
The PCV3 Cap Virus-like Particle Vaccine with the Chimeric PCV2-Neutralizing Epitope Gene Is Effective in Mice
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Viruses and Cells
2.2. Plasmid Construction
2.3. Protein Expression and Purification
2.4. Assembly and Identification of VLPs In Vitro
2.5. Animal Experimental Design
2.6. Enzyme-Linked Immunosorbent Assay
2.7. Virus Neutralization Test
2.8. Spleen Lymphocyte Proliferation Assay
2.9. Statistical Analysis
3. Results
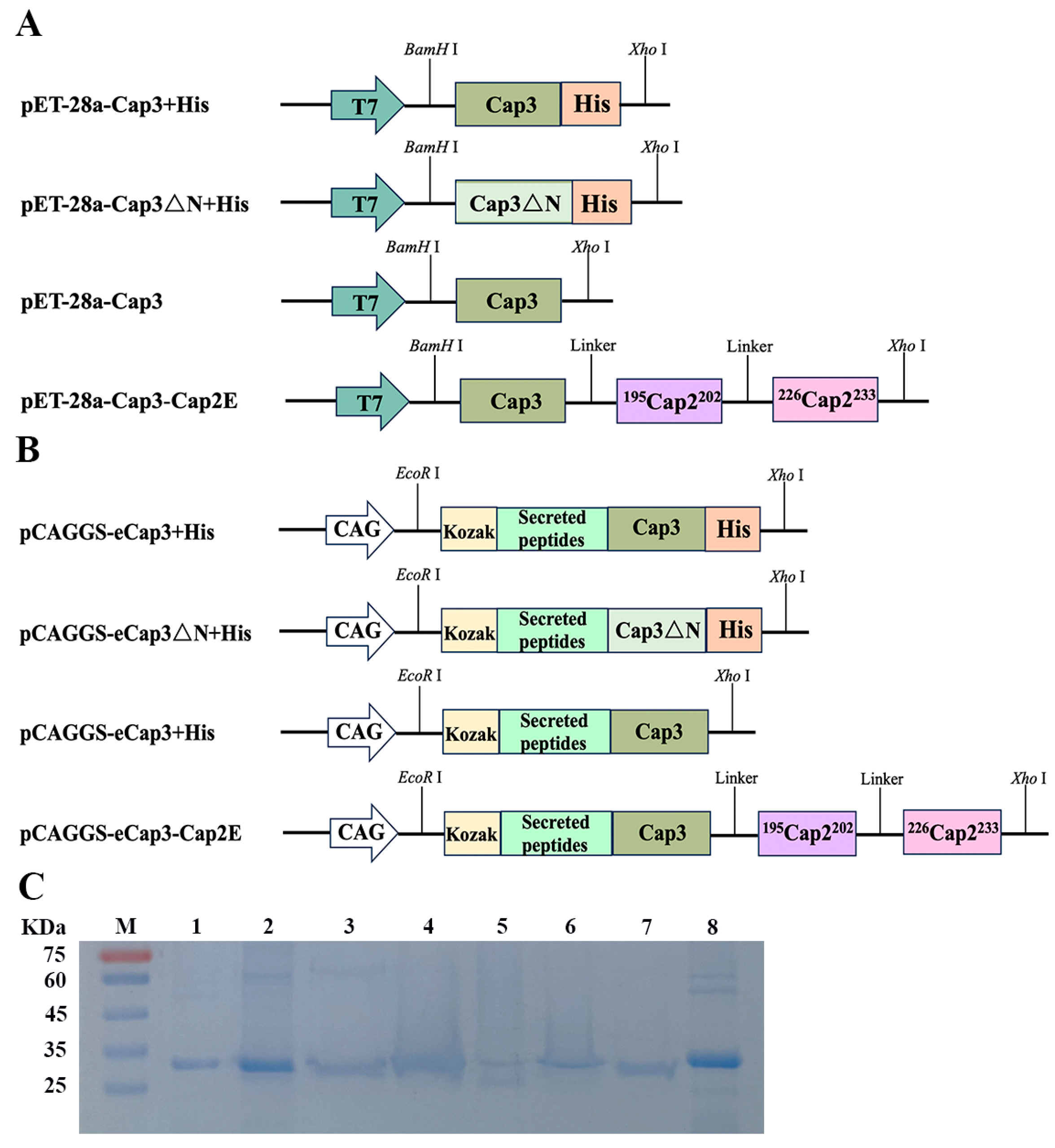
3.1. Plasmid Construction and Protein Purification
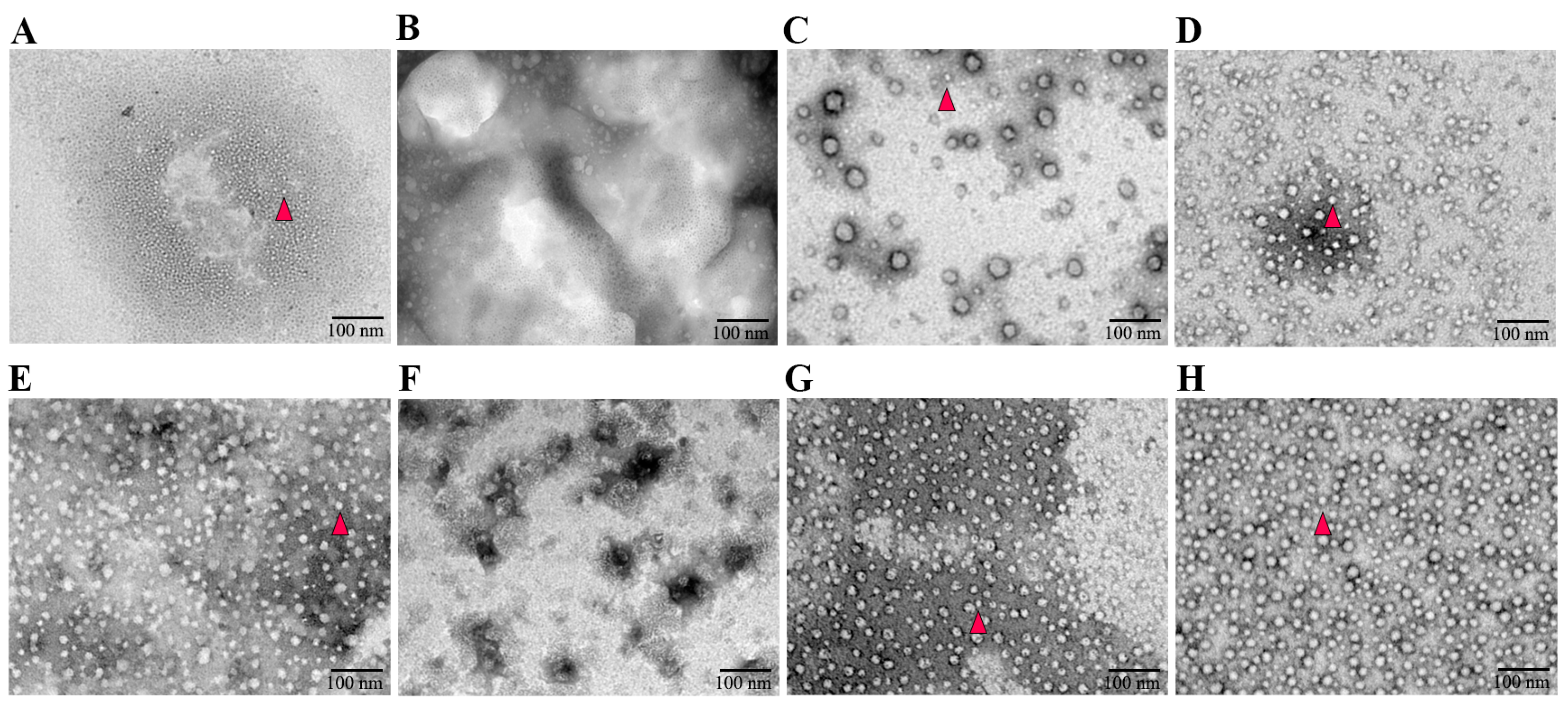
3.2. The Absence of an NLS Prevented the Formation of PCV3 Cap VLPs
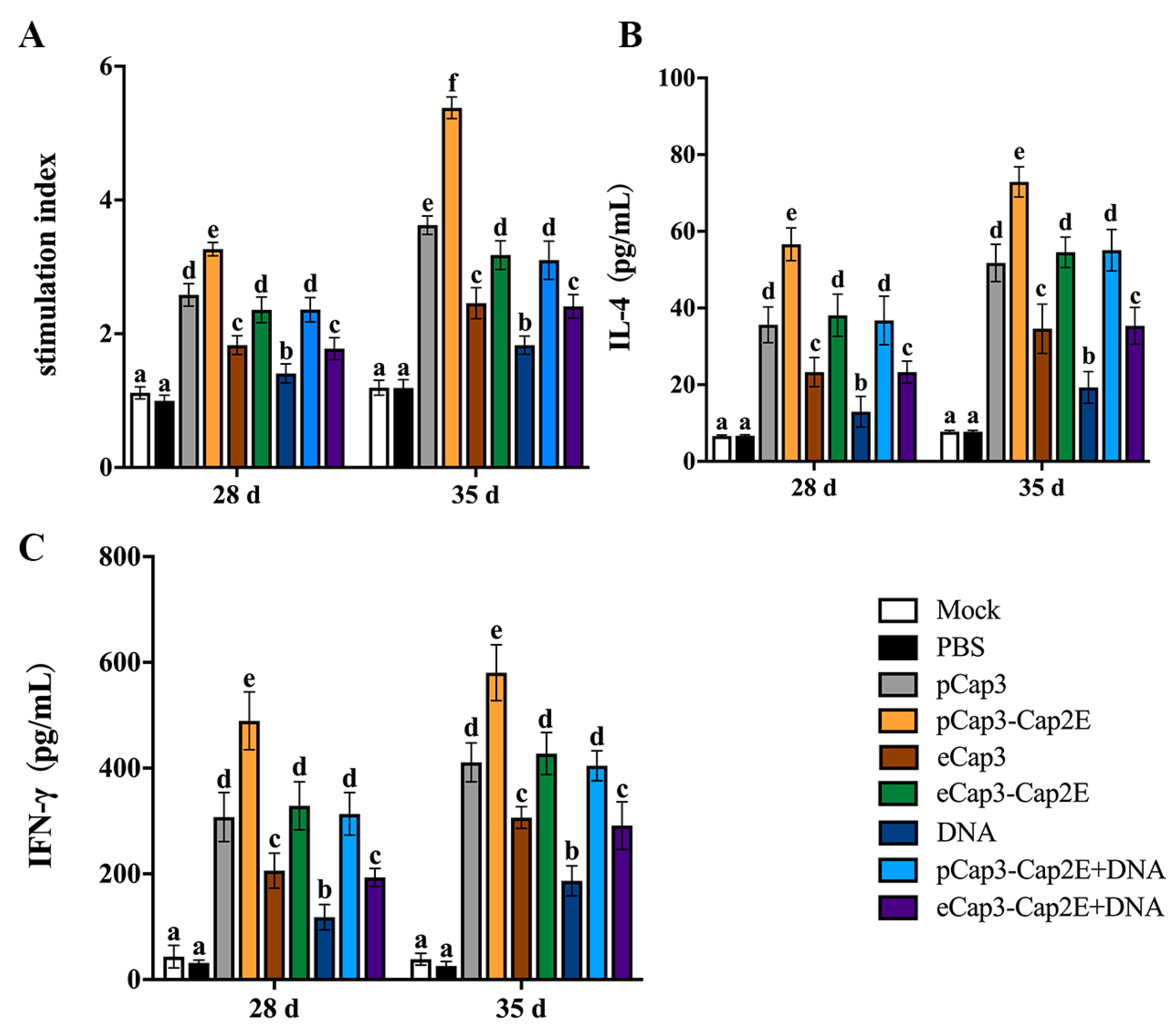
3.3. pCap3-Cap2E VLPs Activated Higher Humoral Immune Responses Than the Other Inoculated Groups
3.4. pCap3-Cap2E VLPs Activated Higher Cellular Immune Responses Than the Other Inoculation Groups
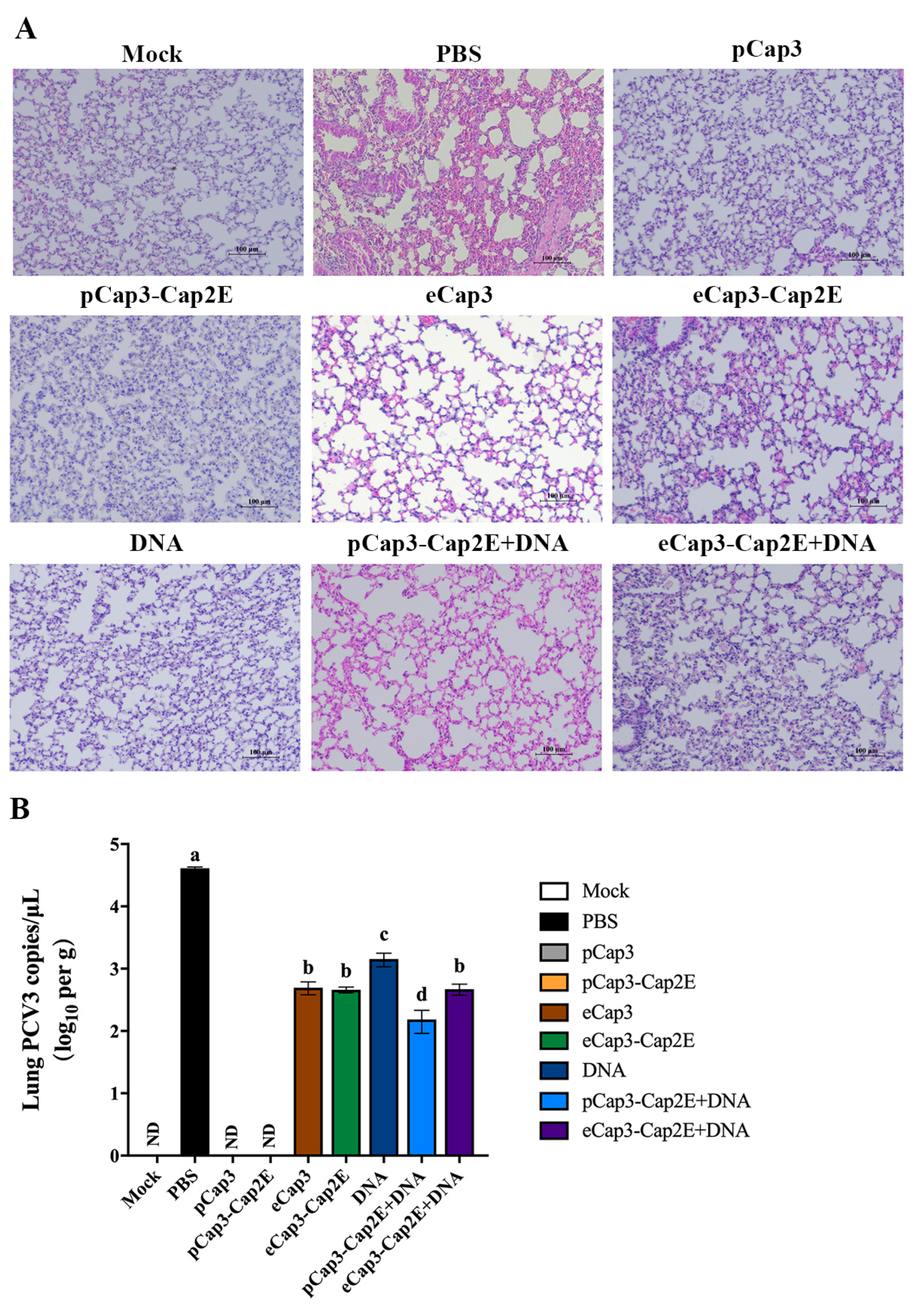
3.5. Viral Load Was Lower and Pathological Damage to the Lungs Was Less Severe in Mice Vaccinated with pCap3-Cap2E VLPs Than in Other Groups
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, J.; Wang, Y.; Zhou, L.; Qiu, Y.; Zhao, J.; Dai, B.; Feng, X.; Hou, L.; Liu, J. Interaction Network of Porcine Circovirus Type 3 and 4 Capsids with Host Proteins. Viruses 2022, 14, 939. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Chen, J.; Wu, X.; Ma, D.; Zhang, X.; Li, R.; Han, C.; Liu, H.; Yin, X.; Du, Q.; et al. PCV2 targets cGAS to inhibit type I interferon induction to promote other DNA virus infection. PLoS Pathog. 2021, 17, e1009940. [Google Scholar] [CrossRef]
- Vargas-Bermudez, D.S.; Mogollón, J.D.; Jaime, J. The Prevalence and Genetic Diversity of PCV3 and PCV2 in Colombia and PCV4 Survey during 2015–2016 and 2018–2019. Pathogens 2022, 11, 633. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Wang, D.; Wang, J.; Zhu, S.; She, R.; Ren, X.; Tian, J.; Quan, R.; Hou, L.; Li, Z.; et al. Induction of Porcine Dermatitis and Nephropathy Syndrome in Piglets by Infection with Porcine Circovirus Type 3. J. Virol. 2019, 93, e02045-18. [Google Scholar] [CrossRef]
- Kedkovid, R.; Woonwong, Y.; Arunorat, J.; Sirisereewan, C.; Sangpratum, N.; Kesdangsakonwut, S.; Tummaruk, P.; Teankum, K.; Assavacheep, P.; Jittimanee, S.; et al. Porcine circovirus type 3 (PCV3) shedding in sow colostrum. Vet. Microbiol. 2018, 220, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Huang, M.; Wang, Y.; Chen, Y.; Yang, H.; Quan, F. Immunogenicity Analysis of PCV3 Recombinant Capsid Protein Virus-like Particles and Their Application in Antibodies Detection. Int. J. Mol. Sci. 2023, 24, 10377. [Google Scholar] [CrossRef]
- Mengfan, Q.; Xifeng, W.; Guowu, Z.; Qingling, M.; Jun, Q.; Lixia, W.; Kuojun, C.; Jinsheng, Z.; Zaichao, Z.; Weiwei, Y.; et al. Molecular Detection and Genetic Diversity of porcine Circovirus Type 3 in Commercial Pig Farms in Xinjiang Province, China. J. Vet. Res. 2019, 63, 481–488. [Google Scholar] [CrossRef]
- Kim, S.H.; Park, J.Y.; Jung, J.Y.; Kim, H.Y.; Park, Y.R.; Lee, K.K.; Lyoo, Y.S.; Yeo, S.G.; Park, C.K. Detection and genetic characterization of porcine circovirus 3 from aborted fetuses and pigs with respiratory disease in Korea. J. Vet. Sci. 2018, 19, 721–724. [Google Scholar] [CrossRef]
- Shang, S.B.; Jin, Y.L.; Jiang, X.T.; Zhou, J.Y.; Zhang, X.; Xing, G.; He, J.L.; Yan, Y. Fine mapping of antigenic epitopes on capsid proteins of porcine circovirus, and antigenic phenotype of porcine circovirus type 2. Mol. Immunol. 2009, 46, 327–334. [Google Scholar] [CrossRef]
- Qi, S.; Su, M.; Guo, D.; Li, C.; Wei, S.; Feng, L.; Sun, D. Molecular detection and phylogenetic analysis of porcine circovirus type 3 in 21 Provinces of China during 2015–2017. Transbound. Emerg. Dis. 2019, 66, 1004–1015. [Google Scholar] [CrossRef]
- Liu, B.Y.; Gao, B.; Liu, M.Z.; Zhang, T.T.; Liu, B.S.; Chen, Z.L. High repetitive arginine in the anterior of PCV3 capsid protein is a severe obstacle for its expression in E. coli. AMB Express 2020, 10, 214. [Google Scholar] [CrossRef] [PubMed]
- Nooraei, S.; Bahrulolum, H.; Hoseini, Z.S.; Katalani, C.; Hajizade, A.; Easton, A.J.; Ahmadian, G. Virus-like particles: Preparation, immunogenicity and their roles as nanovaccines and drug nanocarriers. J. Nanobiotechnol. 2021, 19, 59. [Google Scholar] [CrossRef] [PubMed]
- Chackerian, B.; Durfee, M.R.; Schiller, J.T. Virus-like display of a neo-self antigen reverses B cell anergy in a B cell receptor transgenic mouse model. J. Immunol. 2008, 180, 5816–5825. [Google Scholar] [CrossRef] [PubMed]
- Fontana, D.; Kratje, R.; Etcheverrigaray, M.; Prieto, C. Rabies virus-like particles expressed in HEK293 cells. Vaccine 2014, 32, 2799–2804. [Google Scholar] [CrossRef] [PubMed]
- Ninyio, N.N.; Ho, K.L.; Omar, A.R.; Tan, W.S.; Iqbal, M.; Mariatulqabtiah, A.R. Virus-like Particle Vaccines: A Prospective Panacea Against an Avian Influenza Panzootic. Vaccines 2020, 8, 694. [Google Scholar] [CrossRef] [PubMed]
- Masuda, A.; Lee, J.M.; Miyata, T.; Sato, T.; Hayashi, S.; Hino, M.; Morokuma, D.; Karasaki, N.; Mon, H.; Kusakabe, T. Purification and characterization of immunogenic recombinant virus-like particles of porcine circovirus type 2 expressed in silkworm pupae. J. Gen. Virol. 2018, 99, 917–926. [Google Scholar] [CrossRef] [PubMed]
- Frietze, K.M.; Peabody, D.S.; Chackerian, B. Engineering virus-like particles as vaccine platforms. Curr. Opin. Virol. 2016, 18, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Lua, L.H.; Connors, N.K.; Sainsbury, F.; Chuan, Y.P.; Wibowo, N.; Middelberg, A.P. Bioengineering virus-like particles as vaccines. Biotechnol. Bioeng. 2014, 111, 425–440. [Google Scholar] [CrossRef] [PubMed]
- Xi, X.; Mo, X.; Xiao, Y.; Yin, B.; Lv, C.; Wang, Y.; Sun, Z.; Yang, Q.; Yao, Y.; Xuan, Y.; et al. Production of Escherichia coli-based virus-like particle vaccine against porcine circovirus type 2 challenge in piglets: Structure characterization and protective efficacy validation. J. Biotechnol. 2016, 223, 8–12. [Google Scholar] [CrossRef]
- Park, J.E.; Kim, J.H.; Park, J.Y.; Jun, S.H.; Shin, H.J. A chimeric MERS-CoV virus-like particle vaccine protects mice against MERS-CoV challenge. Virol. J. 2022, 19, 112. [Google Scholar] [CrossRef]
- Tabatabaei, M.S.; Ahmed, M. Enzyme-Linked Immunosorbent Assay (ELISA). Methods Mol. Biol. 2022, 2508, 115–134. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Du, Q.; Wu, B.; Li, J.; Chang, L.; Zhao, X.; Huang, Y.; Tong, D. Immunogenicity of adenovirus vaccines expressing the PCV2 capsid protein in pigs. Vaccine 2017, 35, 4722–4729. [Google Scholar] [CrossRef] [PubMed]
- Bera, B.C.; Choudhary, M.; Anand, T.; Virmani, N.; Sundaram, K.; Choudhary, B.; Tripathi, B.N. Detection and genetic characterization of porcine circovirus 3 (PCV3) in pigs in India. Transbound. Emerg. Dis. 2020, 67, 1062–1067. [Google Scholar] [CrossRef] [PubMed]
- Palinski, R.; Piñeyro, P.; Shang, P.; Yuan, F.; Guo, R.; Fang, Y.; Byers, E.; Hause, B.M. A Novel Porcine Circovirus Distantly Related to Known Circoviruses Is Associated with Porcine Dermatitis and Nephropathy Syndrome and Reproductive Failure. J. Virol. 2017, 91, e01879-16. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Zhang, Y.H.; Tian, R.B.; Hou, C.Y.; Li, X.S.; Zheng, L.L.; Wang, L.Q.; Chen, H.Y. Prevalence and genetic analysis of porcine circovirus type 2 (PCV2) and type 3 (PCV3) between 2018 and 2020 in central China. Infect. Genet. Evol. 2021, 94, 105016. [Google Scholar] [CrossRef] [PubMed]
- Saporiti, V.; Cruz, T.F.; Correa-Fiz, F.; Núñez, J.I.; Sibila, M.; Segalés, J. Similar frequency of Porcine circovirus 3 (PCV-3) detection in serum samples of pigs affected by digestive or respiratory disorders and age-matched clinically healthy pigs. Transbound. Emerg. Dis. 2020, 67, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Beach, N.M.; Ramamoorthy, S.; Opriessnig, T.; Wu, S.Q.; Meng, X.J. Novel chimeric porcine circovirus (PCV) with the capsid gene of the emerging PCV2b subtype cloned in the genomic backbone of the non-pathogenic PCV1 is attenuated in vivo and induces protective and cross-protective immunity against PCV2b and PCV2a subtypes in pigs. Vaccine 2010, 29, 221–232. [Google Scholar] [CrossRef]
- Du, Q.; Shi, T.; Wang, H.; Zhu, C.; Yang, N.; Tong, D.; Huang, Y. The ultrasonically treated nanoliposomes containing PCV2 DNA vaccine expressing gC1qR binding site mutant Cap is efficient in mice. Front. Microbiol. 2022, 13, 1077026. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Sun, Y.; He, Q.; Sun, C.; Dong, T.; Zhang, L.; Zhan, Y.; Wang, N.; Yang, Y.; Sun, Y. Mitochondrial Localization Signal of Porcine Circovirus Type 2 Capsid Protein Plays a Critical Role in Cap-Induced Apoptosis. Vet. Sci. 2021, 8, 272. [Google Scholar] [CrossRef]
- Mo, X.; Li, X.; Yin, B.; Deng, J.; Tian, K.; Yuan, A. Structural roles of PCV2 capsid protein N-terminus in PCV2 particle assembly and identification of PCV2 type-specific neutralizing epitope. PLoS Pathog. 2019, 15, e1007562. [Google Scholar] [CrossRef]
- Klatt, S.; Konthur, Z. Secretory signal peptide modification for optimized antibody-fragment expression-secretion in Leishmania tarentolae. Microb. Cell Fact. 2012, 11, 97. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Gong, C.; Ren, L.; Li, X.; Song, D.; Zheng, G. A simple and effective strategy for solving the problem of inclusion bodies in recombinant protein technology: His-tag deletions enhance soluble expression. Appl. Microbiol. Biotechnol. 2013, 97, 837–845. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, Y.; Zhang, G.; Wang, A.; Dong, Z.; Qi, Y.; Wang, J.; Zhao, B.; Li, N.; Jiang, M. Human papillomavirus L1 protein expressed in Escherichia coli self-assembles into virus-like particles that are highly immunogenic. Virus Res. 2016, 220, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Tan, F.H.; Ng, J.F.; Mohamed Alitheen, N.B.; Muhamad, A.; Yong, C.Y.; Lee, K.W. A simple and high efficiency purification of His-tagged turnip yellow mosaic virus-like particle (TYMV-VLP) by nickel ion affinity precipitation. J. Virol. Methods 2023, 319, 114771. [Google Scholar] [CrossRef] [PubMed]
- Veena Rani, N.; Kapoor, N.; Krishnan, A. Efficient generation and characterization of chimeric dengue viral-like particles. Biochem. Biophys. Res. Commun. 2023, 654, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Imagawa, T.; Ito, M.; Matsuda, M.; Nakashima, K.; Tokunaga, Y.; Ohta, I.; Li, T.C.; Suzuki, R.; Suzuki, T. Virus-like particles with FLAG-tagged envelope protein as a tetravalent dengue vaccine candidate. Sci. Rep. 2021, 11, 17542. [Google Scholar] [CrossRef]
- Vemula, S.V.; Sayedahmed, E.E.; Sambhara, S.; Mittal, S.K. Vaccine approaches conferring cross-protection against influenza viruses. Expert. Rev. Vaccines 2017, 16, 1141–1154. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Deng, Y.; Niu, P.; Song, J.; Wang, W.; Du, Y.; Huang, B.; Wang, W.; Zhang, L.; Zhao, P.; et al. Co-Immunization With CHIKV VLP and DNA Vaccines Induces a Promising Humoral Response in Mice. Front. Immunol. 2021, 12, 655743. [Google Scholar] [CrossRef] [PubMed]
- Jung, B.K.; Kim, H.R.; Lee, Y.H.; Jang, H.; Chang, K.S. Comparison of Immune Responses to the PCV2 Replicase-Capsid and Capsid Virus-Like Particle Vaccines in Mice. J. Microbiol. Biotechnol. 2019, 29, 482–488. [Google Scholar] [CrossRef]
- Baha, S.; Zhang, M.; Behloul, N.; Liu, Z.; Wei, W.; Meng, J. Efficient production and characterization of immunogenic HEV-PCV2 chimeric virus-like particles. Vet. Microbiol. 2022, 268, 109410. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, X.; Wang, Q.; Lu, W.; Wang, Y.; Han, Z.; Liang, L.; Gao, S.; Ma, H.; Luo, X. The PCV3 Cap Virus-like Particle Vaccine with the Chimeric PCV2-Neutralizing Epitope Gene Is Effective in Mice. Vet. Sci. 2024, 11, 264. https://doi.org/10.3390/vetsci11060264
Wu X, Wang Q, Lu W, Wang Y, Han Z, Liang L, Gao S, Ma H, Luo X. The PCV3 Cap Virus-like Particle Vaccine with the Chimeric PCV2-Neutralizing Epitope Gene Is Effective in Mice. Veterinary Sciences. 2024; 11(6):264. https://doi.org/10.3390/vetsci11060264
Chicago/Turabian StyleWu, Xingchen, Qikai Wang, Wang Lu, Ying Wang, Zehao Han, Libin Liang, Shimin Gao, Haili Ma, and Xiaomao Luo. 2024. "The PCV3 Cap Virus-like Particle Vaccine with the Chimeric PCV2-Neutralizing Epitope Gene Is Effective in Mice" Veterinary Sciences 11, no. 6: 264. https://doi.org/10.3390/vetsci11060264
APA StyleWu, X., Wang, Q., Lu, W., Wang, Y., Han, Z., Liang, L., Gao, S., Ma, H., & Luo, X. (2024). The PCV3 Cap Virus-like Particle Vaccine with the Chimeric PCV2-Neutralizing Epitope Gene Is Effective in Mice. Veterinary Sciences, 11(6), 264. https://doi.org/10.3390/vetsci11060264