

Divergent Immune Responses to Minor Bovine Mastitis-Causing Pathogens

 , ,
, ,  , and
, and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Blood Collection, and Bovine Polymorphonuclear Neutrophil (PMN) Isolation
2.2. Bacterial Growth Condition
2.3. Fluorescent Labeling and Opsonization of Bacteria
2.4. Bovine Neutrophil Apoptosis Assay
2.5. Analysis of Surface Microbial Associated Molecular Pattern (MAMP) of Bovine Neutrophils
2.6. Measurement of Intracellular Reactive Oxygen Species (ROS)
2.7. Phagocytosis of Fluorescently Labeled Bacteria
2.8. Bacterial Killing and Spot Dilution Assay
2.9. Gene Expression Using Real-Time PCR (qPCR)
2.10. Data Analysis
3. Results
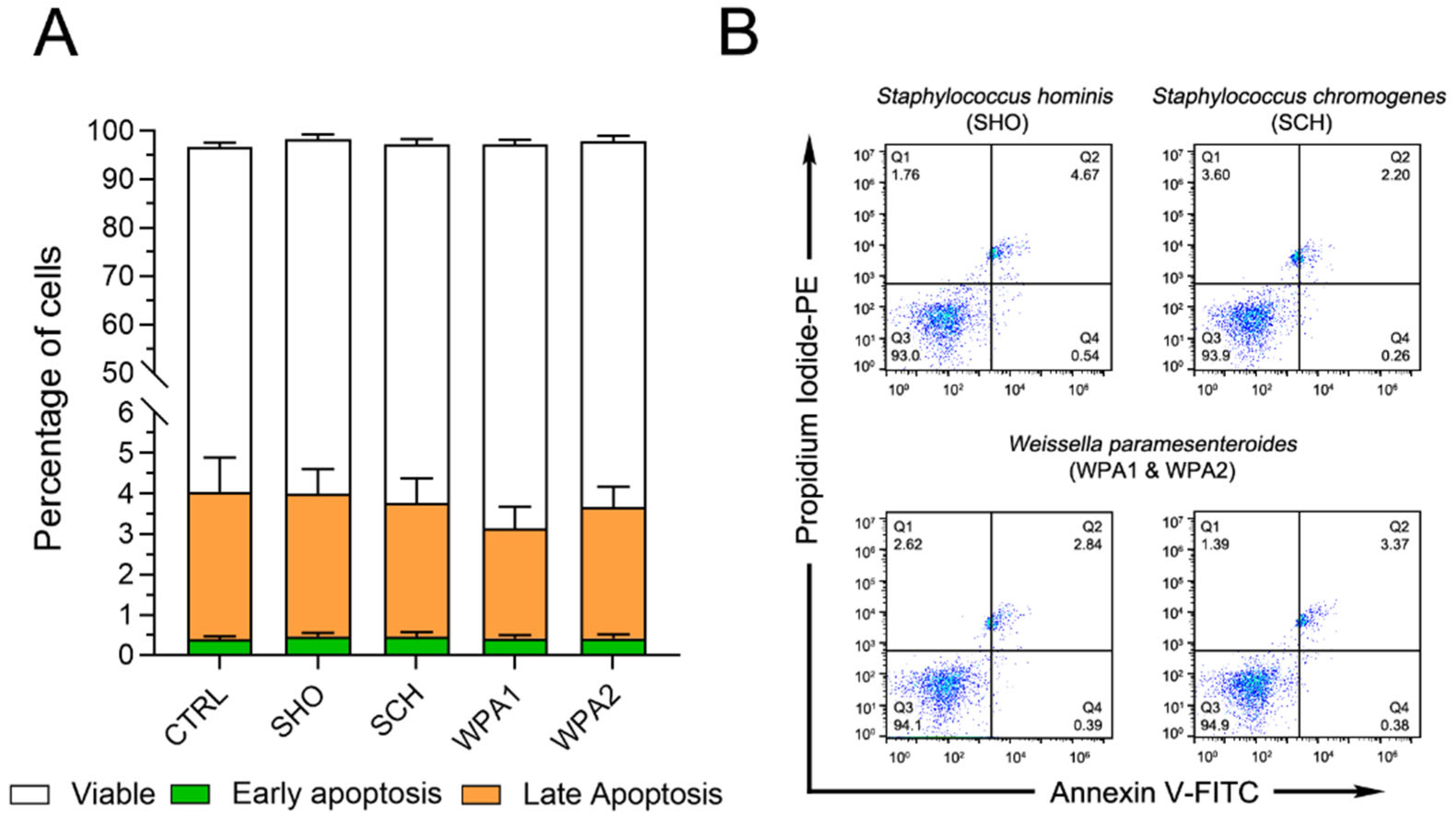
3.1. Bovine Neutrophils Showed Little Response to Minor Pathogens in Terms of Programmed Cell Death Initiation
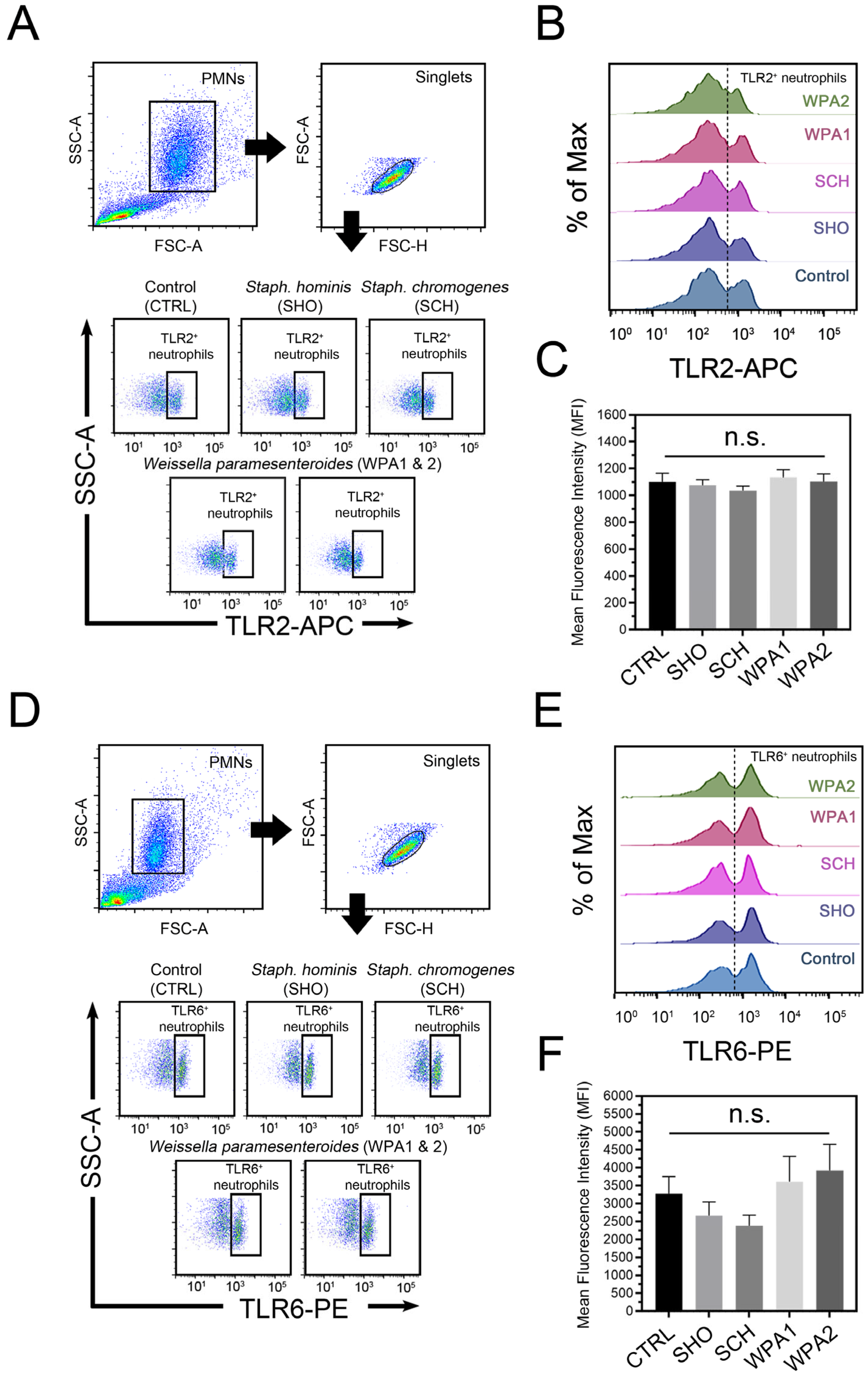
3.2. Specific Microbial Associated Molecular Patterns (MAMPs) on Bovine Neutrophils Were Identified and Quantified
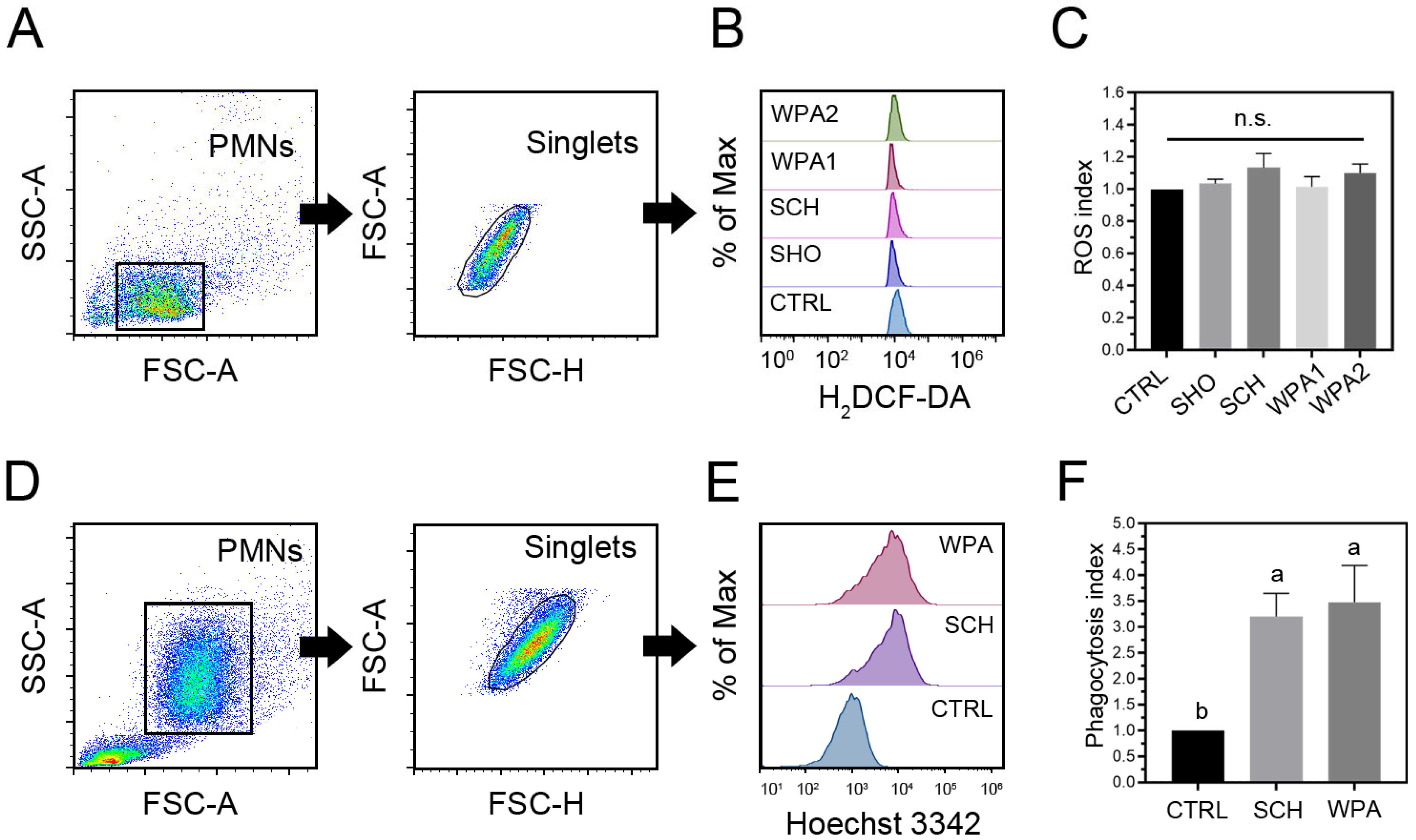
3.3. Exposure to Minor Mastitis-Causing Pathogens Demonstrated a Modest Magnitude of Intracellular ROS in Bovine Neutrophils
3.4. Minor Mastitis Bacteria Substantially Boosted the Ability of Bovine Neutrophils to Engulf the Bacteria
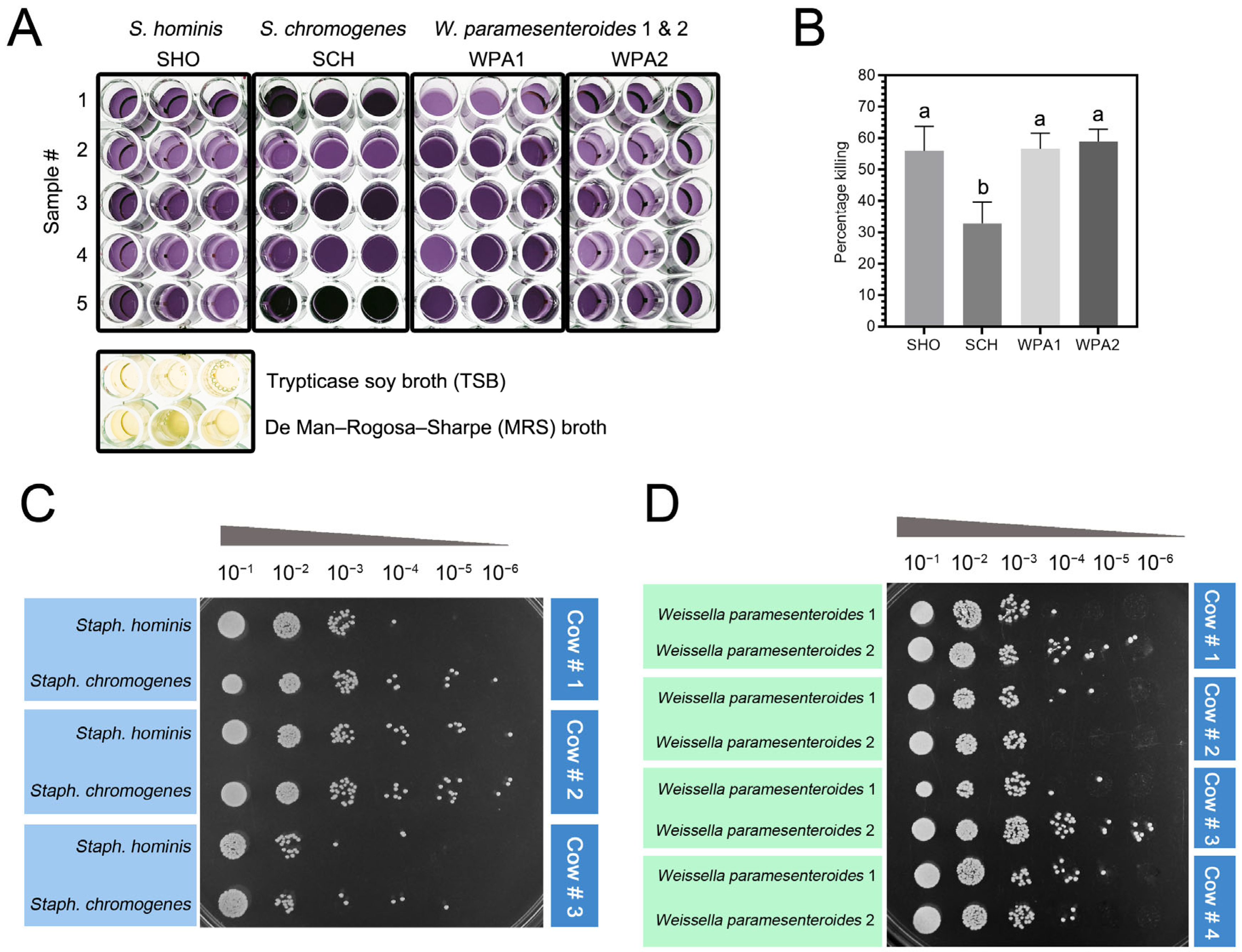
3.5. Staphylococcus Chromogenes Exhibited Potential Resistance to the Bactericidal Activity of Bovine Neutrophils
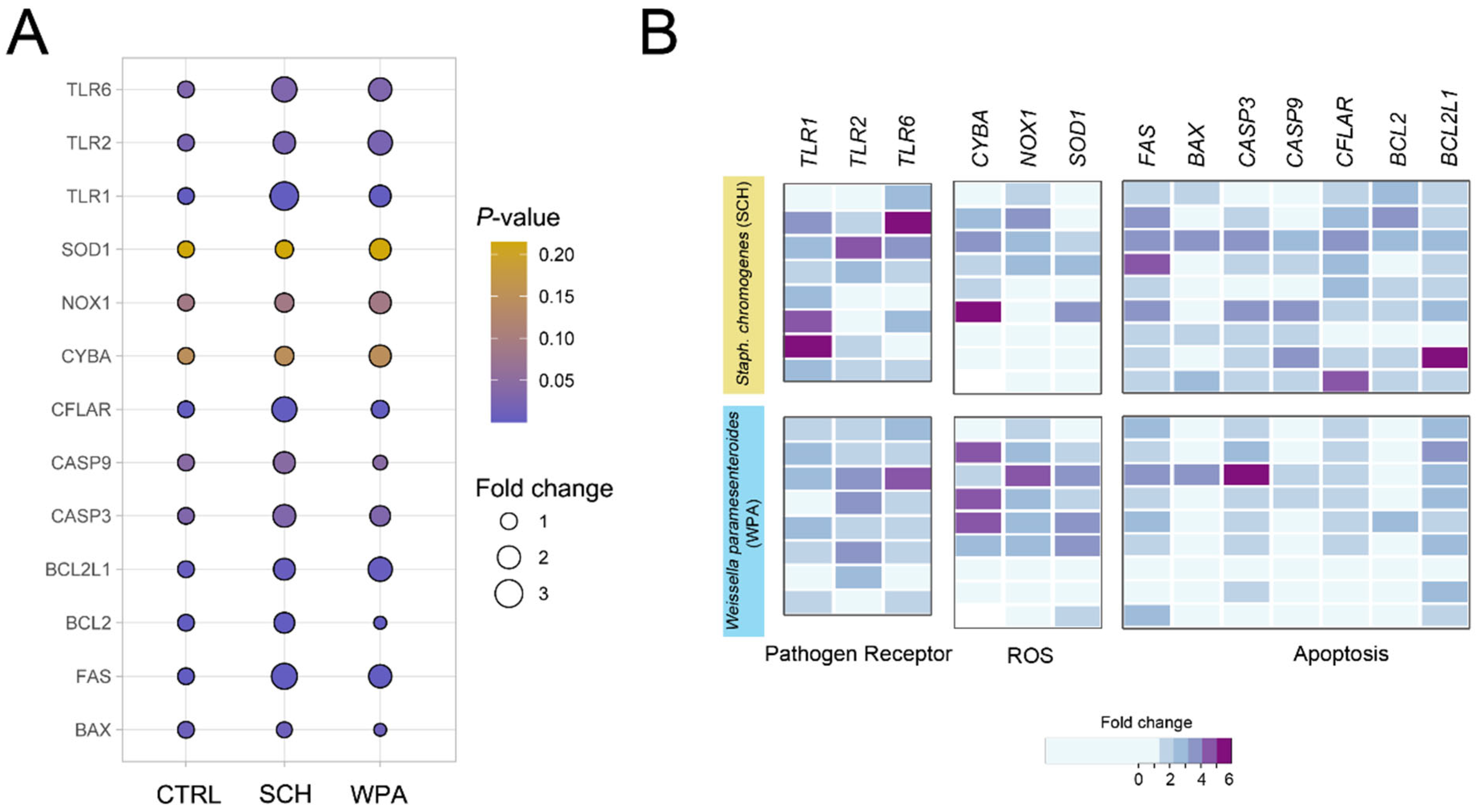
3.6. Minor Mastitis Bacteria Significantly Upregulated Gene Expression in Pathways Related to Bacterial Recognition, ROS Production, and Cellular Apoptosis Pathways
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morales-Ubaldo, A.L.; Rivero-Perez, N.; Valladares-Carranza, B.; Velázquez-Ordoñez, V.; Delgadillo-Ruiz, L.; Zaragoza-Bastida, A. Bovine mastitis, a worldwide impact disease: Prevalence, antimicrobial resistance, and viable alternative approaches. Vet. Anim. Sci. 2023, 21, 100306. [Google Scholar] [CrossRef] [PubMed]
- Kurban, D.; Roy, J.-P.; Kabera, F.; Fréchette, A.; Um, M.M.; Albaaj, A.; Rowe, S.; Godden, S.; Adkins, P.R.; Middleton, J.R. Diagnosing Intramammary infection: Meta-analysis and mapping review on frequency and udder health relevance of microorganism species isolated from bovine Milk samples. Animals 2022, 12, 3288. [Google Scholar] [CrossRef]
- Addis, M.F.; Locatelli, C.; Penati, M.; Poli, S.F.; Monistero, V.; Zingale, L.; Rota, N.; Gusmara, C.; Piccinini, R.; Moroni, P. Non-aureus staphylococci and mammaliicocci isolated from bovine milk in Italian dairy farms: A retrospective investigation. Vet. Res. Commun. 2024, 48, 547–554. [Google Scholar] [CrossRef]
- Freu, G.; Gioia, G.; Gross, B.; Biscarini, F.; Virkler, P.; Watters, R.; Addis, M.; Franklin-Guild, R.; Runyan, J.; Masroure, A. Frequency of non-aureus staphylococci and mammaliicocci species isolated from quarter clinical mastitis: A six-year retrospective study. J. Dairy Sci. 2023, in press. [Google Scholar] [CrossRef]
- Ruiz-Romero, R.A.; Vargas-Bello-Pérez, E. Non-aureus staphylococci and mammaliicocci as a cause of mastitis in domestic ruminants: Current knowledge, advances, biomedical applications, and future perspectives–A systematic review. Vet. Res. Commun. 2023, 47, 1067–1084. [Google Scholar] [CrossRef]
- Supré, K.; Haesebrouck, F.; Zadoks, R.; Vaneechoutte, M.; Piepers, S.; De Vliegher, S. Some coagulase-negative Staphylococcus species affect udder health more than others. J. Dairy Sci. 2011, 94, 2329–2340. [Google Scholar] [CrossRef]
- De Buck, J.; Ha, V.; Naushad, S.; Nobrega, D.B.; Luby, C.; Middleton, J.R.; De Vliegher, S.; Barkema, H.W. Non-aureus staphylococci and bovine udder health: Current understanding and knowledge gaps. Front. Vet. Sci. 2021, 8, 658031. [Google Scholar] [CrossRef]
- Zigo, F.; Farkašová, Z.; Výrostková, J.; Regecová, I.; Ondrašovičová, S.; Vargová, M.; Sasáková, N.; Pecka-Kielb, E.; Bursová, Š.; Kiss, D.S. Dairy cows’ udder pathogens and occurrence of virulence factors in Staphylococci. Animals 2022, 12, 470. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Shi, W.; Meng, N.; Zhao, Y.; Ding, X.; Li, Q. Antimicrobial resistance and virulence profiles of staphylococci isolated from clinical bovine mastitis. Front. Microbiol. 2023, 14, 1190790. [Google Scholar] [CrossRef]
- Hadrich, J.; Wolf, C.; Lombard, J.; Dolak, T. Estimating milk yield and value losses from increased somatic cell count on US dairy farms. J. Dairy Sci. 2018, 101, 3588–3596. [Google Scholar] [CrossRef]
- Isaac, P.; Bohl, L.P.; Breser, M.L.; Orellano, M.S.; Conesa, A.; Ferrero, M.A.; Porporatto, C. Commensal coagulase-negative Staphylococcus from the udder of healthy cows inhibits biofilm formation of mastitis-related pathogens. Vet. Microbiol. 2017, 207, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Diepers, A.-C.; Krömker, V.; Zinke, C.; Wente, N.; Pan, L.; Paulsen, K.; Paduch, J.-H. In vitro ability of lactic acid bacteria to inhibit mastitis-causing pathogens. Sustain. Chem. Pharm. 2017, 5, 84–92. [Google Scholar] [CrossRef]
- Pellegrino, M.S.; Frola, I.D.; Natanael, B.; Gobelli, D.; Nader-Macias, M.E.; Bogni, C.I. In vitro characterization of lactic acid bacteria isolated from bovine milk as potential probiotic strains to prevent bovine mastitis. Probiotics Antimicrob. Proteins 2019, 11, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Espeche, M.C.; Pellegrino, M.; Frola, I.; Larriestra, A.; Bogni, C.; Nader-Macías, M.F. Lactic acid bacteria from raw milk as potentially beneficial strains to prevent bovine mastitis. Anaerobe 2012, 18, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Srithanasuwan, A.; Tata, L.; Tananupak, W.; Jaraja, W.; Suriyasathaporn, W.; Chuammitri, P. Exploring the distinct immunological reactions of bovine neutrophils towards major and minor pathogens responsible for mastitis. Int. J. Vet. Sci. Med. 2023, 11, 106–120. [Google Scholar] [CrossRef] [PubMed]
- Blagitz, M.G.; Souza, F.N.; Batista, C.F.; Santos, B.P.; Parra, A.C.; Azevedo, L.F.F.; Della Libera, A.M. Expression of CD14 and Toll-like receptors 2 and 4 by milk neutrophils in bovine mammary glands infected with Corynebacterium bovis. Pesqui. Vet. Bras. 2015, 35, 1–5. [Google Scholar] [CrossRef]
- Benjamin, A.; Green, B.; Hayden, L.; Barlow, J.; Kerr, D. Cow-to-cow variation in fibroblast response to a Toll-like receptor 2/6 agonist and its relation to mastitis caused by intramammary challenge with Staphylococcus aureus. J. Dairy Sci. 2015, 98, 1836–1850. [Google Scholar] [CrossRef] [PubMed]
- Disbanchong, P.; Punmanee, W.; Srithanasuwan, A.; Pangprasit, N.; Wongsawan, K.; Suriyasathaporn, W.; Chuammitri, P. Immunomodulatory effects of herbal compounds Quercetin and Curcumin on cellular and molecular functions of bovine-milk-isolated neutrophils toward Streptococcus agalactiae infection. Animals 2021, 11, 3286. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Jia, Y.; Qian, Y.; Jiang, X.; Zhang, S.; Liu, B.; Cao, J.; Song, Y.; Mao, W. Staphylococcus aureus increases Prostaglandin E2 secretion in cow neutrophils by activating TLR2, TLR4, and NLRP3 inflammasome signaling pathways. Front. Microbiol. 2023, 14, 1163261. [Google Scholar] [CrossRef] [PubMed]
- Lippolis, J.D.; Reinhardt, T.A.; Goff, J.P.; Horst, R.L. Neutrophil extracellular trap formation by bovine neutrophils is not inhibited by milk. Vet. Immunol. Immunopathol. 2006, 113, 248–255. [Google Scholar] [CrossRef]
- Mektrirat, R.; Chuammitri, P.; Navathong, D.; Khumma, T.; Srithanasuwan, A.; Suriyasathaporn, W. Exploring the potential immunomodulatory effects of gallic acid on milk phagocytes in bovine mastitis caused by Staphylococcus aureus. Front. Vet. Sci. 2023, 10, 1255058. [Google Scholar] [CrossRef]
- Ma, F.; Yang, S.; Wang, G.; Zhou, M.; Zhang, J.; Deng, B.; Yin, W.; Wang, H.; Lu, Y.; Fan, H. Effect of multiplicity of infection on the evasion of neutrophil killing by Streptococcus agalactiae isolated from clinical mastitis bovine. Vet. Microbiol. 2022, 270, 109450. [Google Scholar] [CrossRef]
- Blagitz, M.G.; Souza, F.N.; Batista, C.F.; Azevedo, L.F.F.; Benites, N.R.; Melville, P.A.; Diniz, S.A.; Silva, M.X.; Haddad, J.P.A.; Heinnemann, M.B. The neutrophil function and lymphocyte profile of milk from bovine mammary glands infected with Streptococcus dysgalactiae. J. Dairy Res. 2015, 82, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Gondaira, S.; Nishi, K.; Fujiki, J.; Iwano, H.; Watanabe, R.; Eguchi, A.; Hirano, Y.; Higuchi, H.; Nagahata, H. Innate immune response in bovine neutrophils stimulated with Mycoplasma bovis. Vet. Res. 2021, 52, 58. [Google Scholar] [CrossRef] [PubMed]
- Conejeros, I.; Gibson, A.J.; Werling, D.; Muñoz-Caro, T.; Hermosilla, C.; Taubert, A.; Burgos, R.A. Effect of the synthetic Toll-like receptor ligands LPS, Pam3CSK4, HKLM and FSL-1 in the function of bovine polymorphonuclear neutrophils. Dev. Comp. Immunol. 2015, 52, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Worku, M.; Rehrah, D.; Ismail, H.D.; Asiamah, E.; Adjei-Fremah, S. A review of the Neutrophil Extracellular Traps (NETs) from cow, sheep and goat models. Int. J. Mol. Sci. 2021, 22, 8046. [Google Scholar] [CrossRef]
- Conejeros, I.; Patterson, R.; Burgos, R.; Hermosilla, C.; Werling, D. Induction of reactive oxygen species in bovine neutrophils is CD11b, but not dectin-1-dependent. Vet. Immunol. Immunopathol. 2011, 139, 308–312. [Google Scholar] [CrossRef]
- Maurić Maljković, M.; Vlahek, I.; Piplica, A.; Ekert Kabalin, A.; Sušić, V.; Stevanović, V. Prospects of toll-like receptors in dairy cattle breeding. Anim. Genet. 2023, 54, 425–434. [Google Scholar] [CrossRef]
- Elmaghraby, M.; El-Nahas, A.; Fathala, M.; Sahwan, F.; El-Dien, M.T. Association of Toll-like receptors 2 and 6 polymorphism with clinical mastitis and production traits in Holstein cattle. Iran. J. Vet. Res. 2018, 19, 202–207. [Google Scholar]
- Khan, M.Z.; Wang, J.; Ma, Y.; Chen, T.; Ma, M.; Ullah, Q.; Khan, I.M.; Khan, A.; Cao, Z.; Liu, S. Genetic polymorphisms in immune-and inflammation-associated genes and their association with bovine mastitis resistance/susceptibility. Front. Immunol. 2023, 14, 1082144. [Google Scholar] [CrossRef]
- Bai, X.; Wang, X.; Lin, T.; Dong, W.; Gao, Y.; Ji, P.; Zhang, Y.; Zhao, X.; Zhang, Q. Toll-like receptor 2 is associated with the immune response, apoptosis, and angiogenesis in the mammary glands of dairy cows with clinical mastitis. Int. J. Mol. Sci. 2022, 23, 10717. [Google Scholar] [CrossRef] [PubMed]
- Okusawa, T.; Fujita, M.; Nakamura, J.-i.; Into, T.; Yasuda, M.; Yoshimura, A.; Hara, Y.; Hasebe, A.; Golenbock, D.T.; Morita, M. Relationship between structures and biological activities of mycoplasmal diacylated lipopeptides and their recognition by Toll-like receptors 2 and 6. Infect. Immun. 2004, 72, 1657–1665. [Google Scholar] [CrossRef] [PubMed]
- Souza, R.M.; Souza, F.N.; Batista, C.F.; Piepers, S.; De Visscher, A.; Santos, K.R.; Molinari, P.C.; Ferronatto, J.A.; da Cunha, A.F.; Blagitz, M.G. Distinct behavior of bovine-associated staphylococci species in their ability to resist phagocytosis and trigger respiratory burst activity by blood and milk polymorphonuclear leukocytes in dairy cows. J. Dairy Sci. 2022, 105, 1625–1637. [Google Scholar] [CrossRef] [PubMed]
- Bassel, L.L.; Caswell, J.L. Bovine neutrophils in health and disease. Cell Tissue Res. 2018, 371, 617–637. [Google Scholar] [CrossRef] [PubMed]
- Kampen, A.H.; Tollersrud, T.; Lund, A. Staphylococcus aureus capsular polysaccharide types 5 and 8 reduce killing by bovine neutrophils in vitro. Infect. Immun. 2005, 73, 1578–1583. [Google Scholar] [CrossRef] [PubMed]
- Toledo-Silva, B.; de Souza, F.N.; Mertens, K.; Piepers, S.; Haesebrouck, F.; De Vliegher, S. Bovine-associated non-aureus staphylococci suppress Staphylococcus aureus biofilm dispersal in vitro yet not through agr regulation. Vet. Res. 2021, 52, 114. [Google Scholar] [CrossRef]
- Tremblay, Y.D.; Caron, V.; Blondeau, A.; Messier, S.; Jacques, M. Biofilm formation by coagulase-negative staphylococci: Impact on the efficacy of antimicrobials and disinfectants commonly used on dairy farms. Vet. Microbiol. 2014, 172, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Goetz, C.; Tremblay, Y.D.; Lamarche, D.; Blondeau, A.; Gaudreau, A.M.; Labrie, J.; Malouin, F.; Jacques, M. Coagulase-negative staphylococci species affect biofilm formation of other coagulase-negative and coagulase-positive staphylococci. J. Dairy Sci. 2017, 100, 6454–6464. [Google Scholar] [CrossRef]
- Kubinak, J.L.; Round, J.L. Toll-like receptors promote mutually beneficial commensal-host interactions. PLoS Pathog. 2012, 8, e1002785. [Google Scholar] [CrossRef]
- Islam, M.A.; Takagi, M.; Fukuyama, K.; Komatsu, R.; Albarracin, L.; Nochi, T.; Suda, Y.; Ikeda-Ohtsubo, W.; Rutten, V.; Eden, W.v. Transcriptome analysis of the inflammatory responses of bovine mammary epithelial cells: Exploring immunomodulatory target genes for bovine mastitis. Pathogens 2020, 9, 200. [Google Scholar] [CrossRef]
- Valckenier, D.; Piepers, S.; Schukken, Y.H.; De Visscher, A.; Boyen, F.; Haesebrouck, F.; De Vliegher, S. Longitudinal study on the effects of intramammary infection with non-aureus staphylococci on udder health and milk production in dairy heifers. J. Dairy Sci. 2021, 104, 899–914. [Google Scholar] [CrossRef] [PubMed]
- Mulcahy, M.E.; O’Brien, E.C.; O’Keeffe, K.M.; Vozza, E.G.; Leddy, N.; McLoughlin, R.M. Manipulation of autophagy and apoptosis facilitates intracellular survival of Staphylococcus aureus in human neutrophils. Front. Immunol. 2020, 11, 565545. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | CTRL | SCH | WPA | p Value | Test |
|---|---|---|---|---|---|
| TLR1 | 1.00 ± 0.00 b | 3.22 ± 0.54 a | 1.79 ± 0.29 b | 0.001 | ANOVA |
| TLR2 | 1.00 ± 0.00 b | 1.89 ± 0.44 ab | 2.31 ± 0.32 a | 0.023 | ANOVA |
| TLR6 | 1.00 ± 0.00 b | 2.45 ± 0.50 a | 2.09 ± 0.39 ab | 0.030 | ANOVA |
| CYBA | 1.00 ± 0.00 a | 1.89 ± 0.54 a | 2.19 ± 0.51 a | 0.148 | ANOVA |
| NOX1 | 1.00 ± 0.00 a | 1.36 ± 0.32 a | 1.84 ± 0.30 a | 0.088 | ANOVA |
| SOD1 | 1.00 ± 0.00 a | 1.21 ± 0.32 a | 1.78 ± 0.36 a | 0.214 | Kruskal–Wallis |
| FAS | 1.00 ± 0.00 b | 2.63 ± 0.47 a | 2.07 ± 0.29 ab | 0.0005 | Kruskal–Wallis |
| BAX | 1.00 ± 0.00 a | 0.89 ± 0.27 a | 0.54 ± 0.14 b | 0.0097 | Kruskal–Wallis |
| CFLAR | 1.00 ± 0.00 b | 2.44 ± 0.41 a | 1.17 ± 0.18 b | 0.0011 | ANOVA |
| CASP3 | 1.00 ± 0.00 b | 1.98 ± 0.36 a | 1.58 ± 0.24 ab | 0.0318 | ANOVA |
| CASP9 | 1.00 ± 0.00 ab | 1.81 ± 0.44 a | 0.74 ± 0.23 b | 0.0444 | Kruskal–Wallis |
| BCL2 | 1.00 ± 0.00 b | 1.61 ± 0.21 a | 0.56 ± 0.11 b | <0.0001 | ANOVA |
| BCL2L1 | 1.00 ± 0.00 b | 1.78 ± 0.30 ab | 2.29 ± 0.31 ab | 0.0035 | ANOVA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Srithanasuwan, A.; Pangprasit, N.; Mektrirat, R.; Suriyasathaporn, W.; Chuammitri, P. Divergent Immune Responses to Minor Bovine Mastitis-Causing Pathogens. Vet. Sci. 2024, 11, 262. https://doi.org/10.3390/vetsci11060262
Srithanasuwan A, Pangprasit N, Mektrirat R, Suriyasathaporn W, Chuammitri P. Divergent Immune Responses to Minor Bovine Mastitis-Causing Pathogens. Veterinary Sciences. 2024; 11(6):262. https://doi.org/10.3390/vetsci11060262
Chicago/Turabian StyleSrithanasuwan, Anyaphat, Noppason Pangprasit, Raktham Mektrirat, Witaya Suriyasathaporn, and Phongsakorn Chuammitri. 2024. "Divergent Immune Responses to Minor Bovine Mastitis-Causing Pathogens" Veterinary Sciences 11, no. 6: 262. https://doi.org/10.3390/vetsci11060262
APA StyleSrithanasuwan, A., Pangprasit, N., Mektrirat, R., Suriyasathaporn, W., & Chuammitri, P. (2024). Divergent Immune Responses to Minor Bovine Mastitis-Causing Pathogens. Veterinary Sciences, 11(6), 262. https://doi.org/10.3390/vetsci11060262