1. Introduction
Soybean meal and fish meal are primary protein sources in poultry diets, with fish meal particularly valued for its well-balanced amino acid profile, high digestibility, and palatability. However, many feed manufacturer avoid using fish meal in poultry feed due to its higher variability in quality, potential risk of cross-contamination, and environmental concerns associated with the sustainability of the practice [
1]. Consequently, fish meal is generally replaced by soybean meal, despite its less balanced amino acid profile. However, soybean cultivation has other disadvantages, since it requires significant land and water resources, as well as pesticides, and the production for feed is also in competition with that for food. For the above-mentioned reasons, research in the last decade has been focusing on exploring alternative protein sources for animal feeding that possess remarkable sustainability attributes.
In this regard, insects are promising candidates to satisfy the nutritional needs of poultry species. Among them, the silkworm (
Bombyx mori) is a Lepidopteran species primarily reared for silk production in the textile industry. The pupae constitute a major by-product of sericulture, and, while utilized as a food source in certain regions, they are often discarded, leading to the loss of valuable nutrients and contributing to environmental concerns [
2]. From a nutritional perspective, silkworm meal (SWM) is rich in protein, with a protein content ranging from 52% to 80% (dry matter basis). It also provides a substantial lipid fraction, varying between 10% and 30% (dry matter basis), which includes healthy lipids, mainly polyunsaturated fatty acids of the omega-3 series. However, SWM also contains a bioactive compound, 1-deoxynojirimycin (1-DNJ), which could represent an issue for animal nutrition. In fact, 1-DNJ acts as a potent intestinal α-glucosidase inhibitor. Its mechanism of action involves competitive inhibition of specific glycosidase enzymes by mimicking the structural properties of normal carbohydrate substrates such as D-glucose and D-mannose [
3]. As a result, 1-DNJ can interfere with glycogenolysis, glycoprotein processing, and saccharide hydrolysis, necessitating a thorough evaluation of its implications when incorporating SWM into poultry diets. Furthermore, SWM contains chitin, a structural polysaccharide found in the exoskeletons of arthropods and other organisms. Research indicates that chitin can negatively impact nutrient absorption in poultry by forming complexes with essential nutrients, particularly proteins and lipids, thereby reducing their digestibility [
4]. Given these factors, while SWM presents potential as an alternative protein source in poultry nutrition, its dietary inclusion requires careful assessment to optimize nutrient utilization and mitigate any adverse effects.
Additionally, there are oil seed crops from which oil is extracted and cake is discarded or used as fertilizer, as it contains antinutritional compounds. Among them,
Camelina sativa stands out as a promising feedstuff due to its sustainability. This crop has a relatively short growth cycle, requires minimal agricultural inputs, and tolerates common
Brassica pests and diseases, thereby necessitating fewer treatments. Camelina meal contains around 35% protein content and fat ranging from 5% to 28% of residual oil, particularly rich in essential omega-3 fatty acids. Moreover, the use of improved lines with reduced antinutritional compounds (such as such as glucosinolates, phytic acid, sinapine, trypsin inhibitors, condensed tannins, and erucic acid) could facilitate their successful incorporation as raw feed material for livestock species, particularly poultry [
5,
6,
7,
8].
Determining in vivo digestibility is an effective method for assessing the nutritional value of poultry diets and evaluating digestive tract function, providing valuable information for feed evaluation and gastrointestinal health status. However, this technique is well-known for its disadvantages, including the use of animals, cost considerations, and the fact that it is time-consuming.
It is also accepted that some in vitro digestibility protocols correlate well with in vivo counterparts, with the advantages being that in vitro studies are easier to conduct, faster, and cheaper and do not directly involve animals. This technique has been successfully used to screen or assess the digestibility of possible feed ingredients in different animal species and humans. Generally, in vitro digestibility is investigated using commercially available enzymes or enzymes extracted from swine to represent poultry species [
9,
10,
11,
12,
13,
14,
15]. However, utilizing crude enzymes extracted from the same animal species would be preferable for such feed screening tests. Therefore, the present study aimed to explore the potential of an in vitro digestibility technique using crude enzymes extracted from broiler chickens and quails to preliminarily screen novel feedstuff candidates for poultry. This involved a critical evaluation of the tested in vitro method, both in absolute terms and by comparing results with in vivo trials, when available.
3. Results
The chemical composition of different diets for broiler chickens containing SWM, as well as the results concerning the in vitro digestibility, are presented in
Table 2,
Table 3, and
Table 4. The experimental diets included Starter, Grower, and Finisher versions. The crude protein content was balanced among the diets, which had been designed to be isonitrogenic and isoenergetic. The ether extract content was slightly higher in the SWM diets compared to the Control diets for each growth phase. The in vitro digestibility results highlighted that the Control Finisher diet displayed lower digestibility than the SWM Starter diet (
p < 0.001), while the other groups had intermediate results. The same outcome was observed for the OM digestibility (
p < 0.001). CP was better digested in the SWM Starter and Finisher diets compared to the Control Grower and Control Finisher diets (
p < 0.001), while the Control Starter and SWM Grower diets exhibited intermediate values. Correlation coefficient analysis showed that DMd had a strong and positive correlation with the DM diet (r = 0.894), CP diet (r = 0.914), and EE diet (r = 0.909), and a lower, yet significant, positive correlation with the Ash diet (r = 0.411). In contrast, a negative correlation was observed with the CF diet (r = −0.416). The correlations of OMd with the DM diet, CP diet, EE diet, Ash diet, and CF diet were almost overlapping with the above-mentioned correlations for DMd. CPd was significantly correlated with all the considered digestibility traits, but the relationships were not as robust as those described for DMd and CPd.
Table 5 presents the chemical composition of the diets for broiler quails containing different lines, sowing dates, and inclusion levels of
Camelina sativa cake, while
Table 6 and
Table 7 show the resulting in vitro digestibility outcomes and related correlation coefficients, respectively. Unlike the previous in vitro digestibility test, the Control diet displayed comparable outcomes with all treatment groups, as no significant differences were observed. The digestibility coefficients provided different information compared to the previous trial: specifically, the digestibility coefficients showed non-significant correlations with dietary components (DM diet, Ash diet, CP diet, EE diet, CF diet).
The results concerning the inclusion of camelina cake (same inclusion level, but different lines and sowing dates) or oil (Pearl line, different sowing dates) in the diets for laying quails are depicted in
Table 8 (feed),
Table 9 (in vitro digestibility), and
Table 10 (correlation coefficients). The experimental diets were well balanced in terms of the CP content, which was a formulation target. The digestibility results highlighted that feed containing camelina oil (both Spring and Autumn sowing dates) and cake (Alan Spring) had the highest DMd (
p < 0.05), CPd (
p < 0.01), and OMd (
p < 0.05), while the Pearl Spring diet displayed the worst results. Looking at the correlation coefficients, DMd and OMd were significantly correlated with the EE diet (r = −0.355 and r = −0.367, respectively) and OMd with the CF diet (r = 0.310) and Ash diet (r = 0.310), even though the coefficients were low in absolute terms. In addition, CPd was negatively correlated with the CP diet (r = −0.539), EE diet (r = −0.486), and CF diet (r = −0.154), also in this case with moderate to low coefficients.
4. Discussion
It is widely accepted that in vivo animal studies are ideal to provide accurate information on nutrient utilization, which can reliably predict performance. In contrast, in vitro methods (especially for poultry species) cannot fully replace in vivo studies. However, the development of in vitro protocols has garnered significant research attention for several reasons, including the short time required for evaluations, minimal personnel needs, low economic investment, and the ability to conduct assays in virtually any laboratory [
18]. Additionally, the use of in vitro studies addresses ethical concerns associated with animal use in scientific research and avoids various factors (such as genetics, environmental conditions, potential disease occurrence, and management practices) that can affect in vivo digestibility results. The development of specific in vitro digestibility methods for poultry has been relatively recent, as historically, in vitro data for monogastric species, such as pigs, were adapted for poultry studies [
11]. Given the significant differences in digestive tract anatomy between poultry and pigs (such as the presence of a crop and gizzard in poultry and a well-developed hindgut in pigs), implementing in vitro digestibility protocols specifically designed for poultry is advisable. This approach is particularly pertinent considering the importance of poultry in global animal production. [
19]. An additional beneficial aspect of in vitro enzymatic digestion protocols, as adopted in the present study, is the use of crude digestive enzymes extracted from the same animal species and/or individuals intended for future in vivo studies. This method enhances the potential for better correlation coefficients between in vitro and in vivo results. [
20]. However, this approach presents a possible drawback: sampling digestive organs from slaughtered animals means that enzyme concentration and activity can be affected by the chemical composition of the diets the animals were consuming. This variability can influence observed enzymatic activity and, consequently, affect the outcomes of in vitro digestibility assays utilizing the extracted enzymes. In fact, the results presented in
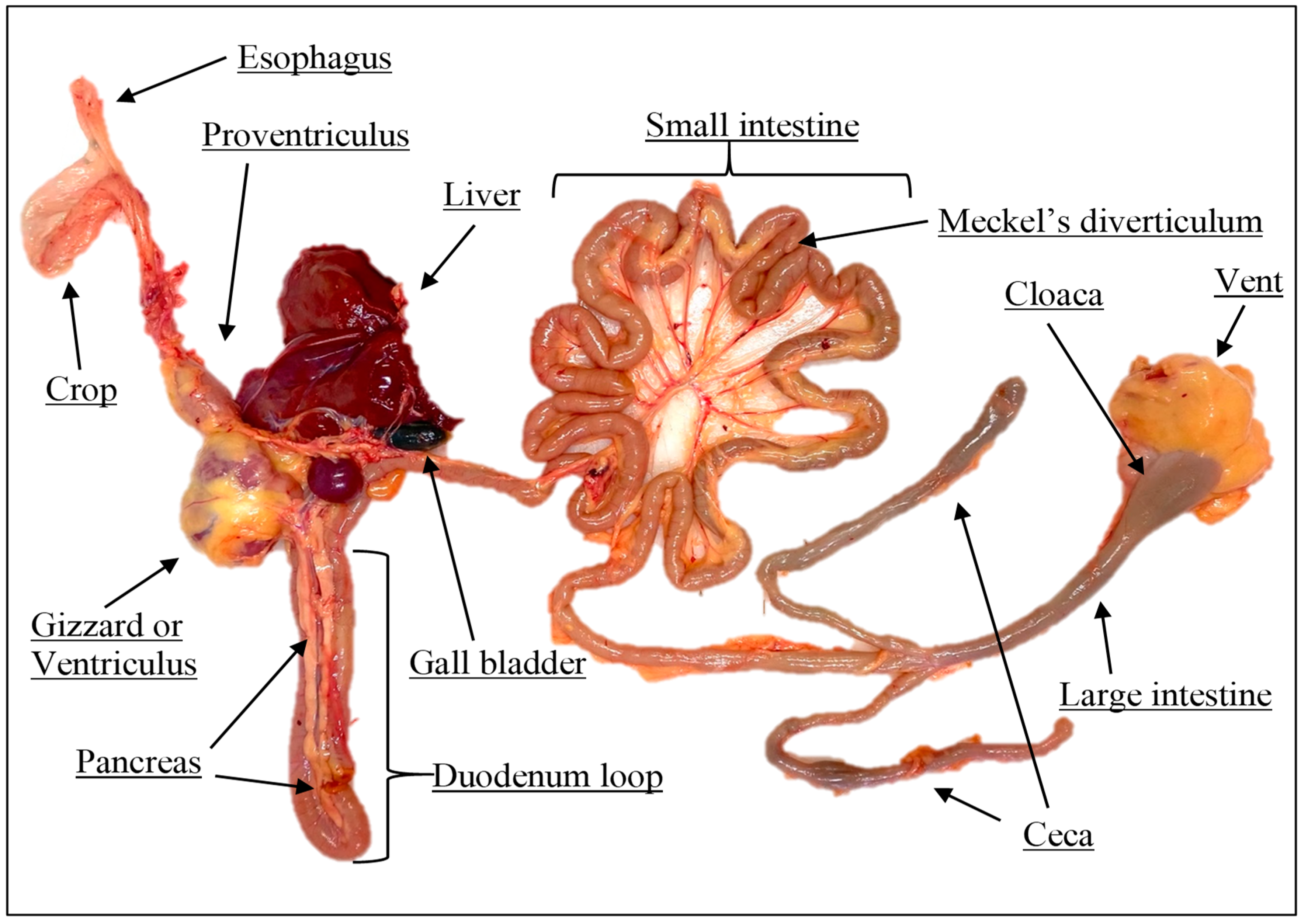
Table 11 highlight that the crude enzyme extracts from the proventriculus, pancreas, and duodenum of 42-day-old broiler chickens vs. 35-day-old quails showed remarkable differences in amylase activity (11.8 vs. 1.01 U/min/mL for broiler chickens and broiler quails, respectively).
The first consideration arising from the results obtained with the present in vitro digestibility assay is that, in absolute terms, the digestibility of DM, OM, and CP showed lower values compared to the usual in vivo studies for the same species for complete diets. For example, recent research testing the dietary inclusion of 12.5% SWM meal (full-fat) in the diet for broiler quails showed approximately 51% DM, 70% CP, and 54% OM digestibility [
21], which are remarkably higher than the values overall observed in the present research. This was only partly surprising, as experimental diets are complex food matrices composed of various ingredients, each with distinct nutritional and antinutritional properties. This complexity is particularly relevant because in vitro digestibility assays are static systems, unlike the dynamic gastrointestinal tract (GIT) of an animal, which involves feed–physiology interactions and responses to ingested feed. In vitro systems lack this dynamic interaction, potentially limiting their ability to accurately simulate the digestive process. In fact, enzymatic in vitro assays can be hindered by non-digestible constituents, such as plant cell walls or insoluble dietary fibers, which may increase in the diet compared to the raw materials. The swelling and viscosity behavior of these components impact macronutrient digestion and absorption by reducing enzyme diffusion, thus limiting their action [
22]. In addition, enzyme dilution and substrate (diet) concentration play a pivotal role in determining the level of hydrolysis and absorption of nutrients. Increasing substrate concentration can enhance the rate of hydrolysis up to a certain point. Once all available enzymes are saturated, any further increase in substrate concentration will not affect the hydrolysis rate, as the enzymes are already operating at their maximum capacity. This suggests that increasing the enzyme concentration or decreasing the dilution rate, as well as reducing the quantity of dietary substrate used in the test, may be considered in future research.
Literature data indicate that, for the all the above-mentioned reasons, dietary ingredients should be considered individually when performing in vitro digestibility experiments, and that, at present, this technique could be successfully used to screen the potential of new emerging feedstuffs, but without claiming to be a reference for feed formulations [
18]. An example in this sense is a recent publication [
15] that successfully screened 18 feed ingredients intended for different poultry species with very diverse nutritional profiles (DM digestibility was between 40% and 72%, and CP digestibility ranged from 15% to 47%). Nevertheless, there are also cases in which also certain feedstuffs, such as lupin meal, have been demonstrated to be unsuitable for in vitro digestion models [
23]; this is attributable to possible interferences in the digestion process due to the presence of antinutritional factors in the tested raw material.
The adequacy of in vitro digestibility studies for single feed ingredients rather than complete diets emerges in the present research too: in fact, when the results of the in vitro digestibility of SWM alone are considered (
Table 2), a different scenario appears than that for experimental diets: the SWM digestibility values for DM (48.9%), OM (46.5%), and CP (47.7%) were consistent with data published by Kovitvadhi et al. [
15], in which the in vitro digestibility values of SWM (broiler chicken) for DM, OM, and CP were 67%, 65% and 41%, respectively. Overall, the digestibility results were also consistent with other studies where single ingredients were tested for broiler chickens and meat-type ducks [
13,
14], as well as black-meat chickens and quail [
15].
Despite the evident limitations of the in vitro digestibility protocol tested in the present trial regarding the quantification, in absolute terms, of DMd, CPd, and OMd, the observed dietary effects were consistent with existing in vivo results. Specifically, the same diets including the SWM for broiler chickens were also tested in a trial of broiler chickens, where performance traits were considered [
6]. The results indicated that birds displayed similar live weight, average daily gain, feed intake, and FCR, thus suggesting the reliability of the results displayed in
Table 2, whereas SWM diets showed comparable digestibility values with Control diets.
In contrast, experimental diets containing
Camelina sativa cake and subjected to in vitro digestion exploiting crude enzyme extract from 35-day-old quails (
Table 6 and
Table 9) provided different indications: on the one hand, the incorporation of different camelina lines (Alan vs. Pearl), sowing dates (spring vs. autumn), and inclusion levels (0% vs. 5% vs. 10%) did not affect the DMd, CPd, and OMd. On the other hand, the incorporation of 15% camelina cake of the Pearl cultivar digested in vitro using the same crude enzyme extract (35-day-old quails) displayed a worse DMd, CPd, and OMd than the Control diet. Camelina is an alternative emerging oilseed crop for poultry dets, thanks to different interesting features including its tolerance to cold climates and drought, the low requirement for pesticides and fertilizers, as well as a remarkable
n-3 FA content, as well as antioxidants [
7]. However, camelina also contains glucosinolates, sinapine, condensed tannins, trypsin inhibitors, and phytic acid, which are antinutritional factors that, at certain ingestion levels, can have negative effects on digestibility by poultry, and thus performance [
24]. Glucosinolates, in particular, once hydrolyzed in the animal intestine, generate some toxic products (isothiocyanates, thiocyanates, and nitriles) which are known to interfere with thyroid and liver function, and which can have negative effects on animal health and performance [
25].
The results of the present study indicate that the Pearl Spring 15 diet had worse in vitro digestibility outcomes, while the same was not observed for other camelina cultivars with different sowing dates, even considering the same inclusion level. This was not surprising, and is attributable to different aspects: while the Alan cultivar was genetically engineered to have a low glucosinolate content [
7], the Pearl cultivar was not. In addition, differences in the sowing season are known to produce seeds with different chemical characteristics [
26], and thus also different glucosinolate contents [
27]. Last but not least, the inclusion level is a key factor in determining digestibility, as it determines also the absolute amounts of antinutritional factors present in the diet and thus ingested by the animal, in this case quail. The above-mentioned hypotheses found confirmation in a trial studying the incorporation of different
Camelina sativa cakes (15% inclusion rate) into diets for broiler quails [
7], where the Pearl group exhibited worse live performance than the Control group.



 ,
, 



{kind=link}