

Stability of Fly Maggot Peptides and Its Alleviating Effect on Lipopolysaccharide Combined with Hemocoagulase Oxidative Stress in Arbor Acres Chicks

 , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Bioactive Peptides
2.2.1. Source
2.2.2. Determination of Antioxidant Activities of FMP
2.2.3. Stability of Hydrolysate of FMP
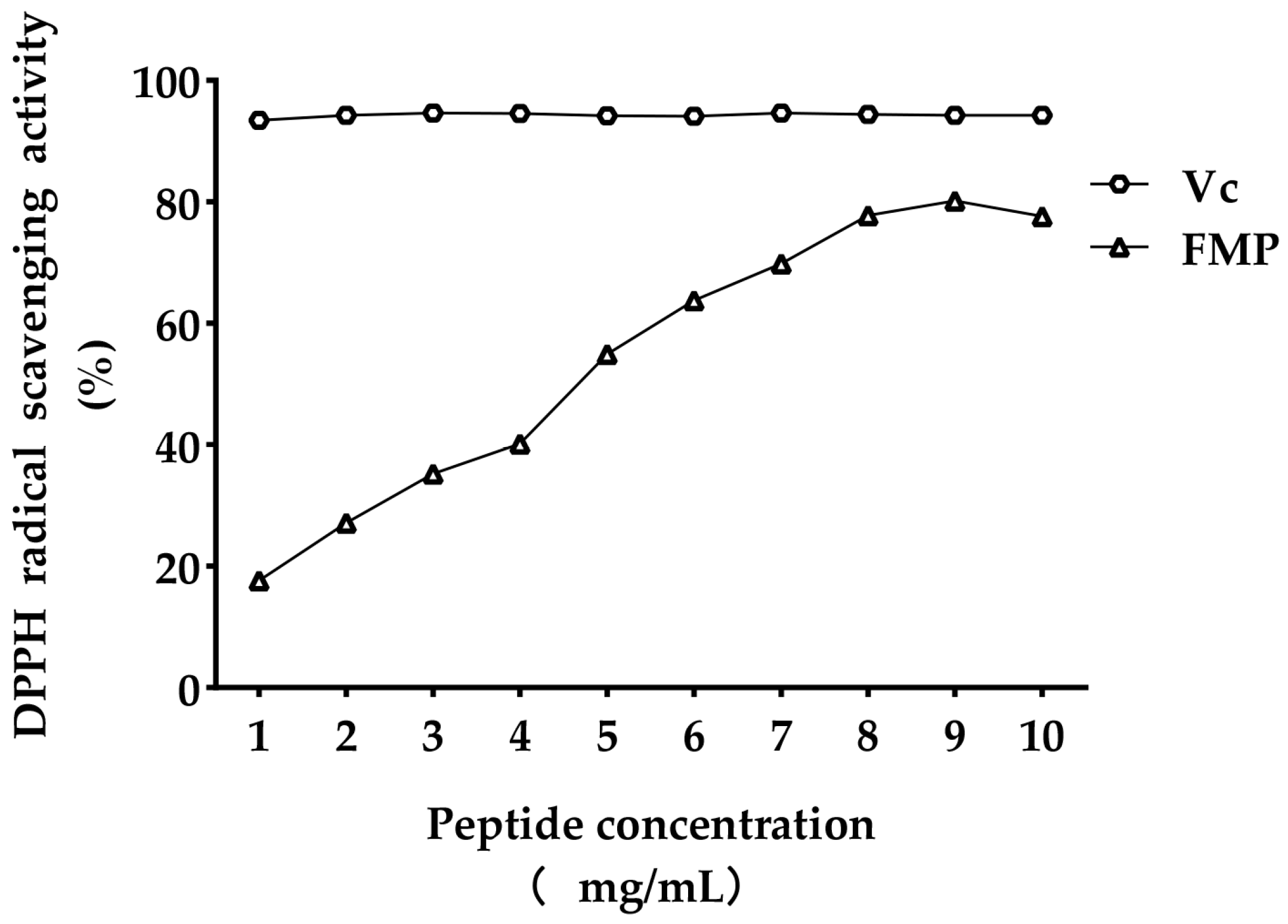
- : The antioxidant activity of FMP.
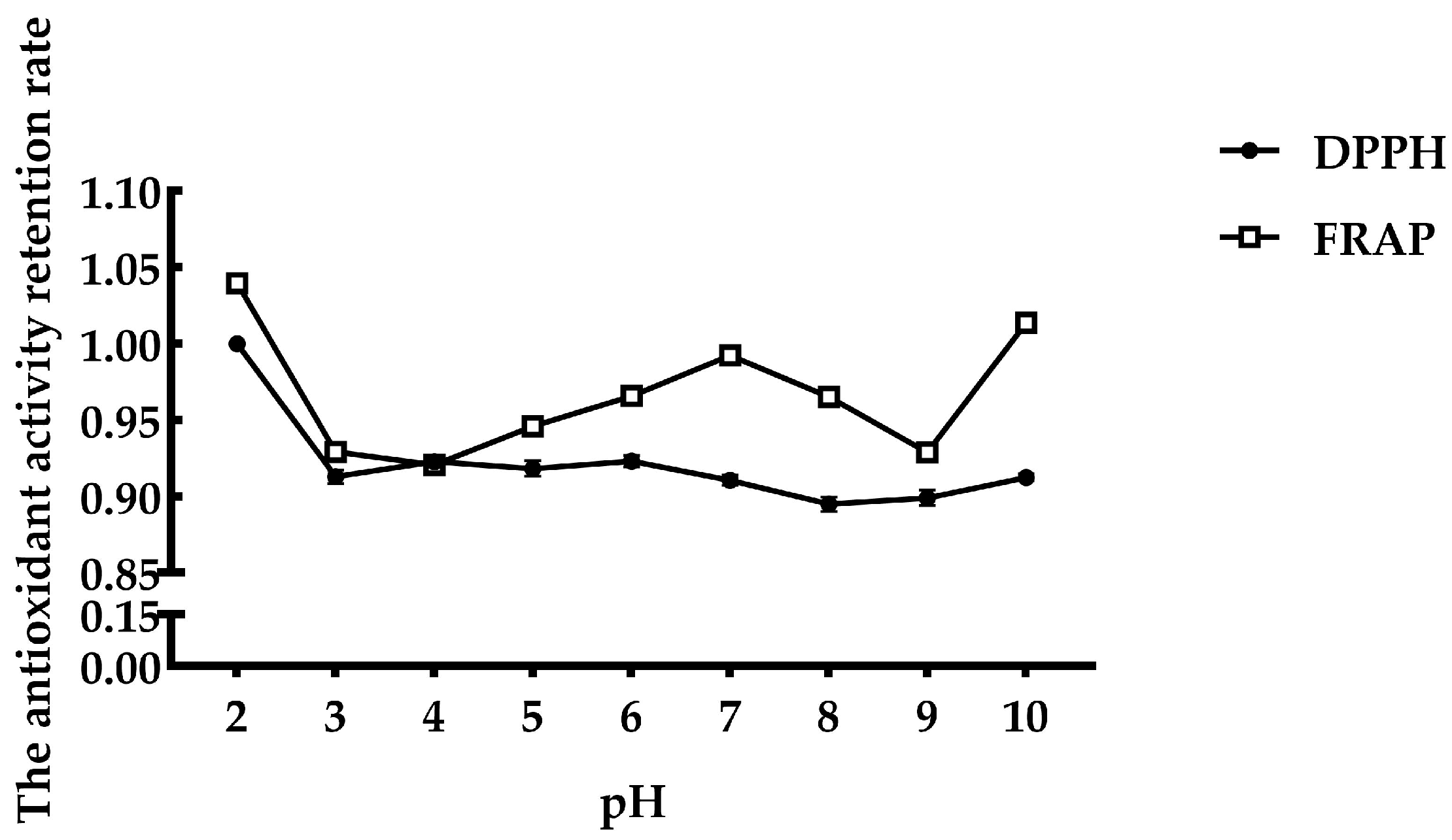
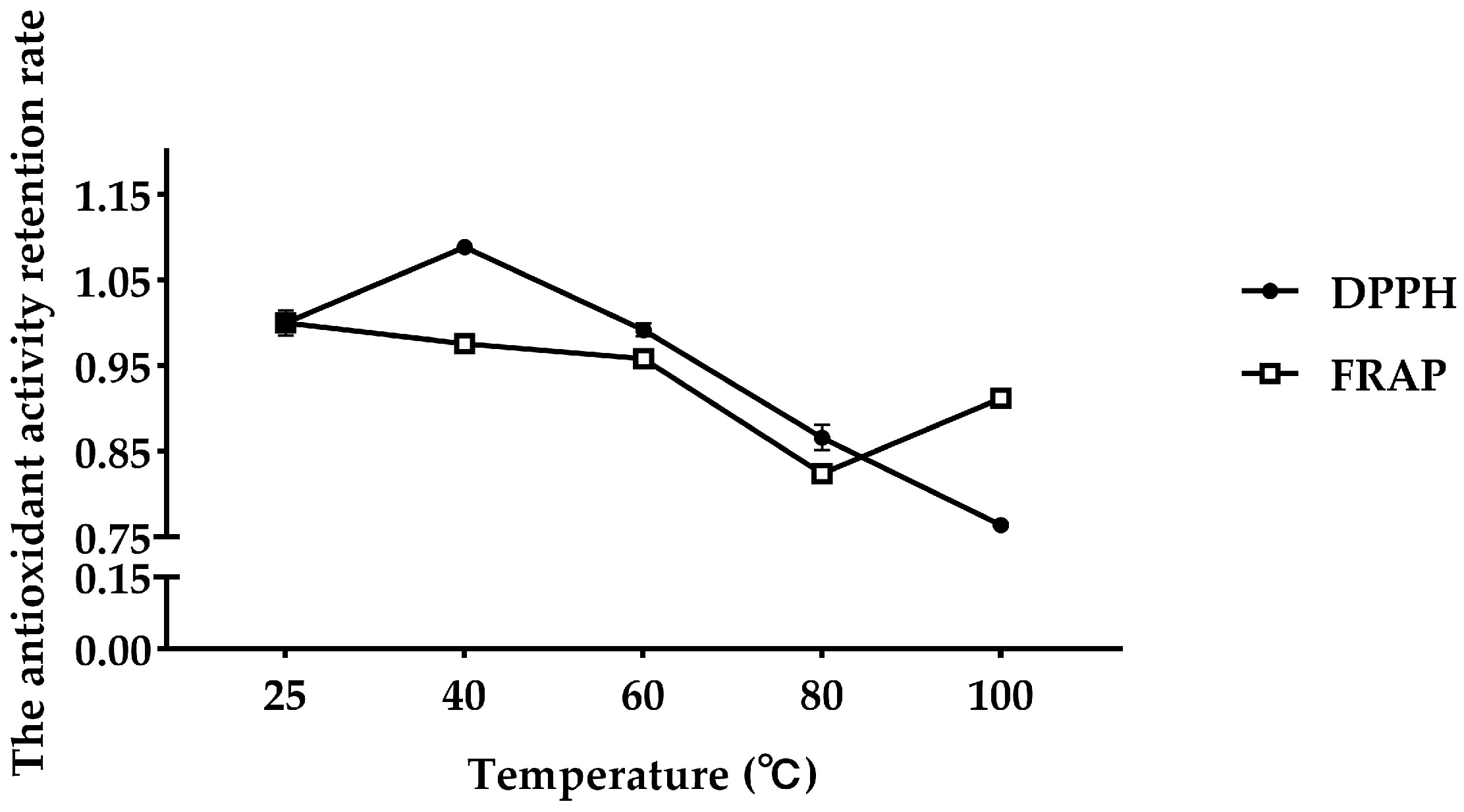
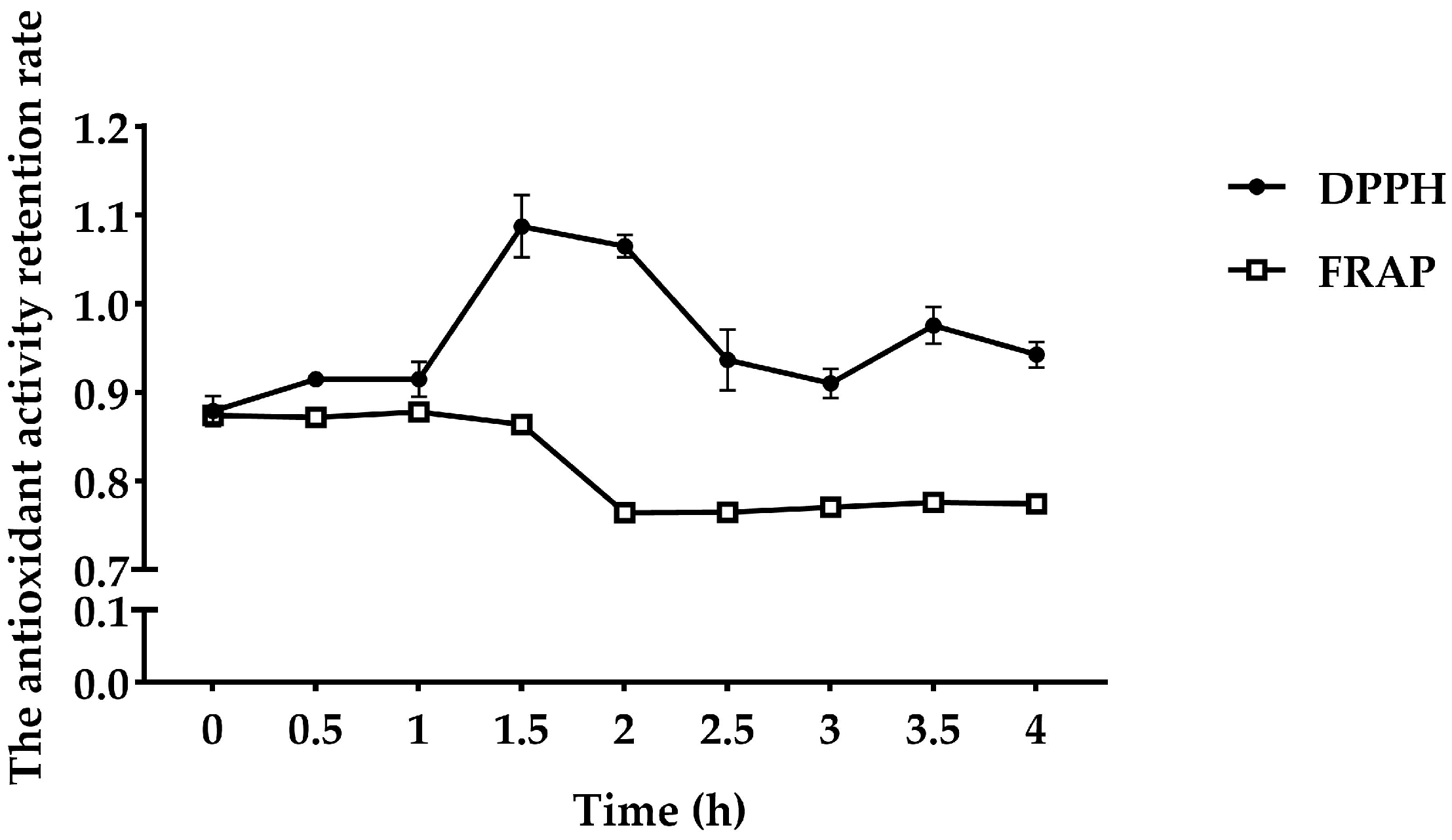
- : Antioxidant activity of FMP after temperature or acid–base conditions or simulated GI digestion.
2.3. Animal Experiment
2.3.1. Treatments and Sample Collection
2.3.2. Growth Performance, the Relative Weight of the Immune Organ, and Rectal Temperature Detection
2.3.3. Antioxidant and Inflammation Parameters
2.3.4. Histological Analysis
2.4. Statistical Analysis
3. Results
3.1. Antioxidant Activities of FMP
3.2. Stability of Antioxidant Peptides from FMP
3.3. Effect of Fly Maggot Polypeptide on Rectal Temperature, Growth Performance, and Relative Weight of Immune Organ
3.4. Effect of FMP on Serum Indexes at Different Time Periods
3.5. Effect of FMP on Inflammatory Cytokines of the Liver
3.6. Effect of FMP on Villus Height, Crypt Depth, and VCR of Small Intestine
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Surai, P.F. Antioxidant systems in poultry biology: Superoxide dismutase. J. Anim. Res. Nutr. 2016, 1, 8. [Google Scholar] [CrossRef]
- van den Bogaard, A.E.; Willems, R.; London, N.; Top, J.; Stobberingh, E.E. Antibiotic resistance of faecal enterococci in poultry, poultry farmers and poultry slaughterers. J. Antimicrob. Chemother. 2002, 49, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Singer, R.S.; Hofacre, C.L. Potential impacts of antibiotic use in poultry production. Avian Dis. 2006, 50, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Landy, N.; Kavyani, A. Effects of using a multi-strain probiotic on performance, immune responses and cecal microflora composition in broiler chickens reared under cyclic heat stress condition. Iran. J. Appl. Anim. Sci. 2013, 3, 703–798. [Google Scholar]
- Toghyani, M.; kazem Mosavi, S.; Modaresi, M.; Landy, N. Evaluation of kefir as a potential probiotic on growth performance, serum biochemistry and immune responses in broiler chicks. Anim. Nutr. 2015, 1, 305–309. [Google Scholar] [CrossRef]
- Ceylan, N.; Çiftçi, İ.; Ilhan, Z. The effects of some alternative feed additives for antibiotic growth promoters on the performance and gut microflora of broiler chicks. Turk. J. Vet. Anim. Sci. 2003, 27, 727–733. [Google Scholar]
- Upadhayay, U.; Vishwa, P.C.V. Growth promoters and novel feed additives improving poultry production and health, bioactive principles and beneficial applications: The trends and advances-a review. Int. J. Pharmacol. 2014, 10, 129–159. [Google Scholar]
- Hou, Y.; Wu, Z.; Dai, Z.; Wang, G.; Wu, G. Protein hydrolysates in animal nutrition: Industrial production, bioactive peptides, and functional significance. J. Anim. Sci. Biotechnol. 2017, 8, 24. [Google Scholar] [CrossRef]
- Moller, N.P.; Scholz-Ahrens, K.E.; Roos, N.; Schrezenmeir, J. Bioactive peptides and proteins from foods: Indication for health effects. Eur. J. Nutr. 2008, 47, 171–182. [Google Scholar] [CrossRef]
- Sayed, Y.; Hassan, M.; Salem, H.M.; Al-Amry, K.; Eid, G.E. Prophylactic influences of prebiotics on gut microbiome and immune response of heat-stressed broiler chickens. Sci. Rep. 2023, 13, 13991. [Google Scholar] [CrossRef]
- Meng, W.S.; Zou, Q.; Xiao, Y.; Ma, W.; Zhang, J.; Wang, T.; Li, D. Growth performance and cecal microbiota of broiler chicks as affected by drinking water disinfection and/or herbal extract blend supplementation. Poult. Sci. 2023, 102, 102707. [Google Scholar] [CrossRef] [PubMed]
- Mouithys-Mickalad, A.; Schmitt, E.; Dalim, M.; Franck, T.; Tome, N.M.; van Spankeren, M.; Serteyn, D.; Paul, A. Black Soldier Fly (Hermetia illucens) Larvae Protein Derivatives: Potential to Promote Animal Health. Animals 2020, 10, 941. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, H.R.; Isono, H.; Miyata, T. Potential antioxidant bioactive peptides from camel milk proteins. Anim. Nutr. 2018, 4, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Hall, F.; Reddivari, L.; Liceaga, A.M. Identification and characterization of edible cricket peptides on hypertensive and glycemic in vitro inhibition and their anti-inflammatory activity on RAW 264.7 macrophage cells. Nutrients 2020, 12, 3588. [Google Scholar] [CrossRef] [PubMed]
- Kotzamanis, Y.P.; Gisbert, E.; Gatesoupe, F.J.; Zambonino Infante, J.; Cahu, C. Effects of different dietary levels of fish protein hydrolysates on growth, digestive enzymes, gut microbiota, and resistance to Vibrio anguillarum in European sea bass (Dicentrarchus labrax) larvae. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 147, 205–214. [Google Scholar] [CrossRef]
- Mohd Zubir, M.Z.; Holloway, S.; Mohd Noor, N. Maggot Therapy in Wound Healing: A Systematic Review. Int. J. Environ. Res. Public Health 2020, 17, 6103. [Google Scholar] [CrossRef]
- Chu, F.; Jin, X.; Ma, H.; Lu, X.; Zhu, J. Effect of Musca domestic maggot polypeptide extract on HUVEC dysfunction induced by early-activated macrophages. Pharm. Biol. 2016, 54, 572–575. [Google Scholar]
- Li, Z.; Zhao, S.; Xin, X.; Zhang, B.; Thomas, A.; Charles, A.; Lee, K.S.; Jin, B.R.; Gui, Z. Purification and characterization of a novel immunomodulatory hexapeptide from alcalase hydrolysate of ultramicro-pretreated silkworm (Bombyx mori) pupa protein. J. Asia-Pac. Entomol. 2019, 22, 633–637. [Google Scholar] [CrossRef]
- Powell, M.F. Peptide stability in aqueous parenteral formulations: Prediction of chemical stability based on primary sequence. In Formulation and Delivery of Proteins and Peptides; ACS Publications: Washington, DC, USA, 1994. [Google Scholar]
- Gao, Y.; Fang, H.; Fang, L.; Liu, D.; Liu, J.; Su, M.; Fang, Z.; Ren, W.; Jiao, H. The Modification and Design of Antimicrobial Peptide. Curr. Pharm. Des. 2018, 24, 904–910. [Google Scholar] [CrossRef]
- Lee, S.M.; Son, K.N.; Shah, D.; Ali, M.; Balasubramaniam, A.; Shukla, D.; Aakalu, V.K. Histatin-1 Attenuates LPS-Induced Inflammatory Signaling in RAW264.7 Macrophages. Int. J. Mol. Sci. 2021, 22, 7856. [Google Scholar] [CrossRef]
- Niki, E. Oxidative stress and antioxidants: Distress or eustress? Arch. Biochem. Biophys. 2016, 595, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Kattoor, A.J.; Pothineni, N.V.K.; Palagiri, D.; Mehta, J.L. Oxidative Stress in Atherosclerosis. Curr. Atheroscler. Rep. 2017, 19, 42. [Google Scholar] [CrossRef] [PubMed]
- Teleanu, D.M.; Niculescu, A.G.; Lungu, I.I.; Radu, C.I.; Vladacenco, O.; Roza, E.; Costachescu, B.; Grumezescu, A.M.; Teleanu, R.I. An Overview of Oxidative Stress, Neuroinflammation, and Neurodegenerative Diseases. Int. J. Mol. Sci. 2022, 23, 5938. [Google Scholar] [CrossRef] [PubMed]
- Lukasiewicz, J.; Lugowski, C. Biologic activity of lipopolysaccharides. Postepy Hig. Med. Dosw 2003, 57, 33–53. [Google Scholar] [PubMed]
- Wang, X.; Quinn, P.J. Lipopolysaccharide: Biosynthetic pathway and structure modification. Prog. Lipid Res. 2010, 49, 97–107. [Google Scholar] [CrossRef]
- Ye, C.; Zhang, N.; Zhao, Q.; Xie, X.; Li, X.; Zhu, H.P.; Peng, C.; Huang, W.; Han, B. Evodiamine alleviates lipopolysaccharide-induced pulmonary inflammation and fibrosis by activating apelin pathway. Phytother. Res. 2021, 35, 3406–3417. [Google Scholar] [CrossRef]
- Zhang, Q.; Luo, P.; Xia, F.; Tang, H.; Chen, J.; Zhang, J.; Liu, D.; Zhu, Y.; Liu, Y.; Gu, L.; et al. Capsaicin ameliorates inflammation in a TRPV1-independent mechanism by inhibiting PKM2-LDHA-mediated Warburg effect in sepsis. Cell. Chem. Biol. 2022, 29, 1248–1259 e1246. [Google Scholar] [CrossRef]
- Xiao, K.; Zou, W.H.; Yang, Z.; Rehman, Z.U.; Ansari, A.R.; Yuan, H.R.; Zhou, Y.; Cui, L.; Peng, K.M.; Song, H. The role of visfatin on the regulation of inflammation and apoptosis in the spleen of LPS-treated rats. Cell Tissue Res. 2015, 359, 605–618. [Google Scholar] [CrossRef]
- Zhou, R.-J.; Ye, H.; Wang, F.; Wang, J.-L.; Xie, M.-L. Apigenin inhibits d-galactosamine/LPS-induced liver injury through upregulation of hepatic Nrf-2 and PPARγ expressions in mice. Biochem. Biophys. Res. Commun. 2017, 493, 625–630. [Google Scholar] [CrossRef]
- Katiyar, A.K.; Vegad, J.L.; Awadhiya, R.P. Attempted induction of local Shwartzman reaction in the chicken. Res. Vet. Sci. 1987, 43, 405–406. [Google Scholar] [CrossRef]
- Gao, X.; Xiao, Z.H.; Liu, M.; Zhang, N.Y.; Khalil, M.M.; Gu, C.Q.; Qi, D.S.; Sun, L.H. Dietary Silymarin Supplementation Alleviates Zearalenone-Induced Hepatotoxicity and Reproductive Toxicity in Rats. J. Nutr. 2018, 148, 1209–1216. [Google Scholar] [CrossRef] [PubMed]
- Marcuse, R. Antioxidative effect of amino-acids. Nature 1960, 186, 886–887. [Google Scholar] [CrossRef] [PubMed]
- Marcuse, R. The effect of some amino acids on the oxidation of linoleic acid and its methyl ester. J. Am. Oil Chem. Soc. 1962, 39, 97–103. [Google Scholar] [CrossRef]
- Karel, M.; Tannenbaum, S.R.; Wallace, D.H.; Maloney, H. Autoxidation of methyl linoleate in freeze-dried model systems. III. Effects of added amino acids. J. Food Sci. 1966, 31, 892–896. [Google Scholar] [CrossRef]
- Yamashoji, S.; Yoshida, H.; Kajimoto, G. Effects of amino acids on the photosensitized oxidation of linoleic acid in an aqueous ethanol solution. Agric. Biol. Chem. 1979, 43, 1505–1511. [Google Scholar]
- Saiga, A.; Tanabe, S.; Nishimura, T. Antioxidant activity of peptides obtained from porcine myofibrillar proteins by protease treatment. J. Agric. Food Chem. 2003, 51, 3661–3667. [Google Scholar] [CrossRef]
- Li, Y.Q.; Bao, R.Y.; Wang, Q.; Qiu, P.F.; Li, X.C.; Shi, H.Y.; Zhang, H.W.; Wang, X.M. Protective Effects of Antioxidant Peptides from Maggot Oxidative Stress Injury of IPEC-J2C Cells. China Anim. Husb. Vet. Med. 2023, 50, 1836–1844. [Google Scholar]
- Samaranayaka, A.G.P.; Li-Chan, E.C.Y. Food-derived peptidic antioxidants: A review of their production, assessment, and potential applications. J. Funct. Foods 2011, 3, 229–254. [Google Scholar] [CrossRef]
- Zhu, C.Z.; Zhang, W.G.; Kang, Z.L.; Zhou, G.H.; Xu, X.L. Stability of an antioxidant peptide extracted from Jinhua ham. Meat Sci. 2014, 96, 783–789. [Google Scholar] [CrossRef]
- Grutter, M.G.; Hawkes, R.B.; Matthews, B.W. Molecular basis of thermostability in the lysozyme from bacteriophage T4. Nature 1979, 277, 667–669. [Google Scholar] [CrossRef]
- Cianciosi, D.; Forbes-Hernández, T.Y.; Giampieri, F.; Zhang, J.; Ansary, J.; Pacetti, M.; Quiles, J.L.; Simal-Gandara, J.; Battino, M. Effect of in vitro gastrointestinal digestion on the bioaccessibility of phenolic compounds and antioxidant activity of manuka honey. eFood 2020, 1, 85–93. [Google Scholar] [CrossRef]
- Ni, X.; Zhai, Z.; Xin, Z.; Tan, C.; Chen, Y.; Deng, J.; Liao, P.; Zhang, L.; Xiao, Z.; Deng, B. Antioxidant properties and transepithelial transportation of di-/tripeptides derived from simulated gastrointestinal digestion of pig blood cells hydrolysates. eFood 2020, 1, 254–269. [Google Scholar] [CrossRef]
- Zhu, L.; Chen, J.; Tang, X.; Xiong, Y.L. Reducing, radical scavenging, and chelation properties of in vitro digests of alcalase-treated zein hydrolysate. J. Agric. Food Chem. 2008, 56, 2714–2721. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.M.; Chen, X.; Huang, J.C.; Zhou, X.H.; Huang, M.; Zhou, G.H. Stability of antioxidant peptides from duck meat after post-mortem ageing. Int. J. Food Sci. Technol. 2017, 52, 2513–2521. [Google Scholar] [CrossRef]
- Ao, J.; Li, B. Stability and antioxidative activities of casein peptide fractions during simulated gastrointestinal digestion in vitro: Charge properties of peptides affect digestive stability. Food Res. Int. 2013, 52, 334–341. [Google Scholar] [CrossRef]
- Rahal, A.; Kumar, A.; Singh, V.; Yadav, B.; Tiwari, R.; Chakraborty, S.; Dhama, K. Oxidative stress, prooxidants, and antioxidants: The interplay. Biomed. Res. Int. 2014, 2014, 761264. [Google Scholar] [CrossRef]
- van Leeuwen, P.; Mouwen, J.M.; van der Klis, J.D.; Verstegen, M.W. Morphology of the small intestinal mucosal surface of broilers in relation to age, diet formulation, small intestinal microflora and performance. Br. Poult. Sci. 2004, 45, 41–48. [Google Scholar] [CrossRef]
- Ghazi Harsini, S.; Habibiyan, M.; Moeini, M.M.; Abdolmohammadi, A.R. Effects of Dietary Selenium, Vitamin E, and Their Combination on Growth, Serum Metabolites, and Antioxidant Defense System in Skeletal Muscle of Broilers under Heat Stress. Biol. Trace Elem. Res. 2012, 148, 322–330. [Google Scholar] [CrossRef]
- Karimzadeh, S.; Teimouri Yansari, A. Effects of canola bioactive peptides on performance, digestive enzyme activities, nutrient digestibility, intestinal morphology and gut microflora in broiler chickens. Poult. Sci. J. 2016, 4, 27–36. [Google Scholar]
- Abdollahi, M.; Zaefarian, F.; Gu, Y.; Xiao, W.; Jia, J.; Ravindran, V. Influence of soybean bioactive peptides on growth performance, nutrient utilisation, digestive tract development and intestinal histology in broilers. J. Appl. Anim. Nutr. 2017, 5, e7. [Google Scholar] [CrossRef]
- Landy, N.; Kheiri, F.; Faghani, M. Effects of periodical application of bioactive peptides derived from cottonseed on performance, immunity, total antioxidant activity of serum and intestinal development of broilers. Anim. Nutr. 2021, 7, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Liu, S.; Hou, L.; Li, K.; Wang, L.; Gao, K.; Yang, X.; Jiang, Z. Supplemental choline modulates growth performance and gut inflammation by altering the gut microbiota and lipid metabolism in weaned piglets. J. Nutr. 2021, 151, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yan, A.; Liu, X.; Ma, Y.; Zhao, F.; Wang, M.; Loor, J.J.; Wang, H. Melatonin ameliorates ochratoxin A induced liver inflammation, oxidative stress and mitophagy in mice involving in intestinal microbiota and restoring the intestinal barrier function. J. Hazard. Mater. 2021, 407, 124489. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.-Q.; Sa, R.-N.; Tong, J.-M. Effect of Apramycin on intestinal flora and intestinal morphology of Piglets. Acta Vet. Zootech. Sin. 2003, 34, 250–257. [Google Scholar]
- Feng, J.; Liu, X.; Xu, Z.R.; Wang, Y.Z.; Liu, J.X. Effects of fermented soybean meal on digestive enzyme activities and intestinal morphology in broilers. Poult. Sci. 2007, 86, 1149–1154. [Google Scholar] [CrossRef]
- Sakurai, H.; Suzuki, S.; Kawasaki, N.; Nakano, H.; Okazaki, T.; Chino, A.; Doi, T.; Saiki, I. Tumor necrosis factor-α-induced IKK phosphorylation of NF-κB p65 on serine 536 is mediated through the TRAF2, TRAF5, and TAK1 signaling pathway. J. Biol. Chem. 2003, 278, 36916–36923. [Google Scholar] [CrossRef]
- Chen, Q.; Liang, Z.; Yue, Q.; Wang, X.; Siu, S.W.I.; Pui-Man Hoi, M.; Lee, S.M. A Neuropeptide Y/F-like Polypeptide Derived from the Transcriptome of Turbinaria peltata Suppresses LPS-Induced Astrocytic Inflammation. J. Nat. Prod. 2022, 85, 1569–1580. [Google Scholar] [CrossRef]
- Ben-Neriah, Y.; Karin, M. Inflammation meets cancer, with NF-κB as the matchmaker. Nat. Immunol. 2011, 12, 715–723. [Google Scholar] [CrossRef]
- Zhang, D.; Li, N.; Wang, Y.; Lu, W.; Zhang, Y.; Chen, Y.; Deng, X.; Yu, X. Methane ameliorates post-operative cognitive dysfunction by inhibiting microglia NF-kappaB/MAPKs pathway and promoting IL-10 expression in aged mice. Int. Immunopharmacol. 2019, 71, 52–60. [Google Scholar] [CrossRef]
- Montagne, L.; Pluske, J.; Hampson, D. A review of interactions between dietary fibre and the intestinal mucosa, and their consequences on digestive health in young non-ruminant animals. Anim. Feed. Sci. Technol. 2003, 108, 95–117. [Google Scholar] [CrossRef]
- Guo, Y.; Balasubramanian, B.; Zhao, Z.H.; Liu, W.C. Marine algal polysaccharides alleviate aflatoxin B1-induced bursa of Fabricius injury by regulating redox and apoptotic signaling pathway in broilers. Poult. Sci. 2021, 100, 844–857. [Google Scholar] [CrossRef] [PubMed]
- Rajput, S.A.; Zhang, C.; Feng, Y.; Wei, X.T.; Khalil, M.M.; Rajput, I.R.; Baloch, D.M.; Shaukat, A.; Rajput, N.; Qamar, H. Proanthocyanidins alleviates aflatoxinB1-induced oxidative stress and apoptosis through mitochondrial pathway in the bursa of fabricius of broilers. Toxins 2019, 11, 157. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative Stress: A Key Modulator in Neurodegenerative Diseases. Molecules 2019, 24, 1583. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | 3 h | 6 h | 12 h | 24 h |
|---|---|---|---|---|
| CG | 40.89 ± 0.10 a | 40.67 ± 0.09 a | 40.98 ± 0.18 a | 40.90 ± 0.12 b |
| DG | 41.17 ± 0.13 a | 41.22 ± 0.12 ab | 41.54 ± 0.12 a | 40.93 ± 0.09 ab |
| VG | 41.21 ± 0.06 a | 41.21 ± 0.09 b | 41.34 ± 0.09 a | 40.73 ± 0.11 ab |
| LPG | 41.19 ± 0.16 a | 41.22 ± 0.11 b | 41.54 ± 0.11 a | 40.91 ± 0.09 ab |
| MPG | 41.33 ± 0.09 a | 41.18 ± 0.11 b | 41.08 ± 0.11 a | 40.54 ± 0.11 a |
| HPG | 41.25 ± 0.14 a | 41.23 ± 0.06 b | 41.28 ± 0.06 a | 40.95 ± 0.09 ab |
| p-value | 0.156 | 0.003 | 0.087 | 0.003 |
| Groups | Initial Weight (g) | Final Weight (g) | Average Daily Gain (ADG, g/d) |
|---|---|---|---|
| CG | 59.68 ± 0.83 a | 149.36 ± 6.16 a | 11.21 ± 0.61 ab |
| DG | 59.38 ± 0.88 a | 127.77 ± 8.12 a | 8.23 ± 0.92 a |
| VG | 58.70 ± 0.52 a | 170.12 ± 5.32 a | 13.93 ± 0.71 b |
| LPG | 59.45 ± 0.65 a | 143.00 ± 10.50 a | 10.44 ± 1.89 ab |
| MPG | 57.44 ± 0.55 a | 146.02 ± 15.07 a | 10.87 ± 0.05 ab |
| HPG | 59.04 ± 0.93 a | 165.82 ± 19.11 a | 16.22 ± 3.52 b |
| p-value | 0.06 | 0.100 | 0.015 |
| Groups | Bursa of Fabricius | Spleen |
|---|---|---|
| CG | 4.29 ± 0.12 a | 1.22 ± 0.07 a |
| DG | 4.11 ± 0.16 a | 1.60 ± 0.06 b |
| VG | 4.33 ± 0.20 a | 1.17 ± 0.05 a |
| LPG | 3.72 ± 0.18 a | 1.54 ± 0.08 b |
| MPG | 3.72 ± 0.14 a | 1.35 ± 0.05 ab |
| HPG | 3.85 ± 0.15 a | 1.14 ± 0.04 a |
| p-value | 0.107 | <0.001 |
| (a) | ||||
|---|---|---|---|---|
| Groups | GSH-Px (μmol/L) | MDA (nmol/mL) | T-AOC (mM) | T-SOD (U/mL) |
| CG | 233.78 ± 1.34 a | 3.90 ± 0.32 ab | 1.57 ± 0.06 b | 104.85 ± 0.87 a |
| DG | 238.54 ± 1.62 ab | 5.06 ± 0.31 b | 0.93 ± 0.11 a | 105.77 ± 4.27 a |
| VG | 238.61 ± 1.59 ab | 3.82 ± 0.12 a | 1.54 ± 0.07 b | 108.51 ± 1.17 a |
| LPG | 236.31 ± 1.55 ab | 4.75 ± 0.27 ab | 1.28 ± 0.06 b | 102.60 ± 2.47 a |
| MPG | 241.68 ± 3.16 b | 3.76 ± 0.32 a | 1.42 ± 0.04 b | 104.12 ± 1.16 a |
| HPG | 238.84 ± 1.37 ab | 4.17 ± 0.30 ab | 1.36 ± 0.04 b | 103.46 ± 1.74 a |
| p-value | 0.101 | 0.009 | <0.001 | 0.462 |
| (b) | ||||
| CG | 302.49 ± 2.77 a | 4.00 ± 0.15 a | 0.77 ± 0.06 ab | 99.15 ± 0.72 a |
| DG | 315.53 ± 1.89 c | 6.72 ± 0.16 b | 0.74 ± 0.05 ab | 99.75 ± 1.61 a |
| VG | 308.31 ± 2.65 abc | 4.23 ± 0.15 a | 0.80 ± 0.03 ab | 102.21 ± 1.10 a |
| LPG | 314.17 ± 1.14 bc | 4.62 ± 0.38 a | 0.70 ± 0.17 a | 101.00 ± 0.87 a |
| MPG | 304.05 ± 4.36 ab | 4.03 ± 0.30 a | 0.83 ± 0.10 ab | 99.96 ± 1.69 a |
| HPG | 306.87 ± 2.61 abc | 4.38 ± 0.22 a | 0.93 ± 0.19 b | 98.14 ± 0.93 a |
| p-value | 0.003 | <0.001 | 0.051 | 0.257 |
| (c) | ||||
| CG | 252.93 ± 1.77 a | 4.07 ± 0.32 b | 1.00 ± 0.05 ab | 98.91 ± 1.41 a |
| DG | 249.87 ± 1.64 a | 4.16 ± 0.09 b | 1.06 ± 0.09 ab | 97.43 ± 0.79 a |
| VG | 250.20 ± 3.40 a | 3.05 ± 0.08 a | 1.32 ± 0.07 b | 104.21 ± 1.7 ab |
| LPG | 250.20 ± 3.40 a | 3.34 ± 0.32 ab | 1.00 ± 0.07 a | 99.71 ± 2.03 a |
| MPG | 256.91 ± 2.81 a | 3.04 ± 0.08 a | 1.00 ± 0.06 ab | 108.59 ± 2.29 b |
| HPG | 250.94 ± 3.03 a | 3.59 ± 0.11 ab | 1.03 ± 0.06 ab | 109.91 ± 1.66 b |
| p-value | 0.35 | <0.001 | 0.028 | <0.001 |
| Groups | IL-1β | IL-6 | IL-10 | TNF-α |
|---|---|---|---|---|
| CG | 22.19 ± 0.35 c | 0.44 ± 0.01 b | 24.04 ± 4.87 ab | 2.37 ± 0.56 b |
| DG | 24.33 ± 0.21 d | 0.78 ± 0.01 c | 25.39 ± 4.23 d | 3.13 ± 0.07 b |
| VG | 19.97 ± 0.14 ab | 0.48 ± 0.02 b | 22.13 ± 2.56 bc | 0.54 ± 0.11 a |
| LPG | 18.92 ± 0.33 a | 0.25 ± 0.01 a | 22.63 ± 4.43 bc | 0.74 ± 0.07 a |
| MPG | 21.73 ± 0.64 c | 0.27 ± 0.02 a | 18.54 ± 1.55 a | 0.93 ± 0.05 a |
| HPG | 21.44 ± 0.33 bc | 0.42 ± 0.02 b | 20.53 ± 3.31 abc | 0.56 ± 0.08 a |
| p-value | <0.001 | <0.001 | <0.001 | <0.001 |
| (a) | |||
|---|---|---|---|
| Groups | Villus Height (μM) | Crypt Depth (μM) | Villus Height to Crypt Depth Ratio (VCR) |
| CG | 1319.52 ± 42.89 b | 97.99 ± 4.67 b | 14.36 ± 1.24 a |
| DG | 932.31 ± 40.63 a | 81.31 ± 3.23 ab | 11.42 ± 0.34 a |
| VG | 1183.96 ± 54.68 ab | 74.58 ± 3.34 a | 15.66 ± 1.83 a |
| LPG | 1088.3 ± 35.73 ab | 76.4 ± 4.14 a | 14.57 ± 2.14 a |
| MPG | 1150.11 ± 30.8 ab | 75.13 ± 3.16 a | 15.47 ± 0.95 a |
| HPG | 1275.57 ± 31.29 ab | 88.33 ± 6.91 ab | 15.05 ± 1.17 a |
| p-value | <0.001 | <0.001 | 0.602 |
| (b) | |||
| CG | 909.38 ± 16.54 b | 77.20 ± 2.11 a | 11.04 ± 0.58 a |
| DG | 774.79 ± 27.75 b | 78.25 ± 2.21 a | 9.97 ± 0.83 a |
| VG | 779.70 ± 67.68 b | 65.78 ± 3.34 a | 9.88 ± 1.22 a |
| LPG | 850.55 ± 56.11 a | 71.04 ± 5.89 a | 11.47 ± 1.96 a |
| MPG | 802.83 ± 60.56 a | 78.12 ± 2.82 a | 11.71 ± 0.97 a |
| HPG | 952.38 ± 53.00 a | 72.91 ± 1.88 a | 13.1 ± 0.97 a |
| p-value | <0.001 | 0.107 | 0.31 |
| (c) | |||
| CG | 686.04 ± 18.40 ab | 70.20 ± 1.67 b | 10.47 ± 0.97 ab |
| DG | 622.69 ± 15.46 bc | 68.05 ± 2.30 ab | 9.00 ± 0.39 ab |
| VG | 530.51 ± 28.12 d | 68.12 ± 1.56 ab | 8.32 ± 0.67 a |
| LPG | 591.67 ± 16.71 cd | 63.19 ± 1.96 ab | 9.61 ± 0.44 ab |
| MPG | 607.55 ± 9.85 bcd | 62.22 ± 2.25 a | 9.35 ± 0.64 ab |
| HPG | 734.47 ± 9.74 a | 64.85 ± 1.38 ab | 11.44 ± 0.35 b |
| p-value | <0.001 | 0.015 | 0.024 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Qiu, P.; Peng, Z.; Wu, J.; Bao, R.; Huang, L.; Li, X.; Shi, H.; Zhang, H.; Wang, X. Stability of Fly Maggot Peptides and Its Alleviating Effect on Lipopolysaccharide Combined with Hemocoagulase Oxidative Stress in Arbor Acres Chicks. Vet. Sci. 2024, 11, 470. https://doi.org/10.3390/vetsci11100470
Wang Q, Qiu P, Peng Z, Wu J, Bao R, Huang L, Li X, Shi H, Zhang H, Wang X. Stability of Fly Maggot Peptides and Its Alleviating Effect on Lipopolysaccharide Combined with Hemocoagulase Oxidative Stress in Arbor Acres Chicks. Veterinary Sciences. 2024; 11(10):470. https://doi.org/10.3390/vetsci11100470
Chicago/Turabian StyleWang, Qin, Pingfei Qiu, Zeru Peng, Junlong Wu, Ruiying Bao, Liangmin Huang, Xiaochun Li, Huiyu Shi, Haiwen Zhang, and Xuemei Wang. 2024. "Stability of Fly Maggot Peptides and Its Alleviating Effect on Lipopolysaccharide Combined with Hemocoagulase Oxidative Stress in Arbor Acres Chicks" Veterinary Sciences 11, no. 10: 470. https://doi.org/10.3390/vetsci11100470
APA StyleWang, Q., Qiu, P., Peng, Z., Wu, J., Bao, R., Huang, L., Li, X., Shi, H., Zhang, H., & Wang, X. (2024). Stability of Fly Maggot Peptides and Its Alleviating Effect on Lipopolysaccharide Combined with Hemocoagulase Oxidative Stress in Arbor Acres Chicks. Veterinary Sciences, 11(10), 470. https://doi.org/10.3390/vetsci11100470