Abundance Trends of Immature Stages of Ticks at Different Distances from Hiking Trails from a Natural Park in North-Western Italy


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
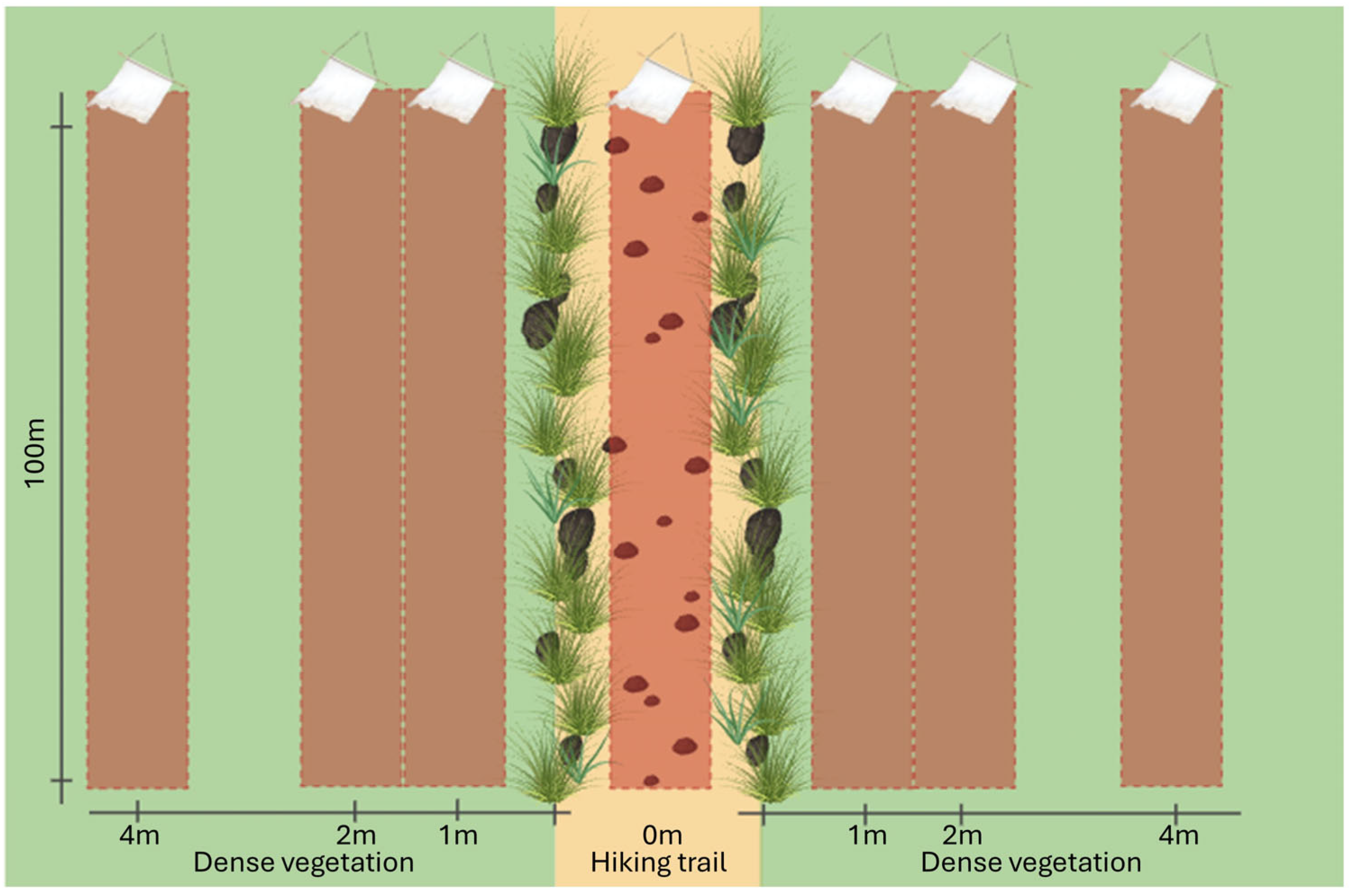
2.2. Sampling Design
2.3. Statistical Analysis
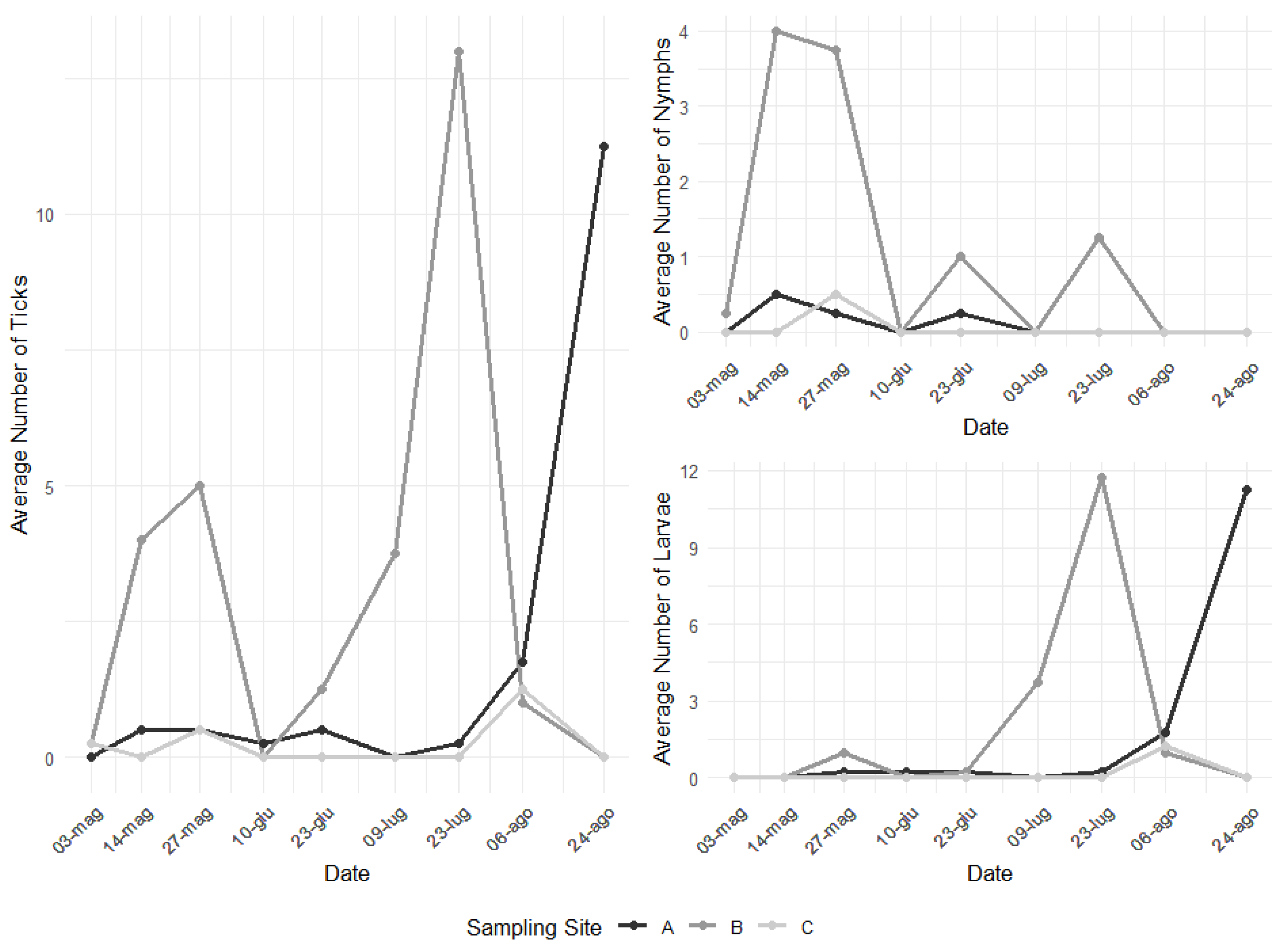
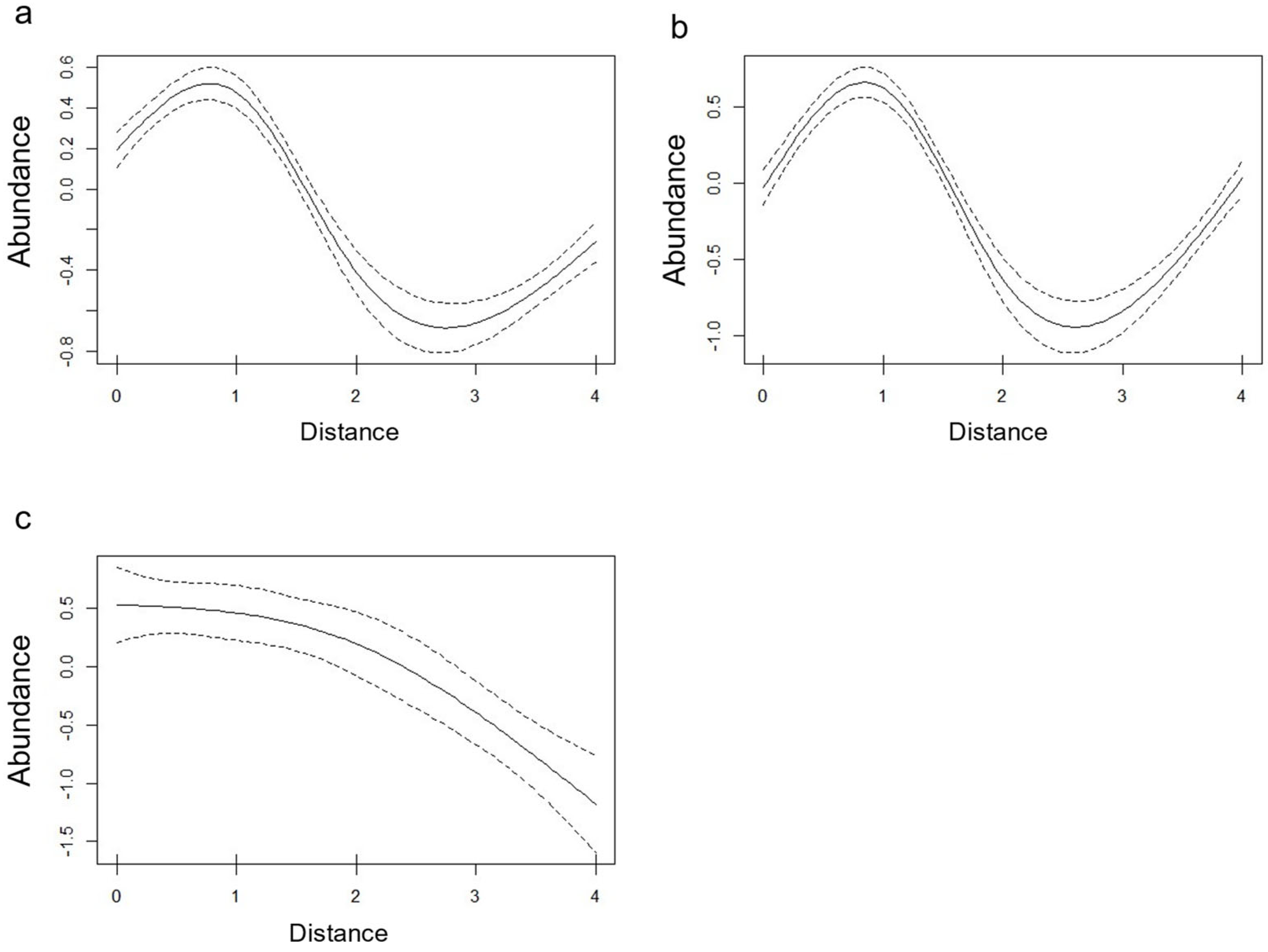
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mackenstedt, U.; Jenkins, D.; Romig, T. The role of wildlife in the transmission of parasitic zoonoses in peri-urban and urban areas. Int. J. Parasitol. Parasites Wildl. 2015, 4, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Kays, R.; Parsons, A.W.; Baker, M.C.; Kalies, E.L.; Forrester, T.; Costello, R.; Rota, C.T.; Millspaugh, J.J.; McShea, W.J. Does hunting or hiking affect wildlife communities in protected areas? J. Appl. Ecol. 2017, 54, 242–252. [Google Scholar] [CrossRef]
- Muhly, T.B.; Semeniuk, C.; Massolo, A.; Hickman, L.; Musiani, M. Human activity helps prey win the predator-prey space race. PLoS ONE 2011, 6, e17050. [Google Scholar] [CrossRef] [PubMed]
- Doherty, T.S.; Dickman, C.R.; Nimmo, D.G.; Ritchie, E.G. Multiple threats, or multiplying the threats? Interactions between invasive predators and other ecological disturbances. Biol. Conserv. 2015, 190, 60–68. [Google Scholar] [CrossRef]
- Erb, P.L.; McShea, W.J.; Guralnick, R.P. Anthropogenic influences on macro-level mammal occupancy in the appalachian trail corridor. PLoS ONE 2012, 7, e42574. [Google Scholar] [CrossRef]
- Mols, B.; Churchill, J.; Cromsigt, J.; Kuijper, D.; Smit, C. Recreation reduces tick density through fine-scale risk effects on deer space-use. Sci. Total. Environ. 2022, 839, 156222. [Google Scholar] [CrossRef]
- Shorb, C.M.; Freymiller, L.A.; Hernandez, D.L. Differential responses of prairie rodents to edge effects from recreational trails. Nat. Conserv. 2020, 41, 113–140. [Google Scholar] [CrossRef]
- Anderson, K.R.; Blekking, J.; Omodior, O. Tick trails: The role of online recreational trail reviews in identifying risk factors and behavioral recommendations associated with tick encounters in Indiana. BMC Public Health 2021, 21, 908. [Google Scholar] [CrossRef]
- Dobson, A.D.M.; Taylor, J.L.; Randolph, S.E. Tick (Ixodes ricinus) abundance and seasonality at recreational sites in the UK: Hazards in relation to fine-scale habitat types revealed by complementary sampling methods. Ticks Tick Borne Dis. 2011, 2, 67–74. [Google Scholar] [CrossRef]
- Hassett, E.; Diuk-Wasser, M.; Harrington, L.; Fernandez, P. Integrating tick density and park visitor behaviors to assess the risk of tick exposure in urban parks on Staten Island, New York. BMC Public Health 2022, 22, 1602. [Google Scholar] [CrossRef]
- Janzén, T.; Hammer, M.; Petersson, M.; Dinnétz, P. Factors responsible for Ixodes ricinus presence and abundance across a natu-ral-urban gradient. PLoS ONE 2023, 18, e0285841. [Google Scholar] [CrossRef] [PubMed]
- Iijima, H.; Watari, Y.; Furukawa, T.; Okabe, K. Importance of host abundance and microhabitat in tick abundance. J. Med. Èntomol. 2022, 59, 2110–2119. [Google Scholar] [CrossRef] [PubMed]
- Raši, T.; Majláth, I.; Bogdziewicz, M.; Dudek, K.; Majláthová, V.; Włodarek, J.; Almáši, M.; Vargová, B.; Tryjanowski, P. Tick distribution along animal tracks: Implication for preventative medicine. Ann. Agric. Environ. Med. 2018, 25, 360–363. [Google Scholar] [CrossRef]
- Van Gestel, M.; Verheyen, K.; Matthysen, E.; Heylen, D. Danger on the track? Tick densities near recreation infrastructures in forests. Urban For. Urban Green. 2021, 59, 126994. [Google Scholar] [CrossRef]
- Aureli, S.; Galuppi, R.; Ostanello, F.; Foley, J.E.; Bonoli, C.; Rejmanek, D.; Rocchi, G.; Orlandi, E.; Tampieri, M.P. Abundanceof questing ticks and molecular evidence for pathogens in ticks in three parks of Emilia-Romagnaregion of Northern Italy. Ann Agric. Environ. Med. 2015, 22, 459–466. [Google Scholar] [CrossRef]
- Corrain, R.; Drigo, M.; Fenati, M.; Menandro, M.L.; Mondin, A.; Pasotto, D.; Martini, M. Study on ticks and tick-borne zoonoses in public parks in Italy. Zoonoses Public Health 2012, 59, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Millet, I.; Ragionieri, M.; Tomassone, L.; Trentin, C.; Mannelli, A. Assessment of the exposure of people to questing ticks carrying agents of zoonoses in Aosta Valley, Italy. Veter. Sci. 2019, 6, 28. [Google Scholar] [CrossRef]
- Alekseev, A.N.; Jensen, P.M.; Dubinina, H.V.; Smirnova, L.A.; Makrouchina, N.A.; Zharkov, S.D. Peculiarities of behaviour of taiga (Ixodes persulcatus) and sheep (Ixodes ricinus) ticks (Acarina: Ixodidae) determined by different methods. Folia Parasitol. 2000, 47, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Vargová, B.; Majláth, I.; Kurimský, J.; Cimbala, R.; Zbojovský, J.; Tryjanowski, P.; Majláthová, V. Locomotor Activity of Ixodes ricinus Females in 900 MHz Electromagnetic Field. Life 2022, 12, 884. [Google Scholar] [CrossRef]
- Perret, J.-L.; Guerin, P.M.; Diehl, P.A.; Vlimant, M.; Gern, L. Darkness induces mobility, and saturation deficit limits questing duration, in the tick Ixodes ricinus. J. Exp. Biol. 2003, 206, 1809–1815. [Google Scholar] [CrossRef]
- Madhav, N.K.; Brownstein, J.S.; Tsao, J.I.; Fish, D. A Dispersal model for the range expansion of blacklegged tick (Acari: Ixodidae). J. Med. Èntomol. 2004, 41, 842–852. [Google Scholar] [CrossRef] [PubMed]
- Buczek, A.; Zając, Z.; Woźniak, A.; Kulina, D.; Bartosik, K. Locomotor activity of adult Dermacentor reticulatus ticks (Ixodida: Ix-odidae) in natural conditions. Ann. Agric. Environ. Med. 2017, 24, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.S. A carbon dioxide trap for prolonged sampling of Ixodes ricinus L. populations. Exp. Appl. Acarol. 1985, 1, 35–44. [Google Scholar] [CrossRef] [PubMed]
- ENETWILD-Consortium; Guerrasio, T.; Acevedo, P.P.; Apollonio, M.; Arnon, A.; Barroqueiro, C.; Belova, O.; Berdión, O.; Blanco-Aguiar, J.A.; Bijl, H.; et al. Wild ungulate density data generated by camera trapping in 37 European areas: First output of the European Observatory of Wildlife (EOW). EFSA Support. Publ. 2023, 20, 7892E. [Google Scholar] [CrossRef]
- Zanet, S.; Trisciuoglio, A.; Bottero, E.; de Mera, I.G.F.; Gortazar, C.; Carpignano, M.G.; Ferroglio, E. Piroplasmosis in wildlife: Babesia and Theileria affecting free-ranging ungulates and carnivores in the Italian Alps. Parasites Vectors 2014, 7, 70. [Google Scholar] [CrossRef]
- ENETWILD-Consortium; Guerrasio, T.; Apollonio, M.; Blanco, J.; Scandura, M.; Keuling, O.; Podgorski, T.; Plis, K.; Smith, G.; Ferroglio, E.; et al. Data generated by camera trapping in 40 areas in Europe including East and South Europe: Report of the field activities (May 2022). EFSA Support. Publ. 2022, 19, 7456E. [Google Scholar] [CrossRef]
- Keeping, D.; Pelletier, R. Animal density and track counts: Understanding the nature of observations based on animal movements. PLoS ONE 2014, 9, e96598. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; Mihalca, A.D.; Petney, T.N. Ticks of Europe and North Africa: A Guide to Species Identification; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Manilla, G.; Iori, A. Chiave illustrata delle zecche d’Italia. II: Stadi ninfali delle specie della sottofamiglia Ixodinate (Acari, Ix-odoidea, Ixodidae). Parassitologia 1993, 35, 37. [Google Scholar]
- Manilla, G.; Iori, A. Chiave illustrata delle zecche d’Italia. I: Stadi larvali delle specie della sottofamiglia Ixodinae (Acari, Ixodoidea, Ixodidae). Parassitologia 1992, 34, 83. [Google Scholar]
- Zając, Z.; Kulisz, J.; Bartosik, K.; Woźniak, A.; Dzierżak, M.; Khan, A. Environmental determinants of the occurrence and activity of Ixodes ricinus ticks and the prevalence of tick-borne diseases in eastern Poland. Sci. Rep. 2021, 11, 15472. [Google Scholar] [CrossRef]
- Tagliapietra, V.; Rosà, R.; Arnoldi, D.; Cagnacci, F.; Capelli, G.; Montarsi, F.; Hauffe, H.C.; Rizzoli, A. Saturation deficit and deer density affect questing activity and local abundance of Ixodes ricinus (Acari, Ixodidae) in Italy. Vet. Parasitol. 2011, 183, 114–124. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org/ (accessed on 15 May 2023).
- Salman, M.D. Ticks and Tick-Borne Diseases: Geographical Distribution and Control Strategies in the Euro-Asia Region; CABI: Wallingford, UK, 2012. [Google Scholar]
- Wood, S.N. Generalized Additive Models: An Introduction with R; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Ferroglio, E.; Vada, R.; Occhibove, F.; Fracchia, M.; Cicco, F.D.; Palencia, P.; Varzandi, A.R.; Zanet, S. An integrated approach to an emerging problem: Implementing a whole year of camera trap survey in evaluating the impact of wildlife on tick abundance. Transbound. Emerg. Dis. 2024, 2024, 4064855. [Google Scholar] [CrossRef]
- Daniels, T.J.; Fish, D. Spatial distribution and dispersal of unfed larval ixodes dammini (Acari: Ixodidae) in southern New York. Environ. Èntomol. 1990, 19, 1029–1033. [Google Scholar] [CrossRef]
- Gray, J.S. Ixodes ricinus seasonal activity: Implications of global warming indicated by revisiting tick and weather data. Int. J. Med. Microbiol. 2008, 298 (Suppl. S1), 19–24. [Google Scholar] [CrossRef]
- Kahl, O.; Gray, J.S. The biology of Ixodes ricinus with emphasis on its ecology. Ticks Tick-Borne Dis. 2023, 14, 102114. [Google Scholar] [CrossRef]
- Mejlon, H.; Jaenson, T. Questing behaviour of Ixodes ricinus ticks (Acari: Ixodidae). Exp. Appl. Acarol. 1997, 21, 747–754. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Adults | Nymphs | Larvae | Total | |
|---|---|---|---|---|
| Dermacentor sp. | 1 | 1 | ||
| Haemaphysalis concinna | 3 | 3 | ||
| Haemaphysalis punctata | 1 | 2 | 60 | 63 |
| Ixodes ricinus | 2 | 41 | 66 | 109 |
| Rhipicephalus sanguineus complex | 5 | 5 | ||
| Total | 3 | 46 | 132 | 181 |
| I. ricinus | H. punctata | H. concinna | Dermacentor sp. | Rhipicephalus sanguineus Complex | Total | |
|---|---|---|---|---|---|---|
| May | 1.06 | 0.11 | 0.06 | 0.00 | 0.00 | 1.23 |
| June | 0.21 | 0.08 | 0.04 | 0.00 | 0.00 | 0.33 |
| July | 1.67 | 1.04 | 0.00 | 0.00 | 0.13 | 2.84 |
| August | 1.08 | 1.33 | 0.00 | 0.04 | 0.08 | 2.53 |
| Sampling Site A | Sampling Site B | Sampling Site C | |
|---|---|---|---|
| 0 m | 2.89 | 3.11 | 0.22 |
| 1 m | 2.89 | 4.22 | 0.67 |
| 2 m | 0.56 | 2.33 | 0.00 |
| 4 m | 0.33 | 2.89 | 0.00 |
| Total | 1.67 | 3.14 | 0.22 |
| General | |||
|---|---|---|---|
| Parametric Coefficients | Estimate | Std. Error | Pr (>|z|) |
| (Intercept) | −2.50 | 0.12 | <2 × 10−16 |
| Date | 0.07 | 0.01 | 1.43 × 10−9 |
| Saturation deficit | 0.7 | 0.03 | <2 × 10−16 |
| Smooth terms | edf | Ref.df | p-value |
| s(distance) | 3 | 3 | <2 × 10−16 |
| Larvae | |||
| Parametric Coefficients | Estimate | Std. Error | Pr (>|z|) |
| (Intercept) | −4.42 | 0.15 | <2 × 10−16 |
| Date | 0.34 | 0.02 | <2 × 10−16 |
| Saturation deficit | 0.65 | 0.03 | <2 × 10−16 |
| Smooth terms | edf | Ref.df | p-value |
| s(distance) | 3 | 3 | <2 × 10−16 |
| Nymphs | |||
| Parametric Coefficients | Estimate | Std. Error | Pr (>|z|) |
| (Intercept) | −3.45 | 0.51 | 1.95 × 10−11 |
| Date | −0.62 | 0.07 | <2 × 10−16 |
| Saturation deficit | 1.41 | 0.19 | 1.34 × 10−13 |
| Smooth terms | edf | Ref.df | p-value |
| s(distance) | 2.06 | 2.4 | 2.25 × 10−7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vada, R.; Zanet, S.; Battisti, E.; Ferroglio, E. Abundance Trends of Immature Stages of Ticks at Different Distances from Hiking Trails from a Natural Park in North-Western Italy. Vet. Sci. 2024, 11, 508. https://doi.org/10.3390/vetsci11100508
Vada R, Zanet S, Battisti E, Ferroglio E. Abundance Trends of Immature Stages of Ticks at Different Distances from Hiking Trails from a Natural Park in North-Western Italy. Veterinary Sciences. 2024; 11(10):508. https://doi.org/10.3390/vetsci11100508
Chicago/Turabian StyleVada, Rachele, Stefania Zanet, Elena Battisti, and Ezio Ferroglio. 2024. "Abundance Trends of Immature Stages of Ticks at Different Distances from Hiking Trails from a Natural Park in North-Western Italy" Veterinary Sciences 11, no. 10: 508. https://doi.org/10.3390/vetsci11100508
APA StyleVada, R., Zanet, S., Battisti, E., & Ferroglio, E. (2024). Abundance Trends of Immature Stages of Ticks at Different Distances from Hiking Trails from a Natural Park in North-Western Italy. Veterinary Sciences, 11(10), 508. https://doi.org/10.3390/vetsci11100508