

Characterization of a Very Short Meq Protein Isoform in a Marek’s Disease Virus Strain in Japan

 , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Background of the Specimen
2.2. Polymerase Chain Reaction (PCR) and DNA Sequencing
2.3. Construction of Expression Plasmids
2.4. Cell Lines and Transfection
2.5. Dual-Luciferase Reporter Assay
2.6. Statistics
3. Results
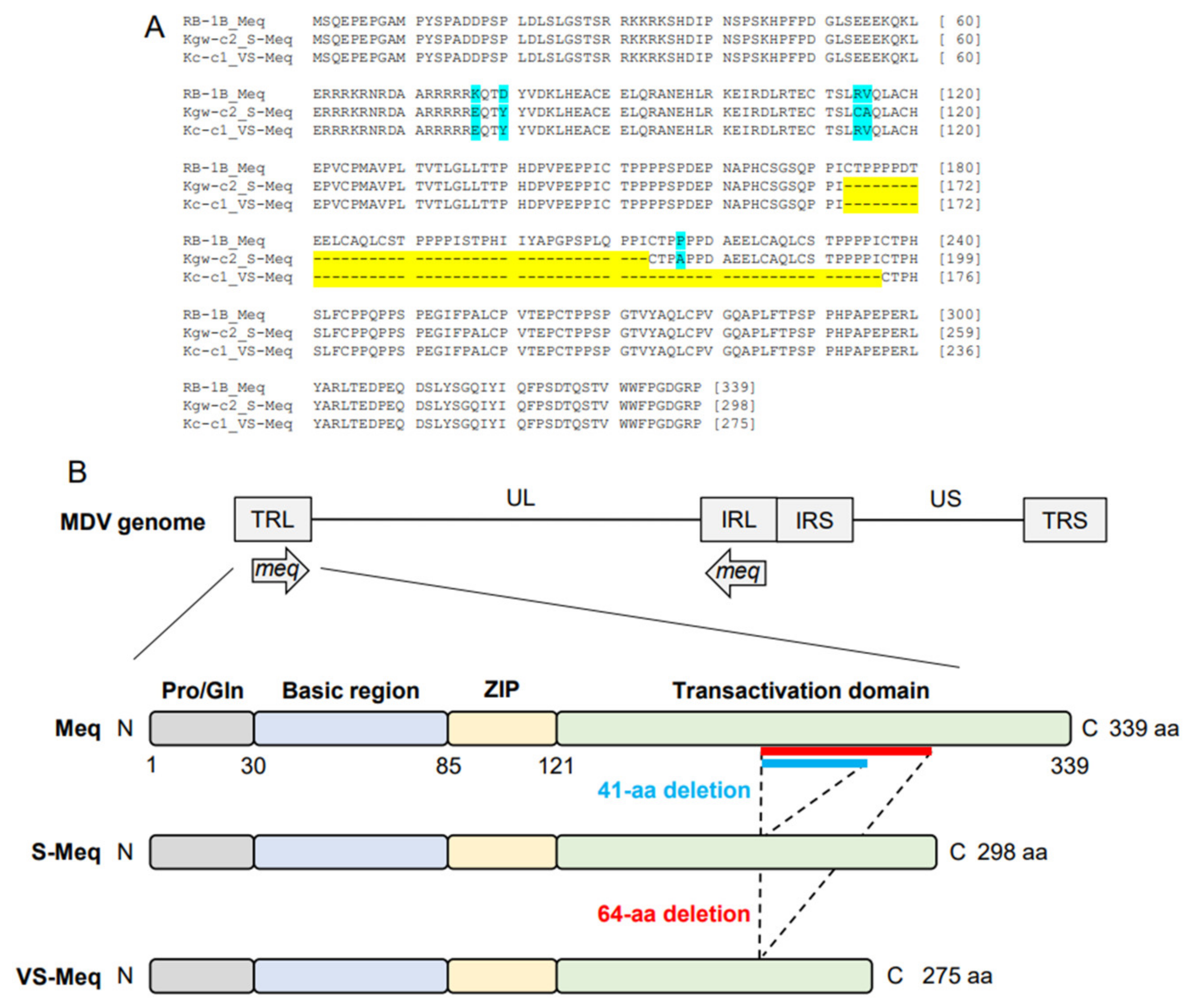
3.1. Comparison of Amino Acid Sequences of Meq
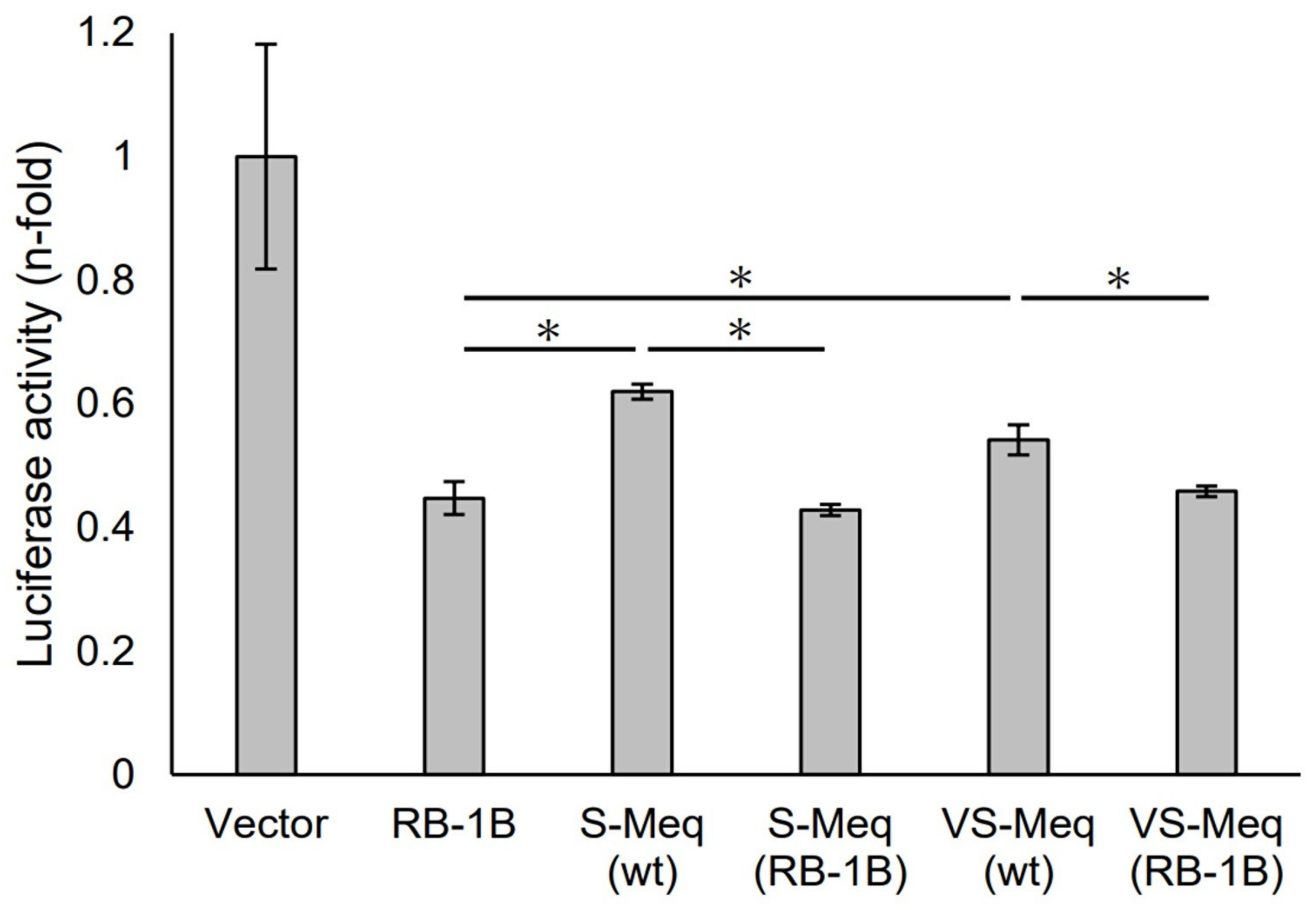
3.2. Transrepression by VS-Meq on the pp38 Promoter
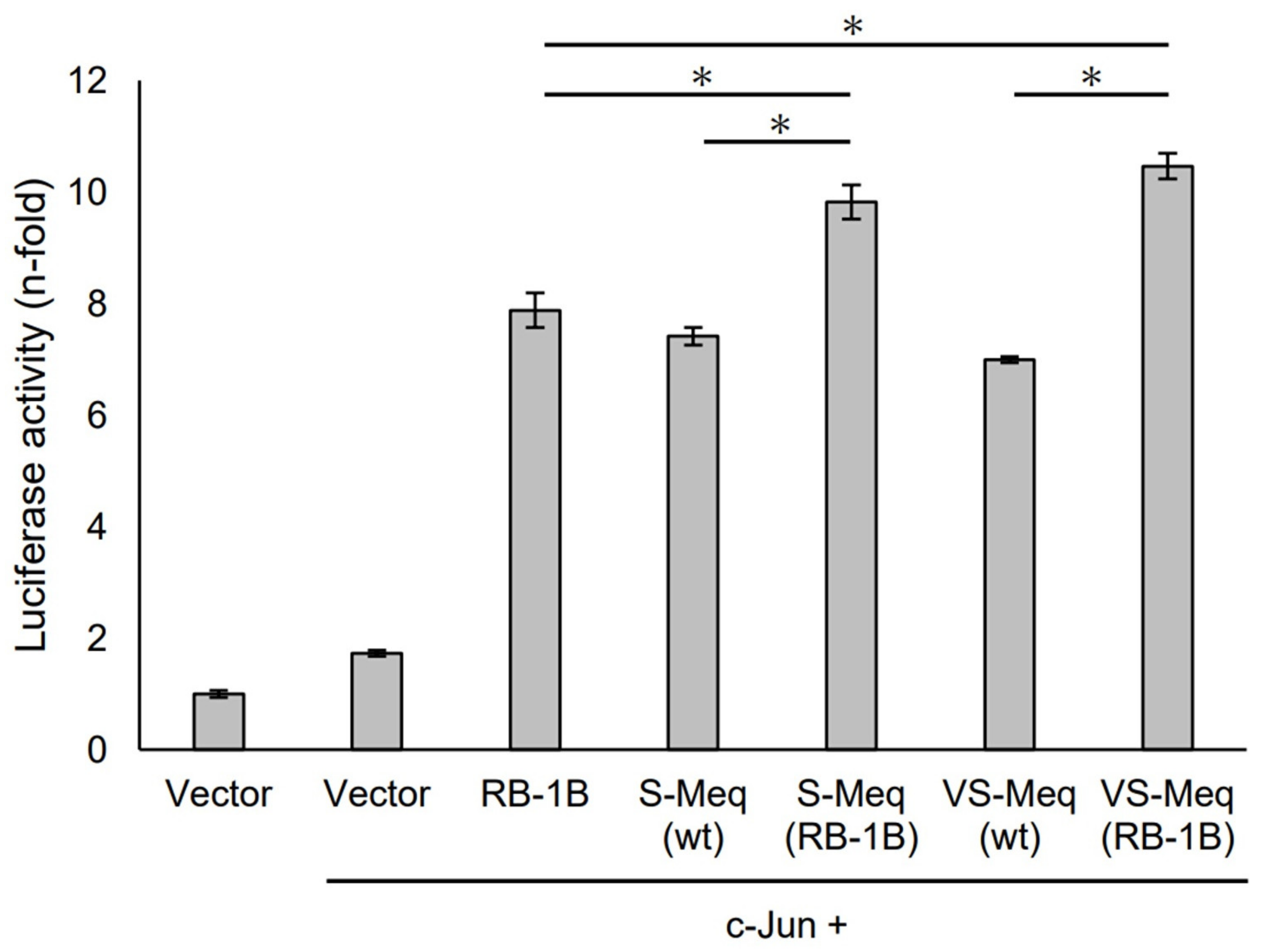
3.3. Transactivation by VS-Meq on the Meq and Bcl-2 Promoters
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bertzbach, L.D.; Conradie, A.M.; You, Y.; Kaufer, B.B. Latest Insights into Marek’s Disease Virus Pathogenesis and Tumorigenesis. Cancers 2020, 12, 647. [Google Scholar] [PubMed]
- Osterrieder, N.; Kamil, J.P.; Schumacher, D.; Tischer, B.K.; Trapp, S. Marek’s Disease Virus: From Miasma to Model. Nat. Rev. Microbiol. 2006, 4, 283–294. [Google Scholar] [CrossRef]
- Abdul-Careem, M.F.; Javaheri-Vayeghan, A.; Shanmuganathan, S.; Haghighi, H.R.; Read, L.R.; Haq, K.; Hunter, D.B.; Schat, K.A.; Heidari, M.; Sharif, S. Establishment of an Aerosol-Based Marek’s Disease Virus Infection Model. Avian Dis. 2009, 53, 387–391. [Google Scholar] [CrossRef]
- Boodhoo, N.; Gurung, A.; Sharif, S.; Behboudi, S. Marek’s Disease in Chickens: A Review with Focus on Immunology. Vet. Res. 2016, 47, 119. [Google Scholar]
- Schat, K.A. History of the First-Generation Marek’s Disease Vaccines: The Science and Little-Known Facts. Avian Dis. 2016, 60, 715–724. [Google Scholar] [CrossRef] [PubMed]
- Read, A.F.; Baigent, S.J.; Powers, C.; Kgosana, L.B.; Blackwell, L.; Smith, L.P.; Kennedy, D.A.; Walkden-Brown, S.W.; Nair, V.K. Imperfect Vaccination Can Enhance the Transmission of Highly Virulent Pathogens. PLoS Biol. 2015, 13, e1002198. [Google Scholar] [CrossRef] [PubMed]
- Padhi, A.; Parcells, M.S. Positive Selection Drives Rapid Evolution of the Meq Oncogene of Marek’s Disease Virus. PLoS ONE 2016, 11, e0162180. [Google Scholar]
- Trimpert, J.; Groenke, N.; Jenckel, M.; He, S.; Kunec, D.; Szpara, M.L.; Spatz, S.J.; Osterrieder, N.; McMahon, D.P. A Phylogenomic Analysis of Marek’s Disease Virus Reveals Independent Paths to Virulence in Eurasia and North America. Evol. Appl. 2017, 10, 1091–1101. [Google Scholar] [CrossRef]
- Lupiani, B.; Lee, L.F.; Cui, X.; Gimeno, I.; Anderson, A.; Morgan, R.W.; Silva, R.F.; Witter, R.L.; Kung, H.-J.; Reddy, S.M. Marek’s Disease Virus-Encoded Meq Gene Is Involved in Transformation of Lymphocytes but Is Dispensable for Replication. Proc. Natl. Acad. Sci. USA 2004, 101, 11815–11820. [Google Scholar] [CrossRef]
- Levy, A.M.; Izumiya, Y.; Brunovskis, P.; Xia, L.; Parcells, M.S.; Reddy, S.M.; Lee, L.; Chen, H.-W.; Kung, H.-J. Characterization of the Chromosomal Binding Sites and Dimerization Partners of the Viral Oncoprotein Meq in Marek’s Disease Virus-Transformed T Cells. J. Virol. 2003, 77, 12841–12851. [Google Scholar] [CrossRef]
- Levy, A.M.; Gilad, O.; Xia, L.; Izumiya, Y.; Choi, J.; Tsalenko, A.; Yakhini, Z.; Witter, R.; Lee, L.; Cardona, C.J.; et al. Marek’s Disease Virus Meq Transforms Chicken Cells via the v-Jun Transcriptional Cascade: A Converging Transforming Pathway for Avian Oncoviruses. Proc. Natl. Acad. Sci. USA 2005, 102, 14831–14836. [Google Scholar] [CrossRef]
- Liu, J.-L.; Kung, H.-J. Marek’s Disease Herpesvirus Transforming Protein MEQ: A c-Jun Analogue with an Alternative Life Style. Virus Genes 2000, 21, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.C.; Smith, L.P.; Kgosana, L.; Baigent, S.J.; Nair, V.; Allday, M.J. Homodimerization of the Meq Viral Oncoprotein Is Necessary for Induction of T-Cell Lymphoma by Marek’s Disease Virus. J. Virol. 2009, 83, 11142–11151. [Google Scholar] [CrossRef]
- Subramaniam, S.; Johnston, J.; Preeyanon, L.; Brown, C.T.; Kung, H.-J.; Cheng, H.H. Integrated Analyses of Genome-Wide DNA Occupancy and Expression Profiling Identify Key Genes and Pathways Involved in Cellular Transformation by a Marek’s Disease Virus Oncoprotein, Meq. J. Virol. 2013, 87, 9016–9029. [Google Scholar] [CrossRef] [PubMed]
- Conradie, A.M.; Bertzbach, L.D.; Trimpert, J.; Patria, J.N.; Murata, S.; Parcells, M.S.; Kaufer, B.B. Distinct Polymorphisms in a Single Herpesvirus Gene Are Capable of Enhancing Virulence and Mediating Vaccinal Resistance. PLoS Pathog. 2020, 16, e1009104. [Google Scholar] [CrossRef]
- Conradie, A.M.; Bertzbach, L.D.; Bhandari, N.; Parcells, M.; Kaufer, B.B. A Common Live-Attenuated Avian Herpesvirus Vaccine Expresses a Very Potent Oncogene. mSphere 2019, 4, e00658-19. [Google Scholar] [CrossRef]
- Sato, J.; Murata, S.; Yang, Z.; Kaufer, B.B.; Fujisawa, S.; Seo, H.; Maekawa, N.; Okagawa, T.; Konnai, S.; Osterrieder, N.; et al. Effect of Insertion and Deletion in the Meq Protein Encoded by Highly Oncogenic Marek’s Disease Virus on Transactivation Activity and Virulence. Viruses 2022, 14, 382. [Google Scholar]
- Lee, S.-I.; Takagi, M.; Ohashi, K.; Sugimoto, C.; Onuma, M. Difference in the Meq Gene between Oncogenic and Attenuated Strains of Marek’s Disease Virus Serotype 1. J. Vet. Med. Sci. 2000, 62, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Shamblin, C.E.; Greene, N.; Arumugaswami, V.; Dienglewicz, R.L.; Parcells, M.S. Comparative Analysis of Marek’s Disease Virus (MDV) Glycoprotein-, Lytic Antigen Pp38- and Transformation Antigen Meq-Encoding Genes: Association of Meq Mutations with MDVs of High Virulence. Vet. Microbiol. 2004, 102, 147–167. [Google Scholar] [CrossRef]
- Renz, K.G.; Cooke, J.; Clarke, N.; Cheetham, B.F.; Hussain, Z.; Fakhrul Islam, A.F.M.; Tannock, G.A.; Walkden-Brown, S.W. Pathotyping of Australian Isolates of Marek’s Disease Virus and Association of Pathogenicity with Meq Gene Polymorphism. Avian Pathol. 2012, 41, 161–176. [Google Scholar] [CrossRef]
- Ajithdoss, D.K.; Reddy, S.M.; Suchodolski, P.F.; Lee, L.F.; Kung, H.J.; Lupiani, B. In Vitro Characterization of the Meq Proteins of Marek’s Disease Virus Vaccine Strain CVI988. Virus Res. 2009, 142, 57–67. [Google Scholar] [CrossRef]
- Murata, S.; Yamamoto, E.; Sakashita, N.; Maekawa, N.; Okagawa, T.; Konnai, S.; Ohashi, K. Research Note: Characterization of S-Meq Containing the Deletion in Meq Protein’s Transactivation Domain in a Marek’s Disease Virus Strain in Japan. Poult. Sci. 2021, 100, 101461. [Google Scholar] [CrossRef]
- Wajid, S.J.; Katz, M.E.; Renz, K.G.; Walkden-Brown, S.W. Prevalence of Marek’s Disease Virus in Different Chicken Populations in Iraq and Indicative Virulence Based on Sequence Variation in the EcoRI-Q (Meq) Gene. Avian Dis. 2013, 57, 562–568. [Google Scholar] [CrossRef] [PubMed]
- Ghalyanchilangeroudi, A.; Hosseini, H.; Nazarpak, H.H.; Molouki, A.; Dezfoulian, O.; Morshed, R. Molecular Characterization and Phylogenetic Analysis of Marek’s Disease Virus in Iran. Avian Dis. 2022, 66, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Mescolini, G.; Lupini, C.; Felice, V.; Guerrini, A.; Silveira, F.; Cecchinato, M.; Catelli, E. Molecular Characterization of the Meq Gene of Marek’s Disease Viruses Detected in Unvaccinated Backyard Chickens Reveals the Circulation of Low- and High-Virulence Strains. Poult. Sci. 2019, 98, 3130–3137. [Google Scholar]
- Molouki, A.; Ghalyanchilangeroudi, A.; Abdoshah, M.; Shoushtari, A.; Abtin, A.; Eshtartabadi, F.; Mahmoudzadeh Akhijahani, M.; Ziafatikafi, Z.; Babaeimarzango, S.S.; Allahyari, E.; et al. Report of a New Meq Gene Size: The First Study on Genetic Characterisation of Marek’s Disease Viruses Circulating in Iranian Commercial Layer and Backyard Chicken. Br. Poult. Sci. 2022, 63, 142–149. [Google Scholar] [CrossRef]
- Liu, H.; Naismith, J.H. An Efficient One-Step Site-Directed Deletion, Insertion, Single and Multiple-Site Plasmid Mutagenesis Protocol. BMC Biotechnol. 2008, 8, 91. [Google Scholar] [CrossRef] [PubMed]
- Murata, S.; Hashiguchi, T.; Hayashi, Y.; Yamamoto, Y.; Matsuyama-Kato, A.; Takasaki, S.; Isezaki, M.; Onuma, M.; Konnai, S.; Ohashi, K. Characterization of Meq Proteins from Field Isolates of Marek’s Disease Virus in Japan. Infect. Genet. Evol. 2013, 16, 137–143. [Google Scholar]
- Chang, K.S.; Ohashi, K.; Onuma, M. Diversity (Polymorphism) of the Meq Gene in the Attenuated Marek’s Disease Virus (MDV) Serotype 1 and MDV-Transformed Cell Lines. J. Vet. Med. Sci. 2002, 64, 1097–1101. [Google Scholar] [CrossRef]
- Murata, S.; Machida, Y.; Isezaki, M.; Maekawa, N.; Okagawa, T.; Konnai, S.; Ohashi, K. Genetic Characterization of a Marek’s Disease Virus Strain Isolated in Japan. Virol. J. 2020, 17, 186. [Google Scholar]
- Tian, M.; Zhao, Y.; Lin, Y.; Zou, N.; Liu, C.; Liu, P.; Cao, S.; Wen, X.; Huang, Y. Comparative Analysis of Oncogenic Genes Revealed Unique Evolutionary Features of Field Marek’s Disease Virus Prevalent in Recent Years in China. Virol. J. 2011, 8, 121. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Zeb, J.; Hussain, S.; Aziz, M.U.; Circella, E.; Casalino, G.; Camarda, A.; Yang, G.; Buchon, N.; Sparagano, O. A Review on the Marek’s Disease Outbreak and Its Virulence-Related Meq Genovariation in Asia between 2011 and 2021. Animals 2022, 12, 540. [Google Scholar] [CrossRef]
- Cory, S. Regulation of Lymphocyte Survival by the bcl-2 Gene Family. Annu. Rev. Immunol. 1995, 13, 513–543. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Li, X.; Shen, Y.; Qiu, Y.; Shi, Z.; Shao, D.; Jin, Y.; Chen, H.; Ding, C.; Li, L.; et al. The Meq Oncoprotein of Marek’s Disease Virus Interacts with p53 and Inhibits Its Transcriptional and Apoptotic Activities. Virol. J. 2010, 7, 348. [Google Scholar] [PubMed]
- Zhao, Y.; Kurian, D.; Xu, H.; Petherbridge, L.; Smith, L.P.; Hunt, L.; Nair, V. Interaction of Marek’s Disease Virus Oncoprotein Meq with Heat-Shock Protein 70 in Lymphoid Tumour Cells. J. Gen. Virol. 2009, 90, 2201–2208. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-L.; Ye, Y.; Qian, Z.; Qian, Y.; Templeton, D.J.; Lee, L.F.; Kung, H.-J. Functional Interactions between Herpesvirus Oncoprotein MEQ and Cell Cycle Regulator CDK2. J. Virol. 1999, 73, 4208–4219. [Google Scholar] [CrossRef]
- Brown, A.C.; Baigent, S.J.; Smith, L.P.; Chattoo, J.P.; Petherbridge, L.J.; Hawes, P.; Allday, M.J.; Nair, V. Interaction of MEQ Protein and C-Terminal-Binding Protein is Critical for Induction of Lymphomas by Marek’s Disease Virus. Proc. Natl. Acad. Sci. USA 2006, 103, 1687–1692. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Lupiani, B.; Izumiya, Y.; Reddy, S.M. Marek’s Disease Virus Meq Oncoprotein Interacts with Chicken HDAC 1 and 2 and Mediates Their Degradation via Proteasome Dependent Pathway. Sci. Rep. 2021, 11, 637. [Google Scholar] [CrossRef]
- Li, K.; Liu, Y.; Xu, Z.; Zhang, Y.; Luo, D.; Gao, Y.; Qian, Y.; Bao, C.; Liu, C.; Zhang, Y.; et al. Avian Oncogenic Herpesvirus Antagonizes the cGAS-STING DNA-Sensing Pathway to Mediate Immune Evasion. PLoS Pathog. 2019, 15, e1007999. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Purpose | Sequence (5′-3′) |
|---|---|---|
| MS-10 | Sequencing | ATGTCTCAGGAGCCAGAGCCGGGCGCT |
| M-AS | Sequencing | GGGGCATAGACGATGTGCTGCTGAG |
| EcoRI Meq | Cloning into pCI-neo vector | CCCGAATTCATGTCTCAGGAGCCAGAGCCGGGCGCT |
| Not I Meq | Cloning into pCI-neo vector | ATAAGAAATGGGGCCGCGGCGGGGCATAGACGATGTGCTGCTGAG |
| E77K Fw | Mutagenesis | AGGAAGCAGACGTACTATGTAGACA |
| E77K Rv | Mutagenesis | TGTCTACATAGTACGTCTGCTTCCTT |
| Y80D Fw | Mutagenesis | AGAAGACGCAGGGAGCAGACGGACT |
| Y80D Rv | Mutagenesis | AGTCCGTCTGCTCCCTGCGTCTTC |
| E77K Y80D Fw | Mutagenesis | AGAAGACGCAGGAAGCAGACGGACT |
| E77K Y80D Rv | Mutagenesis | AGTCCGTCTGCTTCCTGCGTCTTCT |
| C114R A115V Fw | Mutagenesis | GAGTGCACGTCCCTGCGTGTACAGTTGGCTTGT |
| C114R A115V Rv | Mutagenesis | TGACAAGCCAACTGTACACGCAGGGACGTGCACTC |
| A176P Fw | Mutagenesis | ATCTGTACCCCCGCTCCTCCCGATA |
| A176P Rv | Mutagenesis | TATCGGGAGGAGCGGGGGTACAGAT |
| Strain | Isoform | Country | Accession No. | Length | Number of PPPP | ||
|---|---|---|---|---|---|---|---|
| ORF (bp) | Amino Acid (aa) | Deletion (aa) d | |||||
| RB-1B | Meq | USA | EF523390 | 1020 | 339 | 0 | 5 |
| CVI988 | S-Meq | Netherlands | AY243338 | 897 | 298 | 41 | 3 |
| 855/17 | S-Meq | Italy | MK139678 | 897 | 298 | 41 | 2 |
| MDV/2/SA a | S-Meq | Saudi Arabia | LC385871 | 897 | 298 | 41 | 3 |
| Iraq3A | S-Meq | Iraq | KC243262 | 897 | 298 | 41 | 2 |
| Iraq10A | S-Meq | Iraq | KC243264 | 897 | 298 | 41 | 2 |
| Iraq6F | S-Meq | Iraq | KC243263 | 897 | 298 | 41 | 2 |
| 99-26 | S-Meq | Iran | MZ962187 | 897 | 298 | 41 | 2 |
| 99-35 | VS-Meq | Iran | MW990216 | 798 | 265 | 74 | 1 |
| VS-Meq b | VS-Meq | Japan | AB087744 | 744 | 247 | 92 | 1 |
| Nr-c1 c | Meq | Japan | LC385871 | 1020 | 339 | 0 | 3 |
| Kgw-c2 | S-Meq | Japan | CL385874 | 897 | 298 | 41 | 2 |
| Sit-c1 | VS-Meq | Japan | BBE28998 | 828 | 275 | 64 | 1 |
| Kc-c1 | VS-Meq | Japan | LC790435 | 828 | 275 | 64 | 1 |
| Basic Region | Leucine Zipper | Transactivation Domain | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Strain | 36 | 70 | 71 | 77 | 78 | 80 | 88 | 100 | 110 | 114 | 115 | 119 | 141 | 168 | 176 * | 217 * | 218 * | 221 * | 281 * | 326 * |
| RB-1B | S | A | A | K | Q | D | A | R | C | R | V | C | H | S | P | P | P | A | G | T |
| CVI988 | S | A | A | E | Q | D | A | R | C | R | V | C | H | S | - | P | P | A | G | I |
| 855/17 | S | A | A | E | Q | Y | A | R | S | R | V | C | H | S | - | P | S | T | G | T |
| MDV/2/SA | S | A | A | E | Q | Y | A | R | S | R | V | C | P | S | - | P | P | A | G | T |
| Iraq3A | S | A | A | E | Q | Y | A | R | C | R | V | R | H | P | - | A | P | A | G | T |
| Iraq10A | S | A | S | E | Q | D | A | R | C | R | V | C | H | P | - | A | P | A | G | T |
| Iraq6F | S | A | S | E | Q | D | A | R | C | R | V | C | H | S | - | A | P | A | G | T |
| 99-26 | T | A | A | K | H | D | A | R | C | R | V | R | H | S | - | A | P | A | D | T |
| 99-35 | S | A | A | E | Q | Y | T | R | C | R | A | C | H | S | - | - | - | - | G | T |
| VS-Meq | S | S | A | K | Q | D | A | H | C | R | V | C | H | S | - | - | - | - | G | T |
| Nr-c1 | S | A | A | E | Q | Y | A | R | C | R | V | C | H | S | S | A | P | A | G | T |
| Kgw-c2 | S | A | A | E | Q | Y | A | R | C | C | A | C | H | S | - | A | P | A | G | T |
| Sit-c1 | S | A | A | E | Q | Y | A | R | C | R | V | C | H | S | - | - | - | - | G | T |
| Kc-c1 | S | A | A | E | Q | Y | A | R | C | R | V | C | H | S | - | - | - | - | G | T |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Motai, Y.; Murata, S.; Sato, J.; Nishi, A.; Maekawa, N.; Okagawa, T.; Konnai, S.; Ohashi, K. Characterization of a Very Short Meq Protein Isoform in a Marek’s Disease Virus Strain in Japan. Vet. Sci. 2024, 11, 43. https://doi.org/10.3390/vetsci11010043
Motai Y, Murata S, Sato J, Nishi A, Maekawa N, Okagawa T, Konnai S, Ohashi K. Characterization of a Very Short Meq Protein Isoform in a Marek’s Disease Virus Strain in Japan. Veterinary Sciences. 2024; 11(1):43. https://doi.org/10.3390/vetsci11010043
Chicago/Turabian StyleMotai, Yoshinosuke, Shiro Murata, Jumpei Sato, Akihito Nishi, Naoya Maekawa, Tomohiro Okagawa, Satoru Konnai, and Kazuhiko Ohashi. 2024. "Characterization of a Very Short Meq Protein Isoform in a Marek’s Disease Virus Strain in Japan" Veterinary Sciences 11, no. 1: 43. https://doi.org/10.3390/vetsci11010043
APA StyleMotai, Y., Murata, S., Sato, J., Nishi, A., Maekawa, N., Okagawa, T., Konnai, S., & Ohashi, K. (2024). Characterization of a Very Short Meq Protein Isoform in a Marek’s Disease Virus Strain in Japan. Veterinary Sciences, 11(1), 43. https://doi.org/10.3390/vetsci11010043