

Comparative Analysis of Antibiotic Resistance and Biofilm Characteristics of Two Major Enterococcus Species from Poultry Slaughterhouses in South Korea
 , , , ,
, , , ,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates and Culture Media
2.2. Antibiotic Susceptibility Test
2.3. Microtiter Plate Biofilm Assay
2.4. Polymerase Chain Reaction
2.5. Statistical Analysis
3. Results
3.1. Identification of Enterococcus spp.
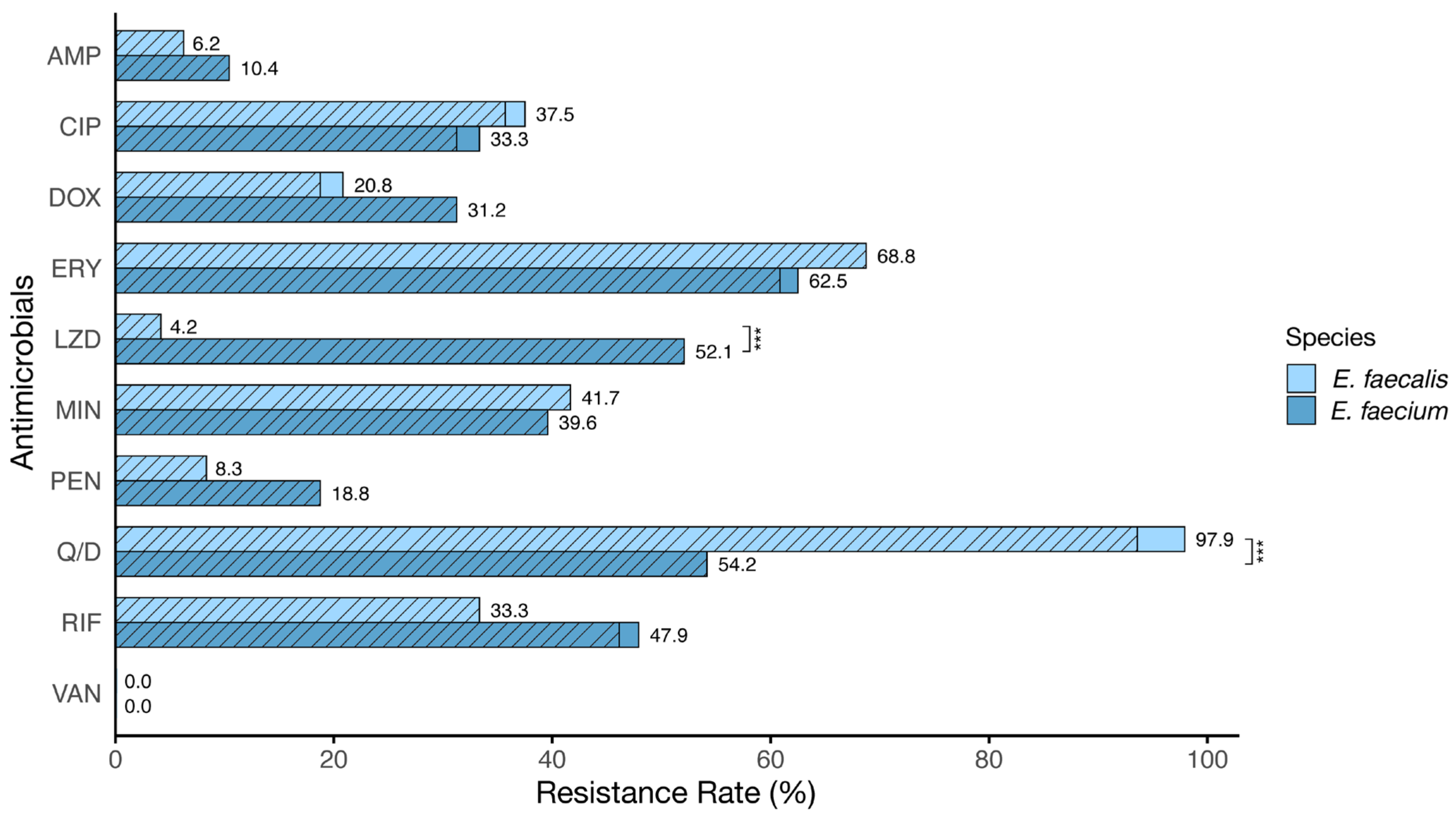
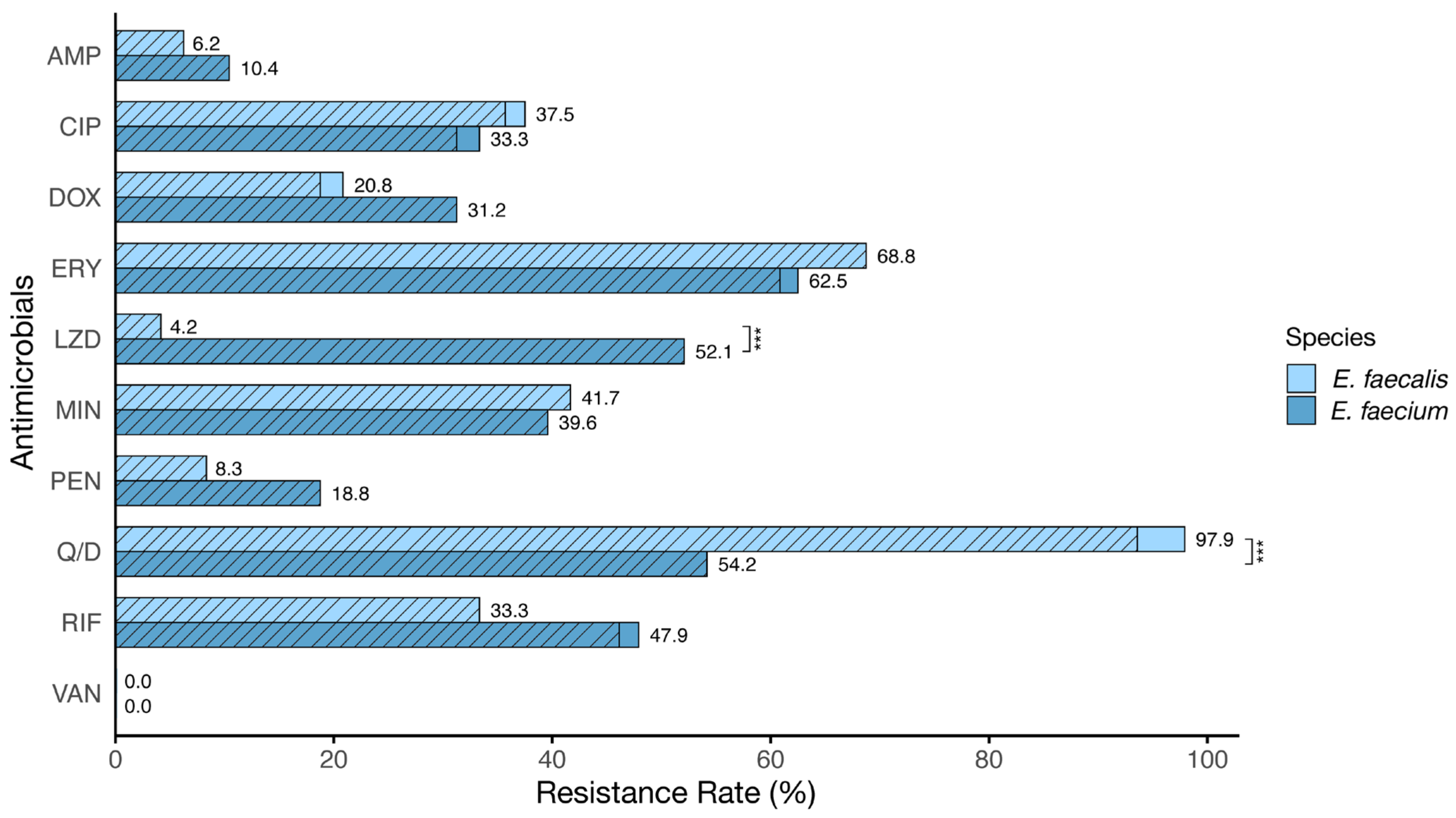
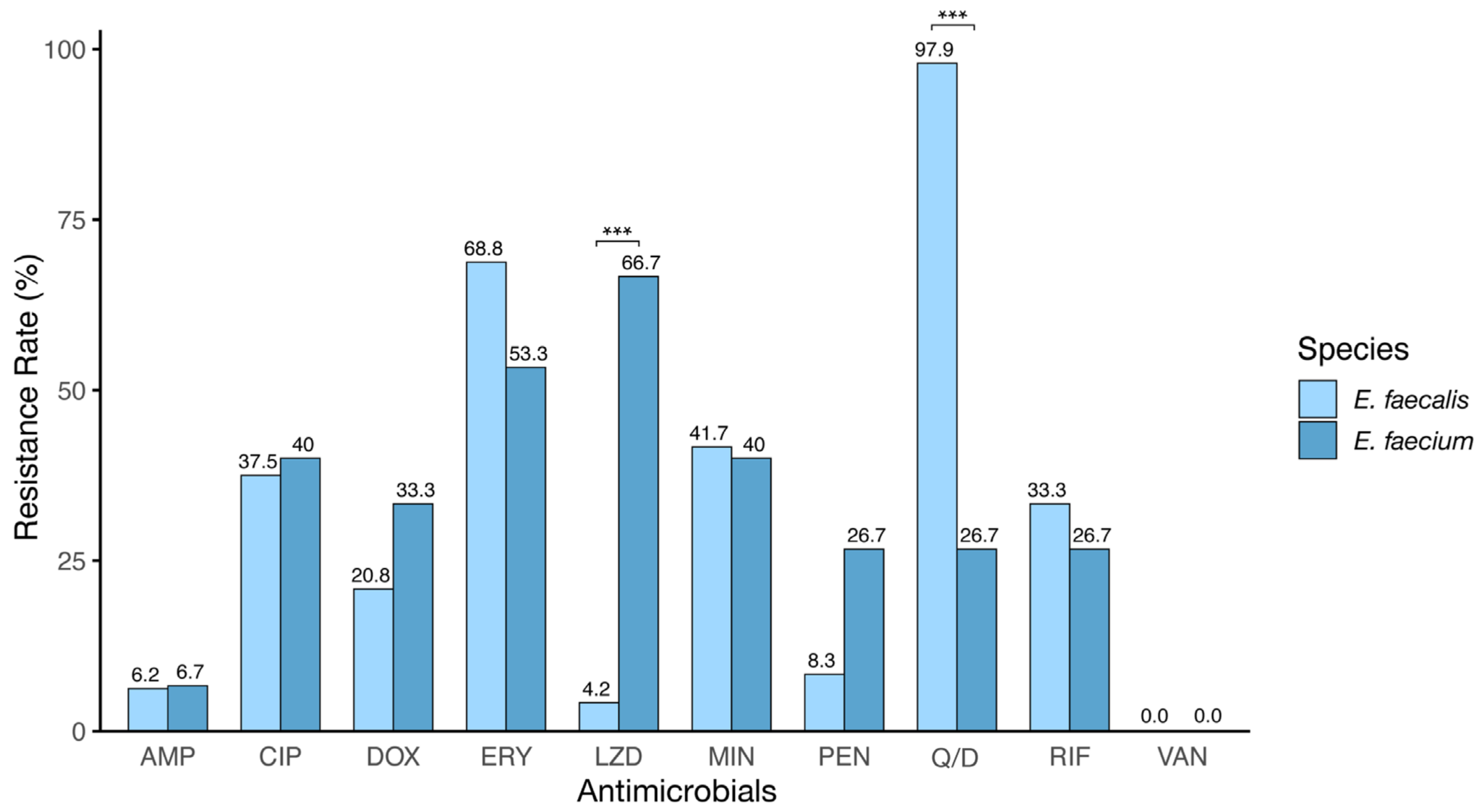
3.2. Antibiotic Susceptibility Profile
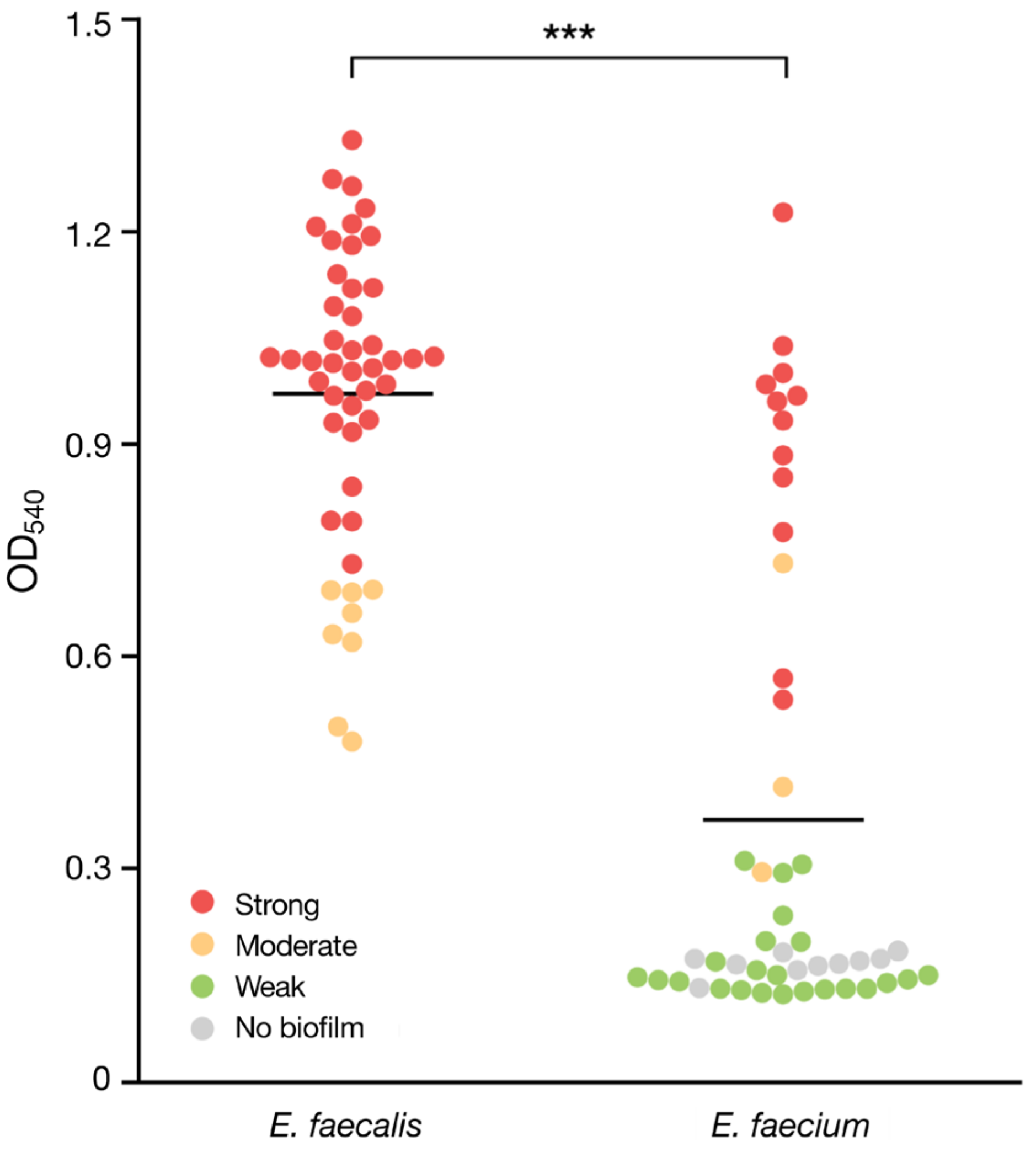
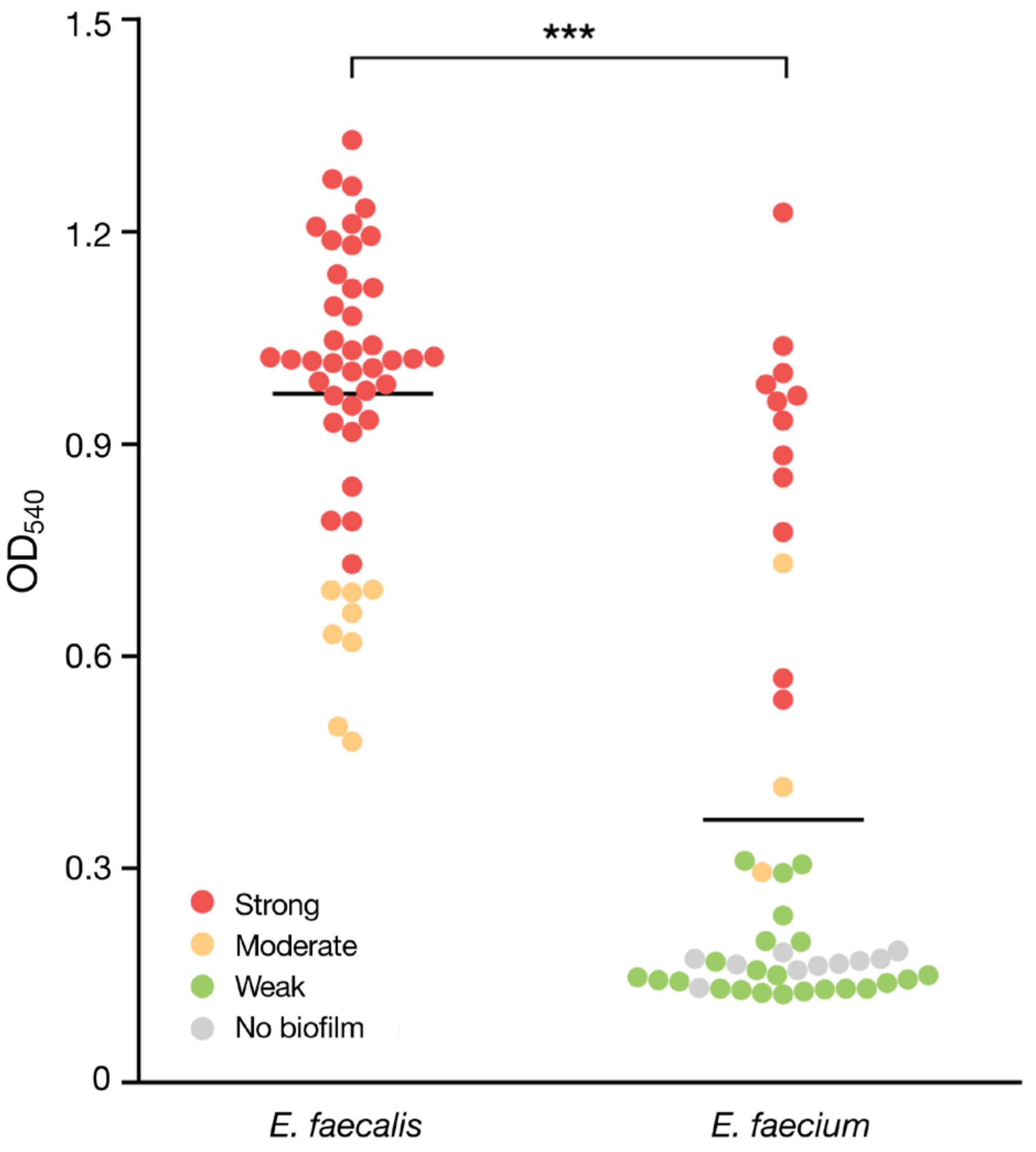
3.3. Biofilm Formation
3.4. Screening Genes Involved in Biofilm Formation
3.5. Antibiotic Resistance across Biofilm Strength
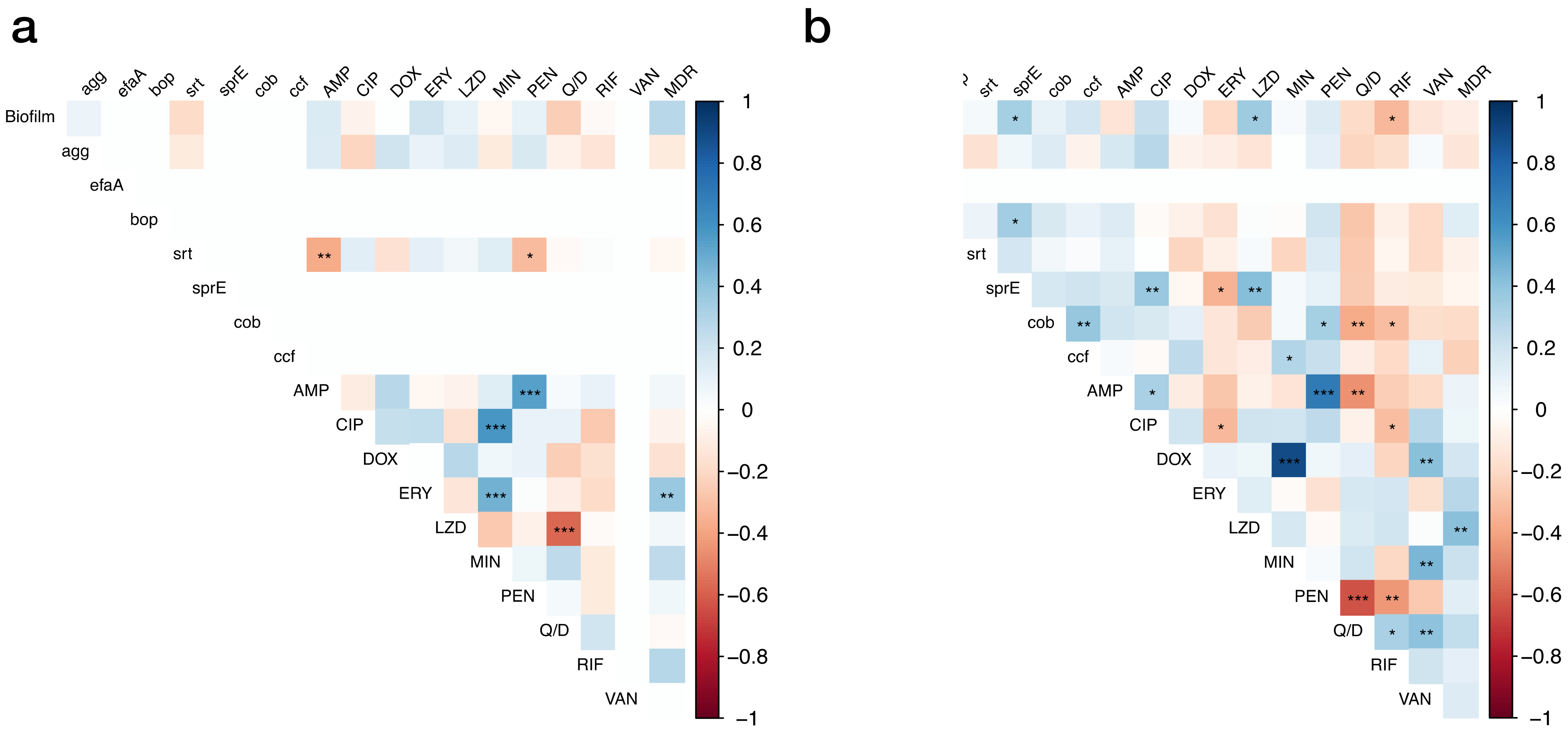
3.6. Correlations between Studied Factors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mohamed, J.A.; Huang, D.B. Biofilm Formation by Enterococci. J. Med. Microbiol. 2007, 56, 1581–1588. [Google Scholar] [CrossRef] [PubMed]
- Manson, A.L.; Van Tyne, D.; Straub, T.J.; Clock, S.; Crupain, M.; Rangan, U.; Gilmore, M.S.; Earl, A.M. Chicken Meat-Associated Enterococci: Influence of Agricultural Antibiotic Use and Connection to the Clinic. Appl. Environ. Microbiol. 2019, 85, e01559-19. [Google Scholar] [CrossRef] [PubMed]
- Murray, B.E. The Life and Times of the Enterococcus. Clin. Microbiol. Rev. 1990, 3, 46–65. [Google Scholar] [CrossRef] [PubMed]
- Galié, S.; García-Gutiérrez, C.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Biofilms in the Food Industry: Health Aspects and Control Methods. Front. Microbiol. 2018, 9, 315815. [Google Scholar] [CrossRef] [PubMed]
- Malidareh, E.B.; Ahanjan, M.; Marghmalek, S.A.; Goli, H.R. Dissemination of Quinupristin-Dalfopristin and Linezolid Resistance Genes among Hospital Environmental and Healthy Volunteer Fecal Isolates of Enterococcus faecalis and Enterococcus faecium. Mol. Biol. Rep. 2022, 49, 7929–7937. [Google Scholar] [CrossRef] [PubMed]
- Hollenbeck, B.L.; Rice, L.B. Intrinsic and Acquired Resistance Mechanisms in Enterococcus. Virulence 2012, 3, 421–569. [Google Scholar] [CrossRef] [PubMed]
- Rudy, M.; Zientara, M.; Bek, T.; Martirosian, G. Occurrence of Antibiotic Resistant Enterococci in Clinical Specimens from a Pediatric Hospital. Pol. J. Microbiol. 2005, 54, 77–80. [Google Scholar] [PubMed]
- Aslangul, E.; Massias, L.; Meulemans, A.; Chau, F.; Andremont, A.; Courvalin, P.; Fantin, B.; Ruimy, R. Acquired Gentamicin Resistance by Permeability Impairment in Enterococcus faecalis. Antimicrob. Agents Chemother. 2006, 50, 3615–3621. [Google Scholar] [CrossRef] [PubMed]
- Abat, C.; Raoult, D.; Rolain, J.M. Low Level of Resistance in Enterococci Isolated in Four Hospitals, Marseille, France. Microb. Drug Resis 2016, 22, 218–222. [Google Scholar] [CrossRef] [PubMed]
- Duez, C.; Zorzi, W. The Penicillin Resistance of Enterococcus faecalis JH2-2r Results from an Overproduction of the Low-Affinity Penicillin-Binding Protein PBP4 and Does Not Involve a psr-like Gene. Microbiology 2001, 147, 2561–2569. [Google Scholar] [CrossRef]
- Murray, B.E. Vancomycin-Resistant Enterococci. Am. J. Med. 1997, 102, 284–293. [Google Scholar] [CrossRef] [PubMed]
- Ch’ng, J.H.; Chong, K.K.L.; Lam, L.N.; Wong, J.J.; Kline, K.A. Biofilm-Associated Infection by Enterococci. Nat. Rev. Microbiol. 2019, 17, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Giraffa, G. Enterococci from Foods. FEMS Microbiol. Rev. 2002, 26, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.; Santos, V.; Fernandes, A.; Bernardo, F.; Vilela, C.L. Antimicrobial Resistance and in vitro Biofilm-Forming Ability of Enterococci from Intensive and Extensive Farming Broilers. Poult. Sci. 2010, 89, 1065–1069. [Google Scholar] [CrossRef] [PubMed]
- Castaño-Arriba, A.; González-Machado, C.; Igrejas, G.; Poeta, P.; Alonso-Calleja, C.; Capita, R. Antibiotic Resistance and Biofilm-Forming Ability in Enterococcal Isolates from Red Meat and Poultry Preparations. Pathogens 2020, 9, 1021. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.R.; English, L.L.; Carter, P.J.; Proescholdt, T.; Lee, K.Y.; Wagner, D.D.; White, D.G. Prevalence and Antimicrobial Resistance of Enterococcus Species Isolated from Retail Meats. Appl. Environ. Microbiol. 2003, 69, 7153–7160. [Google Scholar] [CrossRef] [PubMed]
- Novais, C.; Freitas, A.R.; Silveira, E.; Antunes, P.; Silva, R.; Coque, T.M.; Peixe, L. Spread of Multidrug-Resistant Enterococcus to Animals and Humans: An Underestimated Role for the Pig Farm Environment. J. Antimicrob. Chemother. 2013, 68, 2746–2754. [Google Scholar] [CrossRef] [PubMed]
- Aslam, M.; Diarra, M.S.; Checkley, S.; Bohaychuk, V.; Masson, L. Characterization of Antimicrobial Resistance and Virulence Genes in Enterococcus spp. Isolated from Retail Meats in Alberta, Canada. Int. J. Food Microbiol. 2012, 156, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An Emergent Form of Bacterial Life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Șchiopu, P.; Toc, D.A.; Colosi, I.A.; Costache, C.; Ruospo, G.; Berar, G.; Gălbău, Ș.G.; Ghilea, A.C.; Botan, A.; Pană, A.G.; et al. An Overview of the Factors Involved in Biofilm Production by the Enterococcus Genus. Int. J. Mol. Sci. 2023, 24, 11577. [Google Scholar] [CrossRef] [PubMed]
- Sußmuth, S.D.; Muscholl-Silberhorn, A.; Wirth, R.; Susa, M.; Marre, R.; Rozdzinski, E. Aggregation Substance Promotes Adherence, Phagocytosis, and Intracellular Survival of Enterococcus faecalis within Human Macrophages and Suppresses Respiratory Burst. Infect. Immun. 2000, 68, 4900–4906. [Google Scholar] [CrossRef] [PubMed]
- Hashem, Y.A.; Amin, H.M.; Essam, T.M.; Yassin, A.S.; Aziz, R.K. Biofilm Formation in Enterococci: Genotype-Phenotype Correlations and Inhibition by Vancomycin. Sci. Rep. 2017, 7, 5733. [Google Scholar] [PubMed]
- Nallapareddy, S.R.; Singh, K.V.; Sillanpää, J.; Garsin, D.A.; Höök, M.; Erlandsen, S.L.; Murray, B.E. Endocarditis and Biofilm-Associated Pili of Enterococcus faecalis. J. Clin. Investig. 2006, 116, 2799–2807. [Google Scholar] [CrossRef] [PubMed]
- Hufnagel, M.; Koch, S.; Creti, R.; Baldassarri, L.; Huebner, J. A Putative Sugar-Binding Transcriptional Regulator in a Novel Gene Locus in Enterococcus faecalis Contributes to Production of Biofilm and Prolonged Bacteremia in Mice. J. Infect. Dis. 2004, 189, 420–430. [Google Scholar] [PubMed]
- Nakayama, J.; Abe, Y.; Ono, Y.; Isogai, A.; Suzuki, A. Isolation and Structure of the Enterococcus Faecalis Sex Pheromone, COB1, That Induces Conjugal Transfer of the Hemolysin / Bacteriocin Plasmids, POB1 and PYI1. Biosci. Biotechnol. Biochem. 1995, 59, 703–705. [Google Scholar] [CrossRef] [PubMed]
- Antiporta, M.H.; Dunny, G.M. Ccfa, the Genetic Determinant for the CCF10 Peptide Pheromone in Enterococcus faecalis OG1RF. J. Bacteriol. 2002, 184, 1155–1162. [Google Scholar] [PubMed]
- Thomas, V.C.; Thurlow, L.R.; Boyle, D.; Hancock, L.E. Regulation of Autolysis-Dependent Extracellular DNA Release by Enterococcus faecalis Extracellular Proteases Influences Biofilm Development. J. Bacteriol. 2008, 190, 5690–5698. [Google Scholar] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals, 5th ed.; VET01S; Clinical and Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2021. [Google Scholar]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Christensen, G.D.; Simpson, W.A.; Younger, J.J.; Baddour, L.M.; Barrett, F.F.; Melton, D.M.; Beachey, E.H. Adherence of Coagulase-Negative Staphylococci to Plastic Tissue Plates: A Quantitative Model for the Adherence of Staphylococci to Medical Devices. J. Clin. Microbiol. 1985, 22, 996–1006. [Google Scholar] [PubMed]
- Stepanović, S.; Vuković, D.; Hola, V.; Di Bonaventura, G.; Djukić, S.; Ćirković, I.; Ruzicka, F. Quantification of Biofilm in Microtiter Plates: Overview of Testing Conditions and Practical Recommendations for Assessment of Biofilm Production by Staphylococci. APMIS 2007, 115, 891–899. [Google Scholar]
- Layton, B.A.; Walters, S.P.; Lam, L.H.; Boehm, A.B. Enterococcus Species Distribution among Human and Animal Hosts Using Multiplex PCR. J. Appl. Microbiol. 2010, 109, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Animal and Plant Quarantine Agency; Ministry of Agriculture Food and Rural Affairs; National Institute of Food and Drug Safety Evaluation. National Antibiotic Usage and Resistance Monitoring 2022: Animals, Livestock Products; Antibiotic resistance of indicator bacteria; Animal and Plant Quarantine Agency (APQA): Gimcheon, Gyeonsangbuk-do, Republic of Korea, 2023; Chapter 2. [Google Scholar]
- Yu, L.; Liu, Y.; Liu, M.; Li, Z.; Li, L.; Wang, F. Research Note: Molecular Characterization of Antimicrobial Resistance and Virulence Gene Analysis of Enterococcus faecalis in Poultry in Tai’an, China. Poult. Sci. 2022, 101, 101763. [Google Scholar] [CrossRef] [PubMed]
- Mwikuma, G.; Kainga, H.; Kallu, S.A.; Nakajima, C.; Suzuki, Y.; Hang’ombe, B.M. Determination of the Prevalence and Antimicrobial Resistance of Enterococcus faecalis and Enterococcus faecium Associated with Poultry in Four Districts in Zambia. Antibiotics 2023, 12, 657. [Google Scholar] [CrossRef]
- Kim, Y.B.; Yoon, S.; Seo, K.W.; Shim, J.B.; Noh, E.B.; Lee, Y.J. Detection of Linezolid-Resistant Enterococcus faecium Isolates from the Layer Operation System in Korea. Microb. Drug Resist. 2021, 27, 1443–1449. [Google Scholar] [CrossRef] [PubMed]
- Olsen, R.H.; Schønheyder, H.C.; Christensen, H.; Bisgaard, M. Enterococcus faecalis of Human and Poultry Origin Share Virulence Genes Supporting the Zoonotic Potential of E. faecalis. Zoonoses Public Health 2012, 59, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Woźniak-Biel, A.; Bugla-Płoskońska, G.; Burdzy, J.; Korzekwa, K.; Ploch, S.; Wieliczko, A. Antimicrobial Resistance and Biofilm Formation in Enterococcus spp. Isolated from Humans and Turkeys in Poland. Microb. Drug Resist. 2019, 25, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Park, J.H.; Seo, K.H. Comparison of the Loads and Antibiotic-Resistance Profiles of Enterococcus Species from Conventional and Organic Chicken Carcasses in South Korea. Poult. Sci. 2018, 97, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Alzahrani, O.M.; Fayez, M.; Alswat, A.S.; Alkafafy, M.; Mahmoud, S.F.; Al-Marri, T.; Almuslem, A.; Ashfaq, H.; Yusuf, S. Antimicrobial Resistance, Biofilm Formation, and Virulence Genes in Enterococcus Species from Small Backyard Chicken Flocks. Antibiotics 2022, 11, 380. [Google Scholar] [CrossRef] [PubMed]
- Stępień-Pyśniak, D.; Hauschild, T.; Kosikowska, U.; Dec, M.; Urban-Chmiel, R. Biofilm Formation Capacity and Presence of Virulence Factors among Commensal Enterococcus spp. from Wild Birds. Sci. Rep. 2019, 9, 11204. [Google Scholar] [CrossRef] [PubMed]
- Di Rosa, R.; Creti, R.; Venditti, M.; D’Amelio, R.; Arciola, C.; Montanaro, L.; Baldassarri, L. Relationship between Biofilm Formation, the Enterococcal Surface Protein (Esp) and Gelatinase in Clinical Isolates of Enterococcus faecalis and Enterococcus faecium. FEMS Microbiol. Lett. 2006, 256, 145–150. [Google Scholar] [PubMed]
- Popović, N.; Dinić, M.; Tolinacki, M.; Mihajlović, S.; Terzić-Vidojević, A.; Bojić, S.; Djokić, J.; Golić, N.; Veljović, K. New Insight into Biofilm Formation Ability, the Presence of Virulence Genes and Probiotic Potential of Enterococcus sp. Dairy Isolates. Front. Microbiol. 2018, 9, 78. [Google Scholar] [CrossRef] [PubMed]
- Martín, M.; Gutiérrez, J.; Criado, R.; Herranz, C.; Cintas, L.M.; Hernández, P.E. Genes Encoding Bacteriocins and Their Expression and Potential Virulence Factors of Enterococci Isolated from Wood Pigeons (Columba palumbus). J. Food Prot. 2006, 69, 520–531. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotic Class | Antibiotics | Classification | |
|---|---|---|---|
| WHO | WOAH | ||
| Penicillins | Ampicillin | CIA | VCIA |
| Penicillin | HIA | VCIA | |
| Quinolones | Ciprofloxacin | CIA | VCIA |
| Tetracyclines | Doxycycline | HIA | VCIA |
| Minocycline | HIA | - | |
| Macrolides | Erythromycin | CIA | VCIA |
| Oxazolidinones | Linezolid | CIA | - |
| Streptogramins | Quinupristin/Dalfopristin | HIA | - |
| Ansamycins | Rifampicin | CIA | VHIA |
| Glycopeptides | Vancomycin | CIA | - |
| Genes | Primer Sequence (5′-3′) | Annealing (°C) | Product Size (bp) | Reference | |
|---|---|---|---|---|---|
| Enterococcus 16S rRNA | F R | TCAACCGGGGAGGGT ATTACTAGCGATTCCGG | 55 | 733 | [32] |
| E. faecalis | F R | ACTTATGTGACTAACTTAACC TAATGGTGAATCTTGGTTTGG | 55 | 214 | [32] |
| E. faecium | F R | ACAATAGAAGAATTATTATCTG CGGCTGCTTTTTTGAATTCTTCT | 55 | 360 | [32] |
| agg | F R | TCTTGGACACGACCCATGAT AGAAAGAACATCACCACGAGC | 58 | 413 | [22] |
| bop | F R | GATCGTCTTCGCCATAGTAGG ATACACAACAGCCCTTGGCT | 58 | 312 | [22] |
| ccf | F R | GGGAATTGAGTAGTGAAGAAG AGCCGCTAAAATCGGTAAAAT | 5 | 543 | [22] |
| cob | F R | GCTTTGTTTGCTGAATGTTCC GACAACTGATGAGGTGCTAG | 55 | 385 | [22] |
| efaA | F R | GACAGACCCTCACGAATATG CCAGTTCATCATGCTGTAGTA | 52 | 706 | [22] |
| sprE | F R | CTGAGGACAGAAGACAAGAAG GGTTTTTCTCACCTGGATAG | 55 | 432 | [22] |
| srt | F R | GTATCCTTTTGTTAGCGATGC TGTCCTCGAACTAATAACCGA | 55 | 612 | [22] |
| Biofilm Strength | agg | efaA | bop | srt | sprE | cob | ccf | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| + (%) | − (%) | + (%) | − (%) | + (%) | − (%) | + (%) | − (%) | + (%) | − (%) | + (%) | − (%) | + (%) | − (%) | |
| Moderate | 6 (12.5) | 2 (4.2) | 0 | 8 (16.7) | 8 (16.7) | 0 | 8 (16.7) | 0 | 8 (16.7) | 0 | 8 (16.7) | 0 | 8 (16.7) | 0 |
| Strong | 31 (64.6) | 9 (18.8) | 0 | 40 (83.3) | 40 (83.3) | 0 | 38 (79.2) | 2 (4.2) | 40 (83.3) | 0 | 40 (83.3) | 0 | 40 (83.3) | 0 |
| Total | 37 (77.1) | 11 (22.9) | 0 | 48 (100) | 48 (100) | 0 | 46 (95.8) | 2 (4.2) | 48 (100) | 0 | 48 (100) | 0 | 48 (100) | 0 |
| Biofilm Strength | agg | efaA | bop | srt | sprE | cob | ccf | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| + (%) | − (%) | + (%) | − (%) | + (%) | − (%) | + (%) | − (%) | + (%) | − (%) | + (%) | − (%) | + (%) | − (%) | |
| No biofilm | 9 (18.8) | 1 (2.1) | 0 | 10 (20.8) | 8 (16.7) | 2 (4.2) | 9 (18.8) | 1 (2.1) | 6 (12.5) | 4 (8.3) | 1 (2.1) | 9 (18.8) | 5 (10.4) | 5 (10.4) |
| Weak | 17 (35.4) | 6 (12.5) | 0 | 23 (47.9) | 20 (41.7) | 3 (6.3) | 20 (41.7) | 3 (6.3) | 9 (18.8) | 14 (29.2) | 8 (16.7) | 15 (31.3) | 10 (20.8) | 13 (27.1) |
| Moderate | 2 (4.2) | 1 (2.1) | 0 | 3 (6.3) | 3 (6.3) | 0 | 3 (6.3) | 0 | 3 (6.3) | 0 | 2 (4.2) | 1 (2.1) | 3 (6.3) | 0 |
| Strong | 10 (20.8) | 2 (4.2) | 0 | 12 (25) | 10 (20.8) | 2 (4.2) | 12 (25) | 0 | 9 (18.8) | 3 (6.3) | 5 (10.4) | 7 (14.6) | 8 (16.7) | 4 (8.3) |
| Total | 38 (79.2) | 10 (20.8) | 0 | 48 (100) | 41 (85.4) | 7 (14.6) | 44 (91.7) | 4 (8.3) | 27 (56.3) | 21 (43.8) | 16 (33.3) | 32 (66.7) | 26 (54.2) | 22 (45.8) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Son, Y.; Jin, Y.B.; Cho, E.-J.; Park, A.R.; Flores, R.A.; Nguyen, B.T.; Lee, S.Y.; Altanzul, B.; Park, K.I.; Min, W.; et al. Comparative Analysis of Antibiotic Resistance and Biofilm Characteristics of Two Major Enterococcus Species from Poultry Slaughterhouses in South Korea. Vet. Sci. 2024, 11, 180. https://doi.org/10.3390/vetsci11040180
Son Y, Jin YB, Cho E-J, Park AR, Flores RA, Nguyen BT, Lee SY, Altanzul B, Park KI, Min W, et al. Comparative Analysis of Antibiotic Resistance and Biofilm Characteristics of Two Major Enterococcus Species from Poultry Slaughterhouses in South Korea. Veterinary Sciences. 2024; 11(4):180. https://doi.org/10.3390/vetsci11040180
Chicago/Turabian StyleSon, Yongwoo, Yeung Bae Jin, Eun-Jeong Cho, Ae Ra Park, Rochelle A. Flores, Binh T. Nguyen, Seung Yun Lee, Bujinlkham Altanzul, Kwang Il Park, Wongi Min, and et al. 2024. "Comparative Analysis of Antibiotic Resistance and Biofilm Characteristics of Two Major Enterococcus Species from Poultry Slaughterhouses in South Korea" Veterinary Sciences 11, no. 4: 180. https://doi.org/10.3390/vetsci11040180
APA StyleSon, Y., Jin, Y. B., Cho, E.-J., Park, A. R., Flores, R. A., Nguyen, B. T., Lee, S. Y., Altanzul, B., Park, K. I., Min, W., & Kim, W. H. (2024). Comparative Analysis of Antibiotic Resistance and Biofilm Characteristics of Two Major Enterococcus Species from Poultry Slaughterhouses in South Korea. Veterinary Sciences, 11(4), 180. https://doi.org/10.3390/vetsci11040180