Liu, M. Molecular detection of selected tick-borne pathogens infecting cattle at the wildlife–livestock interface of Queen Elizabeth National Park in Kasese District, Uganda. Ticks Tick-Borne Dis. 2021, 12, 101772. [Google Scholar] [CrossRef]
- Damian, D.; Maghembe, R.; Damas, M.; Wensman, J.J.; Berg, M. Application of viral metagenomics for study of emerging and reemerging tick-borne viruses. Vector-Borne Zoonotic Dis. 2020, 20, 557–565. [Google Scholar] [CrossRef]
- Damian, D. Navigating the Landscape of Tick Diversity: Integrating Molecular Approaches for Enhanced Control Measures. Vector-Borne Zoonotic Dis. 2024, 25, 1–13. [Google Scholar] [CrossRef]
- Labuda, M.; Nuttall, P. Tick-borne viruses. Parasitology 2004, 129 (Suppl. S1), S221–S245. [Google Scholar] [CrossRef]
- Maholbo-Shwabede, S.I.C.; Zishiri, O.T.; Thekisoe, O.; Makalo, M.J. Molecular Detection of Coxiella burnetii, Rickettsia africae and Anaplasma Species in Ticks From Domestic Animals in Lesotho. Pathogens 2021, 10, 1186. [Google Scholar] [CrossRef]
- Cravino, A.; Perelló, A.; Brazeiro, A. Livestock-wildlife interactions: Key aspects for reconnecting animal production and wildlife conservation. Anim. Front. 2024, 14, 13–19. [Google Scholar] [CrossRef]
- Adesiyun, A.A.; Knobel, D.L.; Thompson, P.N.; Wentzel, J.; Kolo, F.B.; Kolo, A.O.; Conan, A.; Simpson, G.J. Sero-epidemiological study of selected zoonotic and abortifacient pathogens in cattle at a wildlife-livestock interface in South Africa. Vector-Borne Zoonotic Dis. 2020, 20, 258–267. [Google Scholar] [CrossRef]
- Kasozi, K.I.; Namayanja, M.; Gaithuma, A.K.; Mahero, M.; Matovu, E.; Yamagishi, J.; Sugimoto, C.; MacLeod, E. Prevalence of hemoprotozoan parasites in small ruminants along a human-livestock-wildlife interface in western Uganda. Vet. Parasitol. Reg. Stud. Rep. 2019, 17, 100309. [Google Scholar] [CrossRef] [PubMed]
- Monakale, K.S.; Ledwaba, M.B.; Smith, R.M.; Gaorekwe, R.M.; Malatji, D.P. A systematic review of ticks and tick-borne pathogens of cattle reared by smallholder farmers in South Africa. Curr. Res. Parasitol. Vector-Borne Dis. 2024, 6, 100205. [Google Scholar] [CrossRef]
- Thompson, B.E. Occurrence of Theileria parva Infection in Cattle on a Farm in KwaZulu-Natal, South Africa. Master’s Thesis, University of Pretoria, Pretoria, South Africa, 2007. [Google Scholar]
- Magadu, R.; Thompson, P.N. Seroprevalence and factors associated with Coxiella burnetii exposure in goats in Moretele. Onderstepoort J. Vet. Res. 2023, 90, 2071. [Google Scholar] [CrossRef] [PubMed]
- Eygelaar, D.; Jori, F.; Mokopasetso, M.; Sibeko, K.P.; Collins, N.E.; Vorster, I.; Troskie, M.; Oosthuizen, M.C. Tick-Borne Haemoparasites in African Buffalo (Syncerus caffer) From Two Wildlife Areas in Northern Botswana. Parasites Vectors 2015, 8, 26. [Google Scholar] [CrossRef]
- Allan, F.K.; Sindoya, E.; Adam, K.E.; Byamungu, M.; Lea, R.S.; Lord, J.S.; Mbata, G.; Paxton, E.; Mramba, F.; Torr, S.J.; et al. A cross-sectional survey to establish Theileria parva prevalence and vector control at the wildlife-livestock interface, Northern Tanzania. Prev. Vet. Med. 2021, 196, 105491. [Google Scholar] [CrossRef]
- Surve, A.A.; Hwang, J.Y.; Manian, S.; Onono, J.O.; Yoder, J. Economics of East Coast fever: A literature review. Front. Vet. Sci. 2023, 10, 1239110. [Google Scholar] [CrossRef] [PubMed]
- Hayatou, H. Prevalence of Tick Infestations and Tick-Borne Diseases in Cattle in Cameroon. Open J. Anim. Sci. 2023, 13, 560–573. [Google Scholar] [CrossRef]
- Springer, A.; Shuaib, Y.A.; Isaa, M.H.; Ezz-Eldin, M.I.; Osman, A.Y.; Yagoub, I.A.; Abdalla, M.A.; Bakiet, A.O.; Mohmed-Noor, S.E.; Schaper, S.; et al. Tick Fauna and Associated Rickettsia, Theileria, and Babesia spp. in Domestic Animals in Sudan (North Kordofan and Kassala States). Microorganisms 2020, 8, 1969. [Google Scholar] [CrossRef]
- Kolar, J. Diagnosis and control of brucellosis in small ruminants. Prev. Vet. Med. 1984, 2, 215–225. [Google Scholar] [CrossRef]
- Ndeereh, D.R. Molecular Epidemiology of Spotted Fever Group Rickettsioses and Q Fever at the Wildlife-Livestock Interface in Maasai Mara and Laikipia Ecosystems, Kenya. Ph.D. Thesis, University of Nairobi, Nairobi, Kenya, 2016. [Google Scholar]
- Matope, G.; Tivapasi, M.; Ndengu, M.; Pfukenyi, D.M.; De Garine-Wichatitsky, M.; Bhandi, S.; Bonfini, B.; Murondoti, A.; Scacchia, M. Brucellosis and chlamydiosis seroprevalence in goats at livestock–wildlife interface areas of Zimbabwe. Onderstepoort J. Vet. Res. 2019, 86, a1670. [Google Scholar] [CrossRef]
- Chepkwony, R.; Van Bommel, S.; Van Langevelde, F. Interactive effects of biological, human and environmental factors on tick loads in Boran cattle in tropical drylands. Parasites Vectors 2021, 14, 188. [Google Scholar] [CrossRef] [PubMed]
- Ramzan, M.; Iqbal, S.; Kousar, I.; Shahid, M.; Hussain, T.; Nadeem, M.; Sajid, M.Y.; Iram, F. Identification of tick species on domesticated cattle in Pakistan. Egypt. Acad. J. Biol. Sci. E Med. Entomol. Parasitol. 2021, 13, 57–61. [Google Scholar] [CrossRef]
- Walker, A.R. Eradication and control of livestock ticks: Biological, economic and social perspectives. Parasitology 2011, 138, 945–959. [Google Scholar] [CrossRef]
- Mwamuye, M.M.; Odongo, D.; Kazungu, Y.; Kindoro, F.; Gwakisa, P.; Bishop, R.P.; Nijhof, A.M.; Obara, I. Variant analysis of the sporozoite surface antigen gene reveals that asymptomatic cattle from wildlife-livestock interface areas in northern Tanzania harbour buffalo-derived T. parva. Parasitol. Res. 2020, 119, 3817–3828. [Google Scholar] [CrossRef]
- Nguro, B. Occurrence of Q Fever at the Wildlife-Livestock Interface of Amboseli Ecosystem, Kenya. Master’s Thesis, University of Nairobi, Nairobi, Kenya, 2017. [Google Scholar]
- Ndengu, M.; Matope, G.; Tivapasi, M.; Scacchia, M.; Bonfini, B.; Pfukenyi, D.M.; de Garine-Wichatitsky, M. Sero-prevalence of chlamydiosis in cattle and selected wildlife species at a wildlife/livestock interface area of Zimbabwe. Trop. Anim. Health Prod. 2018, 50, 1107–1117. [Google Scholar] [CrossRef] [PubMed]
- Haddaway, N.R.; Page, M.J.; Pritchard, C.C.; McGuinness, L.A. PRISMA2020: An R package and Shiny app for producing PRISMA 2020-compliant flow diagrams, with interactivity for optimised digital transparency and Open Synthesis. Campbell Syst. Rev. 2022, 18, e1230. [Google Scholar] [CrossRef]
- Damian, D.; Damas, M.; Wensman, J.J.; Berg, M. Phylogeny and genetic relationship between hard ticks (Ixodidae) infesting cattle collected from selected areas of a wildlife-livestock interface ecosystem of Mikumi National Park, Tanzania. Res. Sq. 2020. [Google Scholar] [CrossRef]
- Damian, D.; Damas, M.; Wensman, J.J.; Berg, M. Diversity of viruses in hard ticks (Ixodidae) from select areas of a wildlife-livestock interface ecosystem at Mikumi National Park, Tanzania. Am. J. BioSci. 2020, 8, 150–157. [Google Scholar] [CrossRef]
- Okal, M.N.; Odhiambo, B.K.; Otieno, P.; Bargul, J.L.; Masiga, D.; Villinger, J.; Kalayou, S. Anaplasma and Theileria pathogens in cattle of Lambwe Valley, Kenya: A case for pro-active surveillance in the wildlife–livestock interface. Microorganisms 2020, 8, 1830. [Google Scholar] [CrossRef]
- Ndeereh, D.; Muchemi, G.; Thaiyah, A.; Otiende, M.; Angelone-Alasaad, S.; Jowers, M.J. Molecular survey of Coxiella burnetii in wildlife and ticks at wildlife–livestock interfaces in Kenya. Exp. Appl. Acarol. 2017, 72, 277–289. [Google Scholar] [CrossRef]
- Simpson, G.J.; Quan, V.; Frean, J.; Knobel, D.L.; Rossouw, J.; Weyer, J.; Marcotty, T.; Godfroid, J.; Blumberg, L.H. Prevalence of selected zoonotic diseases and risk factors at a human-wildlife-livestock interface in Mpumalanga Province, South Africa. Vector-Borne Zoonotic Dis. 2018, 18, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Quembo, C.J.; Jori, F.; Vosloo, W.; Heath, L. Genetic characterization of African swine fever virus isolates from soft ticks at the wildlife/domestic interface in Mozambique and identification of a novel genotype. Transbound. Emerg. Dis. 2018, 65, 420–431. [Google Scholar] [CrossRef]
- Damian, D.; Damas, M.; Wensman, J.J.; Berg, M. Molecular diversity of hard tick species from selected areas of a wildlife-livestock interface ecosystem at Mikumi National Park, Morogoro Region, Tanzania. Vet. Sci. 2021, 8, 36. [Google Scholar] [CrossRef]
- Mukolwe, S.A. Diversity of Ticks and Tick-Borne Protozoan Parasites from Livestock and Wildebeests at the Maasai-Mara Wildlife-Livestock Interface, Narok County, Kenya. Master’s Thesis, University of Nairobi, Nairobi, Kenya, 2018. [Google Scholar]
- Githaka, N.W.O. Molecular investigation of tick-borne protozoan parasites at the livestock-wildlife interface in Kenya and evaluation of a candidate anti-tick vaccine antigen. Jpn. J. Vet. Res. 2013, 61, 4. [Google Scholar]
- Gomo, C.; de Garine-Wichatitsky, M.; Caron, A.; Pfukenyi, D.M. Survey of brucellosis at the wildlife–livestock interface on the Zimbabwean side of the Great Limpopo Transfrontier Conservation Area. Trop. Anim. Health Prod. 2012, 44, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Koual, R.; Buysse, M.; Grillet, J.; Binetruy, F.; Ouass, S.; Sprong, H.; Duhayon, M.; Boulanger, N.; Jourdain, F.; Alafaci, A.; et al. Phylogenetic evidence for a clade of tick-associated trypanosomes. Parasites Vectors 2023, 16, 3. [Google Scholar] [CrossRef]
- Luu, L.; Bown, K.J.; Palomar, A.M.; Kazimírová, M.; Bell-Sakyi, L. Isolation and partial characterisation of a novel Trypanosoma from the tick Ixodes ricinus. Ticks Tick-Borne Dis. 2020, 11, 101501. [Google Scholar] [CrossRef]
- Krige, A.-S.; Thompson, R.C.; Clode, P. ‘Hang on a Tick’—Are Ticks Really the Vectors for Australian Trypanosomes? Trends Parasitol. 2019, 35, 596–606. [Google Scholar] [CrossRef] [PubMed]
- Horak, I.G.; Heyne, H.; Williams, R.; Gallivan, G.J.; Spickett, A.M.; Bezuidenhout, J.D.; Estrada-Peña, A. The Ixodid Ticks (Acari: Ixodidae) of Southern Africa; Springer: Cham, Switzerland, 2018. [Google Scholar]
- Nemaungwe, T.M.; van Dalen, E.; Waniwa, E.O.; Makaya, P.V.; Chikowore, G.; Chidawanyika, F. Biogeography of the theileriosis vector, Rhipicephalus appendiculatus under current and future climate scenarios of Zimbabwe. Exp. Appl. Acarol. 2023, 90, 67–82. [Google Scholar] [CrossRef]
- Makwarela, T.G.; Nyangiwe, N.; Masebe, T.M.; Djikeng, A.; Nesengani, L.T.; Smith, R.M.; Mapholi, N.O. Morphological and Molecular Characterization of Tick Species Infesting Cattle in South Africa. Vet. Sci. 2024, 11, 638. [Google Scholar] [CrossRef]
- Rhipicephalus Evertsi Evertsi. CABI Compendium. Available online: https://www.cabidigitallibrary.org/doi/full/10.1079/cabicompendium.66004 (accessed on 7 April 2025).
- Tarekegn, Z.S.; Leta, S.; Nigatu, S.D.; Mekonnen, S.A.; Molla, W. Modeling the spatial distribution and environmental factors of dominant ixodid tick species parasitizing cattle in northwest Ethiopia. Vet. Parasitol. 2025, 335, 110436. [Google Scholar] [CrossRef]
- Sungirai, M.; Madder, M.; Moyo, D.Z.; De Clercq, P.; Abatih, E.N. An update on the ecological distribution of the Ixodidae ticks in Zimbabwe. Exp. Appl. Acarol. 2015, 66, 269–280. [Google Scholar] [CrossRef]
- Wilson, M.L.; Dykstra, E.A.; Schmidt, B.A. Temperature- and humidity-dependent longevity of unfed adult Hyalomma truncatum (Acari: Ixodidae). J. Med. Entomol. 1993, 30, 467–471. [Google Scholar] [CrossRef] [PubMed]
- De Waal, D. The Transovarial Transmission of Babesia caballi by Hyalomma truncatum. Onderstepoort J. Vet. Res. 1990, 57, 99–100. [Google Scholar] [PubMed]
- Silatsa, B.A.; Simo, G.; Githaka, N.; Kamga, R.; Oumarou, F.; Keambou Tiambo, C.; Machuka, E.; Domelevo, J.-B.; Odongo, D.; Bishop, R.; et al. First detection of Theileria parva in cattle from Cameroon in the absence of the main tick vector Rhipicephalus appendiculatus. Transbound. Emerg. Dis. 2020, 67, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Maboko, B.B.; Sibeko-Matjila, K.P.; Pierneef, R.; Chan, W.Y.; Josemans, A.; Marumo, R.D.; Mbizeni, S.; Latif, A.A.; Mans, B.J. South African buffalo-derived Theileria parva is distinct from other buffalo and cattle-derived T. parva. Front. Genet. 2021, 12, 666096. [Google Scholar] [CrossRef]
- Booysen, S.; Horak, I.G.; Heyne, H.; Smit, W.J.; Halajian, A. Parasites of domestic and wild animals in South Africa. L. Ixodid ticks infesting horses and donkeys. Onderstepoort J. Vet. Res. 2017, 84, 1–6. [Google Scholar]
- Barré, N.; Garris, G.; Camus, E. Propagation of the tick Amblyomma variegatum in the Caribbean. Rev. Sci. Tech. 1995, 14, 841–855. [Google Scholar] [CrossRef]
- Mathison, B.A.; Gerth, W.J.; Pritt, B.S.; Baugh, S. Introduction of the exotic tick Hyalomma truncatum on a human with travel to Ethiopia: A case report. Ticks Tick-Borne Dis. 2015, 6, 152–154. [Google Scholar] [CrossRef]
- Mapholi, N.; Banga, C.; Dzama, K.; Matika, O.; Riggio, V.; Nyangiwe, N.; Maiwashe, N. Prevalence and tick loads in Nguni cattle reared in different environmental conditions across four provinces of South Africa. Vet. World 2022, 15, 1943–1953. [Google Scholar] [CrossRef]
- Wanzala, W. Distribution of Ticks and Tick-Borne Pathogens, Hosts, Habitat and Diseases in Kenya and Some Parts of Africa: A Mini Review. J. Anim. Res. Nutr. 2023, 8, 066. [Google Scholar]
- Gashaw, A.; Tessema, T. Prevalence of Ticks on Local and Crossbred Cattle in and Around Asella Town, Southeast Ethiopia. Ethiop. Vet. J. 2011, 14, 79–89. [Google Scholar] [CrossRef]
- Belete, W.; Mekuria, S. Prevalence, Species Identification, and Associated Risk Factor Ixodid Tick Infestation of Cattle in Selamago District, Southern Ethiopia. Environ. Health Insights 2023, 17, 11786302231211076. [Google Scholar] [CrossRef] [PubMed]
- Bhagoos, M.; Siame, T.; Munyeme, M.; Mwase, E.T. Comparative Analysis of Tick Burden, Diversity, and Distribution in Intensively Managed Wildlife Estates With and Without Prescribed Burning. GSC Adv. Res. Rev. 2023, 17, 169–180. [Google Scholar] [CrossRef]
- Jones, R.T.; Tytheridge, S.; Smith, S.J.; Levine, R.S.; Hodges, M.H.; Ansumana, R.; Wulff, S.; Whitworth, J.; Logan, J.G. The Threat of Vector-Borne Diseases in Sierra Leone. Am. J. Trop. Med. Hyg. 2023, 109, 10–21. [Google Scholar] [CrossRef]
- Theiler, A. The Immunisation of Cattle Against East Coast Fever; Cape Times Limited, Government Printers: Cape Town, South Africa, 1912. [Google Scholar]
- Nene, V.; Kiara, H.; Lacasta, A.; Pelle, R.; Svitek, N.; Steinaa, L. The biology of Theileria parva and control of East Coast fever—Current status and future trends. Ticks Tick-Borne Dis. 2016, 7, 549–564. [Google Scholar] [CrossRef]
- Ruybal, P.; Moretta, R.; Perez, A.; Petrigh, R.; Zimmer, P.; Alcaraz, E.; Echaide, I.; De Torioni Echaide, S.; Kocan, K.M.; De la Fuente, J.; et al. Genetic diversity of Anaplasma marginale in Argentina. Vet. Parasitol. 2009, 162, 176–180. [Google Scholar] [CrossRef]
- Espinaze, M.P.A.; Hellard, E.; Horak, I.G.; Cumming, G.S. Domestic mammals facilitate tick-borne pathogen transmission networks in South African wildlife. Biol. Conserv. 2018, 221, 228–236. [Google Scholar] [CrossRef]
- Allsopp, B.A. Natural history of Ehrlichia ruminantium. Vet. Parasitol. 2010, 167, 123–135. [Google Scholar] [CrossRef]
- Golnar, A.J.; Martin, E.; Wormington, J.D.; Kading, R.C.; Teel, P.D.; Hamer, S.A.; Hamer, G.L. Reviewing the potential vectors and hosts of African swine fever virus transmission in the United States. Vector-Borne Zoonotic Dis. 2019, 19, 512–524. [Google Scholar] [CrossRef]
- Bhandi, S. Seroprevalence and Risk Factors of Brucellosis, Chlamydiosis and Ehrlichiosis in Goats at Different Wildlife/Livestock Interface Areas in the South-Eastern Lowveld of Zimbabwe. Master’s Thesis, University of Zimbabwe, Harare, Zimbabwe, 2019. [Google Scholar]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef] [PubMed]


| Database | Search Count | Search String | Reason for Reduction | Imported Count |
|---|---|---|---|---|
| Google Scholar | 431 | TITLE: ((tick* OR Ixodidae OR “hard tick*”) AND “wildlife–livestock interface” AND (prevalence OR distribution OR “tick-borne disease”)) | Irrelevant, language, and outside Africa | 23 |
| PubMed/MEDLINE | 14 | ((tick* OR ticks OR Ixodidae OR “hard tick*”) AND “wildlife–livestock interface”) | None | 14 |
| Scopus | 24 | TITLE-ABS-KEY (ticks OR hard AND ticks AND wildlife–livestock AND interface) AND PUBYEAR > 2014 AND PUBYEAR < 2026 | Years | 23 |
| Core | 319 | (tick* OR Ixodidae OR “hard tick”) AND “wildlife–livestock interface” AND (prevalence OR distribution OR “tick-borne disease”) | Language and years | 298 |
| Dimension AI | 14 | ((tick* OR Ixodidae OR “hard tick”) AND “wildlife–livestock interface” AND (prevalence OR distribution OR “tick-borne disease”)), 2025 or 2023 or 2021 or 2020 or 2018 or 2017 or 2016, Article | None | 13 |
| Lens | 53 | Scholarly Works (53) = ((tick* OR (Ixodidae OR “hard tick”)) AND (“wildlife–livestock interface” AND (prevalence OR (distribution OR “tick-borne disease”)))) | None | 53 |
| Science Direct | 124 | tick AND wildlife AND livestock AND interface | None | 124 |
| Country/Region | Hosts | Sample Size | Total Pathogens | Pathogens with Prevalence | Ref |
|---|---|---|---|---|---|
| Queen Elizabeth NP, Uganda | Cattle | 208 | 6 |
| [16] |
| Serengeti NP, Tanzania | Cattle | 770 | 1 |
| [28] |
| Budongo Forest, Uganda | Goats (Caprines), Sheep (Ovines) | 712 (666 goats, 46 sheep) | 6 |
| [23] |
| Zimbabwe (south-eastern lowveld—Gonarezhou NP, Malilangwe Conservancy) | Goats | 563 | 2 |
| [34] |
| Mikumi NP, Tanzania | Cattle | 252 cattle, 630 ticks collected | 0 | N/A—The study identified tick species | [42] |
| South Africa (Mnisi community—bordering Kruger National Park, Mpumalanga) | Cattle | 184 | 5 |
| [22] |
| Tanzania (Mikumi National Park interface) | Ticks | 400 ticks | 17+ virus families (metagenomic detection) |
| [43] |
| Botswana (Northern—Maun West & Chobe West) | Cattle | 301 | 4 |
| [13] |
| Kenya (Lambwe Valley—Ruma National Park interface) | Cattle (zebu) | 680 | 6 |
| [44] |
| Kenya (Laikipia and Maasai Mara National Reserve) | Wildlife, ticks | Wildlife: 152 Ticks: 851 (Laikipia: 756; MMNR: 95) | 1 |
| [45] |
| Bushbuckridge, South Africa | Humans | 138 | 4 |
| [46] |
| Mozambique (Gorongosa National Park and buffer zone) | Soft ticks | 1865 soft ticks | 1 |
| [47] |
| Tanzania (Northern—Simanjiro and Monduli interface areas) | Cattle, Buffalo | Cattle: 160 Buffalo: 22 | 1 |
| [38] |
| Tanzania (Mikumi National Park interface, Morogoro Region) | Cattle, Goats, Environment | 436 animals (260 cattle, 176 goats) 847 ticks collected | 0 | Not assessed—tick diversity study | [48] |
| Zimbabwe | Cattle, African buffalo, kudu, impala | Cattle: 1011 Buffalo: 111 Kudu: 18 Impala: 32 | 1 |
| [40] |
| Kenya (Narok County—Maasai Mara wildlife–livestock interface) | Cattle, Sheep, Wildebeest | 61 animals (30 cattle, 30 wildebeest, 1 sheep); 165 ticks collected | 5 |
| [49] |
| Kenya (Laikipia and Maasai Mara ecosystems) | Wildlife; Ticks | 152 wildlife 851 ticks | 2 |
| [33] |
| Kenya (Various wildlife–livestock interface zones: Nairobi NP, Maasai Mara, Laikipia, Marula) | Lions, Leopards, Cheetahs, Giraffes, Waterbuck | 253 | 2 |
| [50] |
| Kenya (Amboseli ecosystem, Kajiado County) | Impalas, Sheep, Goats | Sheep: 200 Goats: 300 Impalas: 20 | 1 |
| [39] |
| Zimbabwe (Southeast lowveld—Gonarezhou and Kruger NP interface) | Cattle, Wildlife (buffalo, impala, kudu, giraffe) | Cattle: 1158 Wildlife: 97 | 1 |
| [51] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Makwarela, T.G.; Seoraj-Pillai, N.; Nangammbi, T.C. Distribution and Prevalence of Ticks and Tick-Borne Pathogens at the Wildlife-Livestock Interface in Africa: A Systematic Review. Vet. Sci. 2025, 12, 364. https://doi.org/10.3390/vetsci12040364
Makwarela TG, Seoraj-Pillai N, Nangammbi TC. Distribution and Prevalence of Ticks and Tick-Borne Pathogens at the Wildlife-Livestock Interface in Africa: A Systematic Review. Veterinary Sciences. 2025; 12(4):364. https://doi.org/10.3390/vetsci12040364
Chicago/Turabian StyleMakwarela, Tsireledzo Goodwill, Nimmi Seoraj-Pillai, and Tshifhiwa Constance Nangammbi. 2025. "Distribution and Prevalence of Ticks and Tick-Borne Pathogens at the Wildlife-Livestock Interface in Africa: A Systematic Review" Veterinary Sciences 12, no. 4: 364. https://doi.org/10.3390/vetsci12040364
APA StyleMakwarela, T. G., Seoraj-Pillai, N., & Nangammbi, T. C. (2025). Distribution and Prevalence of Ticks and Tick-Borne Pathogens at the Wildlife-Livestock Interface in Africa: A Systematic Review. Veterinary Sciences, 12(4), 364. https://doi.org/10.3390/vetsci12040364